河北壩上樟子松人工林徑向生長及其對氣候因素的響應
2022-03-30 08:58:04劉亞玲信忠保李宗善買爾當克依木閆騰飛
生態學報 2022年5期
關鍵詞:生長
劉亞玲,信忠保,*, 李宗善, 買爾當·克依木, 閆騰飛
1 北京林業大學水土保持學院, 北京 100083 2 中國科學院生態環境研究中心城市與區域生態國家重點實驗室, 北京 100085 3 張北縣林業局, 張北 076450
隨著人類文明的進步和科技的發展,人為溫室氣體排放量不斷增加,導致全球溫度快速上升,氣候變化成為世界上最關注的科學熱點之一[1]。森林作為地球上最大的陸地生態系統,是地球系統物質循環和能量流動中極為重要的一環[2—3],在維持全球的生態平衡中扮演著不可或缺的角色。全球氣候變暖導致干旱的發生強度和發生頻率在逐年增加,導致部分森林樹種出現衰退的現象,生態系統的結構和功能發生了巨大變化[4—5]。生態彈性是評估樹木遭受干擾后的抵抗能力和恢復到初始狀態的能力[6]。生態彈性可以準確評估森林對當前和過去氣候變化的反應,有助于預測未來森林對氣候變化的適應能力[7]。樹木年輪以其定年準確,分辨率高等優點已廣泛應用于樹木對干擾和環境脅迫的響應研究[8]。通過研究樟子松的生態彈性,可以預測樟子松人工林對氣候變化的適應性,為樟子松的經營管理提供理論依據。
河北壩上地區干旱少雨,植被稀少,土壤沙化嚴重,生態環境脆弱。為改善當地生態環境,20世紀70年代末營造了以小葉楊為主的人工防護林,20世紀90年代初補植了以樟子松為主的人工防護林。該區防護林是三北防護林工程的重要組成部分,同時是京津風沙源的重要生態屏障,經過40 a的建設其生態效益和經濟效益顯著[9]。但自2002年開始,該地區小葉楊防護林逐步出現退化、枯死現象,危及當地生態平衡,樟子松防護林的優勢逐漸凸顯[10],在河北壩上地區衰退楊樹林的更新改造中,不再使用小葉楊作為主要造林樹種,而普遍選用耐寒耐旱的樟子松。目前我國樟子松的樹木年輪學研究主要集中在內蒙古呼倫貝爾沙地和毛烏素沙地、黑龍江大興安嶺和帽兒山、遼寧章古臺以及河北塞罕壩地區。大興安嶺和帽兒山等地區樟子松生長主要受溫度的影響[11—13];呼倫貝爾沙地、毛烏素沙地、章古臺及塞罕壩等地區樟子松徑向生長主要受降水的限制,并且章古臺地區樟子松出現了生長衰退的現象[14—16]。河北壩上地區樟子松徑向生長對氣候響應的研究未見報道,因此我們研究了河北壩上地區樹齡最大的樟子松林,研究其徑向生長過程、限制其生長的主要氣候因素以及遭遇干旱后樟子松的動態變化,對評估樟子松在河北壩上地區的生長適宜性具有重要代表性和前瞻性。
樟子松(Pinussylvestrisvar.mongolica)原產于中國大興安嶺、紅花爾基和呼倫貝爾沙地,為淺根性樹種,其耐寒耐旱性強,可在-40—-50 ℃的低溫中生長,對水分要求不嚴格,并且因其抗逆性強,壽命長,有很好的生態和經濟價值而被廣泛應用于三北防護林的造林工程中。河北壩上地區在20世紀90年代初種植的樟子松人工林目前沒有出現衰退林相,本研究以河北壩上地區樟子松防護林為研究對象,運用樹木年輪氣候學原理及方法,建立樟子松差值年表、BAI年表。旨在揭示樟子松徑向生長特征,明確限制樟子松在該地區生長的主要氣候因素,以及樟子松在河北壩上地區的適宜性。研究結果對于提高森林生態防護作用非常重要,為未來衰退楊樹林的更新改造和樟子松人工林的栽植和經營管理提供科學依據。
1 研究區與研究方法
1.1 研究區概況
