大白菜CMS 不育類型的鑒定及細胞學觀察
2022-04-06 07:17:10王曉玲李慶飛原讓花王夢夢董彥琪常懷成張倩倩
中國瓜菜 2022年3期
王曉玲,李慶飛,原讓花,王夢夢,肖 艷,董彥琪,常懷成,張倩倩
(1.河南省新鄉市農業科學院 河南新鄉 453000; 2.河南科技學院園藝園林學院 河南新鄉 453003)
在十字花科作物大白菜的雜交優勢育種中,利用其雄性不育系雜交育種是一條非常重要的途徑。雄性不育性在高等植物中普遍存在,雄性不育可以分為細胞質不育型、細胞核不育型和核質互作不育型3 種類型。由于大白菜資源中沒有細胞質雄性不育源,生產中使用的細胞質雄性不育系均來源于異源細胞質。細胞質雄性不育系主要有3種類型:蘿卜Ogura CMS、甘藍型油菜Polima CMS和甘藍型油菜nap CMS。雄性不育系的花粉由于受多種因素的影響,花粉不能正常的發育或者沒有生活力,雄蕊的敗育會因品種或類型的不同而有所差異,不同大白菜材料的CMS 和保持系的線粒體DNA 的RFLP、RAPD 和AFLP 呈多態性,CMS 和保持系的線粒體基因組結構也存在著顯著差異。產生細胞質雄性不育系的分子機制主要有兩種:一是線粒體DNA 產生新的閱讀框(open reading fragment,ORF),產生有毒的蛋白,從而影響到線粒體的正常功能;二是與線粒體雄性不育有關的基因與正常線粒體基因重組或共表達,直接或間接影響正常基因的功能,產生雄性不育。如在Ogura 型蘿卜中發現的orf138、nap 甘藍型油菜中發現的orf222、Polima 型油菜中發現的orf224、葉用芥菜中發現的orf220等都是在不育系材料中發現的與不育性相關的開放閱讀框。
細胞和組織在離開機體后很快會發生溶解和腐敗,很難維持它原有的形態和結構,因此,通過固定、脫水、透明、浸蠟、包埋、切片、脫蠟、染色等步驟制作石蠟切片可免使細胞組織死去,以便于后期進行觀察分析。
本試驗在以往研究的基礎上,探求大白菜細胞質雄性不育發生的時期及其特點,對現有的不育系材料進行鑒定分類和細胞學觀察。采用同源序列基因擴增技術,依據與細胞質雄性不育有關的線粒體嵌合基因orf224、orf138、orf222 設計出3 對引物,對3種大白菜花蕾粒線粒體DNA進行PCR擴增,從而獲得差異片段,并通過電泳試驗對大白菜細胞質雄性不育系的類型進行鑒定;繼而采用石蠟切片的方法,用光學顯微鏡觀察花蕾在不同長度時花藥的發育程度。為細胞質雄性不育系在大白菜新品種選育中的應用奠定理論基礎。
1 材料與方法
1.1 材料
本試驗采用3個不育系及相對應的保持系(表1)。

表1 6 個大白菜花蕾樣本的編號、名稱、類型及來源
1.2 器材與試劑
器材:光學顯微鏡、真空抽氣機、恒溫箱、蠟片切片機、相機、溫臺、展片臺、蠟杯、酒精燈、載玻片、蓋玻片、毛筆。
試劑:FAA固定液、乙醇溶液、醋酸、二甲苯、甲醛、伊紅、粘片劑、石蠟、染色液(固綠、番紅)、加拿大樹膠。
1.3 方法
于2019 年12 月10 日進行冬季育苗,2020 年2月25 日定植于大棚,每份材料種植40 株,2020 年4月20 日在新鄉市農業科學院研發中心試驗大棚進行采樣,并在此研發中心實驗室進行室內試驗。采樣前注意先將塑料袋和鑷子在液氮中預冷,用鑷子將大白菜花蕾粒輕輕采下放入采樣袋中。根據試驗田種植的材料,取配對的3 個不育源,3 個與其對應的保持系。在種植的植株上,完全隨機選擇花序,每個材料取頂部花蕾3~6個,取樣完畢后將樣品放入液氮罐中,于超低溫冰箱-80 ℃冷凍儲存。不育系類型的鑒定和細胞學觀察于河南科技學院園藝園林學院分子實驗室進行,采用混合樣,無重復。