采樣點位于河北省張家口市張北縣波羅素林場內(41.336°N,114.879°E),是河北壩上樟子松人工林的主要分布區(圖1)。張北縣地處內蒙古東南緣,地勢東南高西北低,東南部為壩上高原區,西北部為丘陵區,中部屬平原區,丘陵洼地交錯分布。研究區屬該縣中部平原地區,海拔1370—1390 m,相對高差20 m。氣候為中溫帶大陸性季風氣候,四季分明,夏季炎熱多雨,冬季寒冷干燥。年平均氣溫為4.0 ℃(1992—2019年),最冷月(1月)的均溫為-14.4 ℃,最熱月(7月)的均溫為19.7 ℃,極端高溫為25.3 ℃,極端低溫為-19.7 ℃;年均降水量為390.1 mm,6—9月降水量占全年降水量的75%,年蒸發量1700—1800 mm,為降水量的4—5倍(圖2)。1992年以來平均氣溫(0.34 ℃/10a)、最高氣溫(0.35 ℃/10a)和最低氣溫(0.21 ℃/10a)均上升迅速;降水量呈下降趨勢,最大降水量(1995年)533.9 mm,最小降水量(1997年)245.2 mm;帕默爾干旱指數下降明顯(-0.30/10a);多年平均相對濕度為56.9%(圖3)。年均日照時數2897.8 h,年均7級以上大風日數30 d左右。主要土壤類型為栗鈣土類。喬木主要包括樟子松(Pinussylvestrisvar.mongolica)、小葉楊(Populussimonii)、山楊(Populusdavidiana)、云杉(Piceaasperata)等;灌木主要包括檸條(Caraganakorshinskii)、沙棘(Hippophaerhamnoides)、小紅柳(Salixmicrostachya)等;草本植物豐富,主要分為鹽漬化灘地蒿類、低洼地草甸類以及坡梁地禾本類[17]。該地區樟子松為20世紀90年代種植的人工純林,造林時使用的是2年生裸苗,初植密度為1300株/hm2,后期撫育管理措施缺乏,現存密度為950株/hm2,行距為4 m,株距為3 m,研究區內樟子松樹齡基本一致。

圖1 河北壩上地區樹木年輪采樣點及氣象站分布圖

圖2 1992—2019年河北壩上地區溫度降水季節分布

圖3 河北壩上地區氣候因素年際變化趨勢(1992—2019年)
1.2 樣品采集及年表建立
在河北壩上地區波羅素林場內選取40株長勢良好、樹齡較大、受病蟲干擾小的樟子松,在胸高處(1.3 m)用生長錐沿東西方向鉆取樹芯,每棵樹鉆取2—3個樹芯,共取樣80個。采集的樹芯根據國際上樹輪樣品標準處理過程進行預處理。在實驗室內將采集的樹芯固定風干[18],用不同粗細的干砂紙(400目、800目和1200目)依次打磨到樣芯光滑且亮,并且在電子顯微鏡下能看清樟子松的年輪。利用骨架圖法交叉定年后,用年輪分析儀Win DENDROTM(Regent instrument inc.Canada)測量樟子松樹輪的原始寬度,其精度為0.001 mm。用COFECHA程序對交叉定年和測量結果進行檢驗,剔除相關性差的樣芯,最后余下57根樣芯進行年表建立[19]。
由于樣本樹齡較短,研究主要集中在樹木生長的高頻變化上,故應去除樹木隨年齡增長的生長趨勢和其他非一致性擾動導致的非氣候信號,而突出其中的氣候信號[20],利用ARSTAN程序建立年表,去趨勢方法為負指數函數,該方法可減少樹間競爭、地形、地貌等因素對年表指數擬合的影響,最終得到樟子松標準年表(Standard chronology,STD)、自回歸年表(Arstan chronology,ARS)和差值年表(Residual chronology,RES)。本研究選用RES進行樹木生長與氣候間的相關分析,RES是在STD的基礎上,考慮到森林內樹與樹的競爭以及可能存在的人類活動導致的樹輪寬度序列的低頻變化,以及時間序列的自回歸模式對STD進行擬合并再次標準化,去掉樹木個體特有的前期生理條件對后期生長造成的連續性影響而建立的一種年表,它只含有群體共有的高頻變化。
1.3 胸高斷面積增量
當樹輪寬度相對穩定或變窄時,樹木生長量可能繼續增大[21],為了保留氣候條件對樹木生長影響的信息,本文使用BAI來評估研究區內樟子松人工林的生長速率和生長趨勢。樹木胸高斷面積增量:
式中,Rn是n年的樹盤半徑;Rn-1是n-1年的樹輪半徑。
1.4 氣象數據獲取
本研究采用的氣候要素數據來源于張北氣象站(114.42°E,41.09°N,海拔為1393.3 m),時間跨度為1992—2019年。氣候要素為月均溫度、月均最低溫度、月均最高溫度、月均相對濕度和降水量,以上數據從中國氣象數據網(http://data.cma.cn/)獲取。由于溫度和降水等綜合作用對土壤有效含水量的影響明顯,本研究選用帕默爾干旱指數(PDSI)反映樟子松生長與土壤水分的關系,其數據來源于荷蘭皇家氣象局服務網(http://climexp.knmi.nl/start.cgi)中的PDSI網格點數據集[22](114.50°—115.00°E,41.00°—41.50°N),時間跨度為1992—2017年。考慮到樹木生長的連貫性,當年的生長必定受到前一年氣候的影響,在進行氣候響應分析時選用上一年6月到當年10月共17個月的氣象數據。
1.5 數據處理
該地區位于半干旱地區,干旱缺水是其主要氣候特征,在干旱年份樟子松生長受到嚴重影響,樟子松徑向生長顯著下降,結合樟子松差值年表和BAI年表與PDSI年際波動數據,研究樟子松生長遭遇干旱時其生態彈性。選定樟子松3個生長低點年份2000年(PDSI=-1.92)、2007年(PDSI=-3.34)和2014年(PDSI=-1.54)來研究樟子松的抵抗力、恢復力和彈性力。
利用Lloret等[23]的公式來進行樟子松抵抗力、恢復力和彈性力的計算:
Rt=Gr/Ga;Rc=Gb/Gr;Rs=Gb/Ga
式中,Rt、Rc和Rs分別表示樹木的抵抗力、恢復力和彈性力;Gr表示生長顯著下降時數值;Ga和Gb分別表示生長顯著下降前3年和后3年的平均值。Rt>0.75說明樹木對干旱具有較高的抵抗力;Rc>1.25說明發生干旱事件后樹木仍有較好的恢復力。
本文利用Excel 2013軟件分析河北壩上地區樟子松的徑向生長過程,揭示該地區樟子松的生長狀況。使用Dendroclim 2002國際樹木年輪程序[24]分析年表與各氣候數據間的相關性,并用Origin 2018繪制樟子松差值年表、BAI年表以及相關圖表的制作。
2 研究結果
2.1 樟子松年表統計特征
本研究建立了樟子松標準年表和差值年表,年表質量的高低決定著研究結果是否準確,利用標準差、平均敏感度、序列間相關系數、信噪比、樣本總體代表性等來評價年表質量(表1)。標準差反映年表的上下波動,標準差越大表明年表所包含的氣候信息越多,對氣候越敏感[25],樟子松標準年表和差值年表的標準差分別為0.358和0.308。一介自相關系數反映上一年氣候狀況對當年年輪寬度生長的影響強弱,其值分別為0.295和0.063,表明樟子松徑向生長受上年氣候條件的影響[26]。樟子松序列間相關系數分別為0.579和0.607,表示不同樹芯間的年輪寬度變化具有較好的一致性,數據質量較好[20]。平均敏感度分別為0.304和0.324,信噪比分別為12.364和13.906,主要代表氣候的短期變化或高頻變化,平均敏感度大于0.15以及信噪比大于4代表年表質量較高[27—28]。樣本總體代表性分別為0.925和0.933,是指所采集樣芯對整個區域的代表程度,通常樣本總體代表性大于0.85表明年表質量較高[29]。綜上所述,樟子松差值年表質量較好,以下運用差值年表來進行氣候響應分析。樟子松差值年表的起始年份樣本量大于5根,可以較好地反映徑向生長年際波動(圖4)。

圖4 河北壩上地區樟子松差值年表

表1 樟子松差值年表統計特征
2.2 樟子松樹輪寬度和BAI變化趨勢
河北壩上地區樟子松生長隨樹齡增加呈現在波動中下降趨勢,在各個生長階段的徑向生長速率差距顯著(圖5),在1—10 a生長速率較快為(4.81±0.63)mm/a;11—20 a樟子松生長速率逐漸下降,平均生長速率為(2.95±0.87)mm/a,相對1—10 a生長速率下降了38.7%;21—28 a樟子松徑向生長處于平穩狀態,其平均生長速率為(2.20±0.43)mm/a,相對1—10 a生長速率下降54.3%。
河北壩上地區樟子松樹輪寬度年際變化顯示(圖5),樟子松樹輪寬度在1992—2000年期間為生長平穩期,平均生長速率為(4.49±1.15)mm/a;在2001—2010年期間呈現下降趨勢,平均生長速率為(3.62±1.39)mm/a,較前期生長速率下降了19.3%;2011—2019年期間樟子松生長呈平穩特征,平均生長速率為(2.21±0.68)mm/a,較前期生長速率下降了50.8%。胸高斷面積生長量(BAI)是每年樹輪增加的面積,相比與年輪寬度,它能夠更好地反映樹木的生長速率和生長趨勢[30]。樟子松生長期間BAI年際波動較大(圖5),1992—2000年樟子松BAI一直呈上升趨勢,平均生長速率為(5.09±2.26)cm2/a;2001—2019年期間呈平穩特征,平均生長率為(10.46±0.67)cm2/a,較前期生長速率增加了105.5%,表明樟子松表現出穩定、旺盛的生產力狀態,尚未出現生產力衰退現象。

圖5 河北壩上地區樟子松樹輪寬度徑向生長特征、樹輪寬度和胸高斷面積增量(BAI)年際變化
2.3 樟子松RES與氣候因素的關系
河北壩上地區樟子松徑向生長受溫度和降水兩者共同的影響。樟子松RES與溫度因素多呈負相關,樟子松徑向生長與上一年9、10、12月及當年7月份的平均溫度和最高溫度呈顯著負相關,其中最高溫度對樟子松生長影響更為顯著(P<0.05,圖6)。樟子松徑向生長與降水和相對濕度多呈正相關,樟子松樹輪寬度與上一年9月及當年5、7月份的降水和相對濕度呈顯著正相關(P<0.05,圖7)。帕默爾干旱指數在干旱區的研究中廣泛應用,它是根據降水、溫度以及土壤含水量數據進行計算,代表土壤水分的有效狀況,其值越小表明有效土壤水分越低。樟子松徑向生長與帕默爾干旱指數多呈正相關,其中與上一年9月—當年10月份的帕默爾干旱指數呈顯著正相關,該地區樟子松遭受干旱脅迫(P<0.05,圖7)。

圖6 河北壩上樟子松差值年表與溫度因素的相關關系

圖7 樟子松差值年表與降水、相對濕度和PDSI的相關關系
2.4 樟子松的恢復力、抵抗力與彈性力
該地區氣候暖干化趨勢明顯,樟子松生長極易遭受嚴重干旱脅迫,在干旱缺水年份樟子松生長受到嚴重影響,結合樟子松差值年表和BAI年表與PDSI年際波動數據,研究樟子松生長顯著下降時其生態彈性。選出2000年、2007年和2014年來分析樟子松生長顯著下降時的抵抗力、恢復力和彈性力(圖8)。通過樟子松差值年表研究樟子松的生態彈性,發現樟子松隨樹齡增加,抵抗力由0.82下降到0.55,在2000年樟子松具有較高的抵抗力;樟子松的恢復力呈上升趨勢,2014年達到了1.91,恢復力較強;樟子松的彈性力呈現穩定特征。通過樟子松BAI年表研究樟子松的生態彈性,發現樟子松隨樹齡增加,抵抗力同樣在下降,由0.93下降到0.52,僅在2000年具有較高的抵抗力;樟子松的恢復力呈上升趨勢,2014年達到了1.72;樟子松的彈性力較為穩定(圖9)。

圖8 樟子松差值年表和BAI年表與PDSI的關系
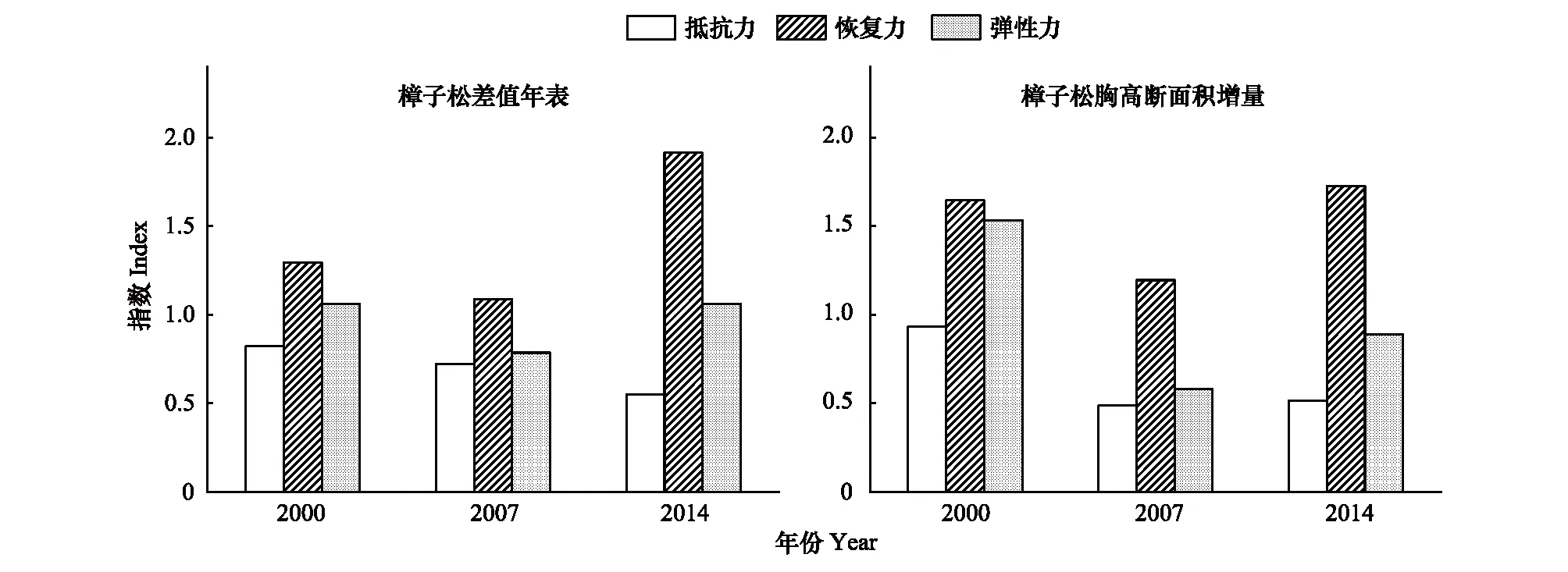
圖9 樟子松的抵抗力、恢復力和彈性力
3 討論
3.1 樟子松樹輪寬度及BAI變化特征
樟子松在2000、2007和2014年遭遇干旱時生長下降嚴重。樟子松樹輪寬度在2000年時達到生長低點,2000年當地干旱指數達到-1.92,處于輕微干旱等級,干旱脅迫導致樟子松徑向生長下降。干旱脅迫對樹木年輪寬度影響顯著,樹木在面對嚴重的干旱時會表現出應激癥狀,如葉片過早的發黃、脫落,樹木處于較脆弱的狀態,碳水化合物儲量變少,樹木的高度、年輪寬度等因此受到限制[31]。2007年和2014年均為干旱年,尤其2007年達到嚴重干旱程度,樟子松樹輪寬度在這兩個時期達到較窄水平。2000年樹輪寬度有所上升,在第二年或后幾年出現顯著的下降,表明樟子松對氣候的響應具有明顯的“滯后性”。樹木在受到極端干旱脅迫時,當年的響應可能不會十分明顯,在極端干旱的次年或之后幾年形成窄輪[31]。《中華人民共和國林業行業標準主要樹種齡級與齡組劃分》(LY/T2908—2017)中將樟子松的生長劃分為五個階段:1—30 a為幼齡期,31—50 a為中齡期,51—60 a為近熟期,61—80 a為成熟期,80 a之后為過熟期。河北壩上地區樟子松樹輪寬度隨樹齡的增大,徑向生長速率呈下降趨勢,在21—28 a時處于平穩期,樟子松的生長階段較國家行標的生長階段劃分提前,當地樟子松可能出現衰退的現象。
干旱脅迫影響樟子松BAI增長速率,樟子松BAI在1992—2000年期間增長迅速,在2007年遭遇嚴重干旱事件后其增長速率下降至近幾年最低值,其后樟子松BAI增長逐漸恢復,說明樟子松遭遇干旱后有一定的恢復機能,可以較好的適應河北壩上地區干旱的氣候條件。當地樟子松的樹齡較小,僅有20—30 a,理論上樟子松的最大樹齡可達350年左右[32],當地樟子松BAI年際增長量變化趨勢不能完全反映當地樟子松是否出現衰退現象,還需要對當地樟子松進行連續觀測。
3.2 樟子松差值年表與氣候因子關系特征
河北壩上地區樟子松年內樹干徑向生長呈現季節性規律,主要劃分為4個階段:4月初到5月中旬的春季萌芽期、5月中旬到7月中旬的夏季生長期、7月中旬到10月中旬的秋冬交替期以及10月中旬到下年4月初的冬季休眠期[33]。河北壩上地區樟子松徑向生長主要與上一年生長季末期(9、10月)、休眠期(12月)和當年生長季(7月)的月平均最高溫度呈顯著負相關,高溫顯著抑制樟子松的徑向生長。上年生長季末期和休眠期溫度越高,導致蒸發作用增強,水分減少,在第二年進入生長季后沒有充足的水分,抑制樟子松的光合速率,樟子松徑向生長受到限制[34]。生長季內溫度過高,樹木蒸騰作用和呼吸作用均加強,光合作用減弱,消耗掉過多的養分,樟子松徑向生長速率減緩,易形成窄輪[35—36]。
樟子松差值年表與上年生長季末(9月)及當年生長季(5、7月)降水呈顯著正相關,其中受上年降水的影響更為顯著,說明樟子松徑向生長與降水存在“滯后效應”,上年生長季后期降水可以為根系層補充充足的水分,為促進當年樹木的生長起著重要作用[37]。5、7月樟子松進入生長期,氣溫逐漸增加,蒸騰作用加強,導致土壤水分不足,樹木生長受到限制,有效的降水可以增加土壤水分,為樟子松的正常生長提供必要的水分條件。相對濕度是受溫度和降水共同調控的氣候因子[38],影響樟子松的生長,樟子松生長主要受當年生長季(5、7、8月)的影響。植物生長空氣濕度在50%—80%為宜,而河北壩上地區相對濕度在56%左右,屬于半干旱氣候,當樹木周圍環境中的相對濕度增高時,會促進植物氣孔張開,CO2進入葉肉細胞,促進植物的光合作用,有助于樟子松的徑向生長。
PDSI反映干旱嚴重程度,具有較好時空可比較性,不但綜合了水分虧缺量和持續時間因子對干旱程度的影響,并充分考慮了干旱持續時間,是評估干旱程度的較好指標[39—40]。河北壩上地區樟子松徑向生長與上一年9月至當年10月PDSI呈顯著正相關。干旱程度增大會抑制樟子松的生長,當樟子松遭遇干旱脅迫時,葉片氣孔關閉會阻礙樹木與外界進行氣體交換,減緩光合作用,樹木體內碳水化合物缺乏,呼吸作用和新陳代謝會消耗體內營養物質,使其處于饑餓的狀態,極易形成窄輪[41]。
3.3 河北壩上樟子松的生態彈性
本研究中隨樟子松樹齡的增加,樟子松的抵抗力呈現下降趨勢,而恢復力出現上升趨勢,在遭遇干旱時樹木的恢復力和抵抗力存在一種平衡關系,抵抗力較弱的區域在干旱后會有較強的恢復力[42]。近30年來,河北壩上地區溫度上升明顯,有效土壤水分利用率呈降低的趨勢(圖3),該地區易發生干旱事件,如果水分可用性的變化超過閾值,光合作用減弱,導致碳水化合物減少,抵抗力減弱,樹木無法抵抗干旱將面臨生長下降的風險,在抵抗力弱的地區干旱極易引發樹木生長的衰退[43]。此外,如果發生持續的干旱,它們可能持續影響樹木的生長,導致樹木生長下降的風險增加[44]。當抵抗力減小到一定程度后,其恢復力可能會減小,在評估樟子松在河北壩上生長適應性時要綜合評估抵抗力和恢復力以及影響它們的因素,充分考慮降水和土壤水分對樟子松的影響[45]。
4 結論
河北壩上地區樟子松差值年表有效長度為1992—2019年,根據其生長過程將其劃分為3個階段,1—10 a的快速生長期,11—20 a的生長下降期以及21—28 a的生長平穩期。樟子松胸高斷面積增量在2000年后呈現平穩特征,樟子松對河北壩上地區的干旱環境有一定的適應性。樟子松徑向生長主要與平均溫度和最高溫度呈顯著負相關,其中最高溫度對其影響最顯著。樟子松徑向生長主要與上一年9月和當年生長季(5、7月)的降水顯著正相關,適量的降水促進河北壩上地區樟子松的生長。干旱脅迫會抑制樟子松的徑向生長,干旱指數越小說明干旱程度越嚴重,樟子松與多月份(上一年9月—當年10月)的干旱指數呈顯著正相關。隨著氣候變暖,當地暖干化特征顯著,樟子松的抵抗力下降,恢復力上升,若持續發生干旱,樟子松生長下降的風險增加,易引發樹木的衰退。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