1.3.1 大白菜細胞質雄性不育系類型的鑒定 在NCBI中檢索的細胞質雄性不育有關的線粒體嵌合基因orf224(DQ400846.1)、orf138、orf222,利用生物學軟件primer5軟件設計出3對引物,由上海生物技術生物工程有限公司進行合成。使用改進的CTAB 方法分別提取6 個大白菜花蕾粒樣本花蕾DNA;并對其進行PCR 擴增,擴增產物經2.0%瓊脂糖凝膠電泳檢測,鑒定其細胞質雄性不育系的不育類型。
1.3.2 大白菜細胞質雄性不育系的細胞學觀察 于2020 年5 月6 日9:00—10:00 采取長度分別為(L≤1 mm、1 mm<L≤2 mm、2 mm<L≤3 mm,L 代表花蕾的長度)新鮮大白菜花蕾,此時植物細胞分裂活動明顯,取材效果最好。為了維持大白菜細胞的自然狀態,使形態和結構不發生變化,取材后參照段國祿等的固定方法馬上進行固定,參考何艷艷、張寶華的方法制作石蠟切片。在光學顯微鏡(XSP-8C,上海精密儀器儀表有限公司)下觀察切片,并進行拍照記錄。
2 結果與分析
2.1 引物設計結果
從NCBI 獲得線粒體嵌合基因orf138、orf224、orf222的序列,并通過Primer 5軟件設計引物(表2)。

表2 設計引物的名稱及序列
2.2 CMS類型的鑒定
用設計的orf138 引物、orf224 引物和orf222 引物對3 個大白菜細胞質雄性不育系及其保持系的6份花蕾粒DNA 進行擴增。結果如圖1 所示,orf138引物擴增出的條帶2 清晰可見,即Ⅲ(紫不育系)為Ogura 型雄性不育系類型,ⅢB 為紫保持系;orf224引物擴增出的條帶4和條帶6很清晰,即Ⅰ(晉毛不育系)、Ⅱ(苔不育系)為Polima型雄性不育系類型;條帶5(ⅡB)、條帶7(ⅠB)分別為苔保持系、晉毛保持系。orf222引物未擴增出條帶,說明Ⅰ(晉毛不育系)、Ⅱ(苔不育系)、Ⅲ(紫不育系)都不屬于nap 型雄性不育系類型;orf224、orf138 和orf222 對保持系材料擴增條帶均不清晰,即基因序列中沒有與細胞質雄性不育相關的開放閱讀框序列。試驗結果表明:Ⅰ(晉毛不育系)、Ⅱ(苔不育系)為Polima 型雄性不育系類型,Ⅲ(紫不育系)為Ogura 型雄性不育系類型;Ⅰ(晉毛不育系)、Ⅱ(苔不育系)、Ⅲ(紫不育系)都不屬于nap型雄性不育系類型。
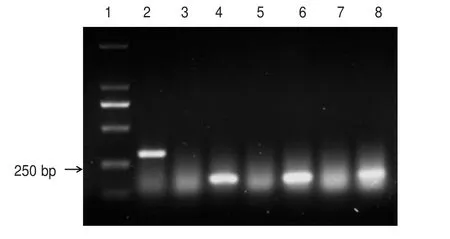
圖1 orf224、orf138、orf222 引物PCR 擴增產物電泳圖譜
2.3 保持系材料和不育系材料花藥發育的特點
大白菜正常的花藥由4 個藥室組成,藥室由外到內分別是表皮、纖維層、中層、絨氈層。絨氈層呈長方形規則緊密地排列在藥室最里層,主要為花粉母細胞提供營養,隨著花粉發育會逐漸降解,花藥同一側2 個相鄰小孢子囊之間的組織也會逐漸消失,打通變成1個藥室。通過切片觀察,可以看到保持系材料IB 晉毛保持系、IIB 苔保持系和IIIB 紫保持系的花藥在各個時期都發育正常(圖2~4)。
大白菜CMS不育主要是雄蕊發育不正常,不能產生可育的花粉。通過切片觀察可以看到,不育系材料中大多在花蕾長度≤1 mm 時就可以明顯看到差異,而后期本該形成藥室的地方只存在大量薄壁細胞。個別藥室可以形成花粉囊產生花粉,但在后期因花粉延遲發育或粘連在一起無法正常散粉。此外,還出現個別花藥有多藥隔、無藥隔情況(圖2~4)。
2.4 保持系材料和不育系材料花藥發育的區別
根據對保持系材料與不育材料花藥切片的細胞學觀察,在花蕾長度小于等于1 mm時,保持系材料IB 晉毛保持系(圖2-2)、IIB 苔保持系(圖3-2)、IIIB紫保持系(圖4-2)在花蕾的橫切面上,可見1個雌蕊和4個雄蕊,花藥呈蝶形,4個藥室已經形成,可以看見絨粘層在花粉囊內染色較深,且維管束和花粉囊內細胞均被染色。而I晉毛不育系(圖2-1)和II苔不育系(圖3-1)的4個花粉室尚未形成,III紫不育系(圖4-1)可以看見有花粉囊的形成,但染色較淺,沒有產生明顯的花藥。
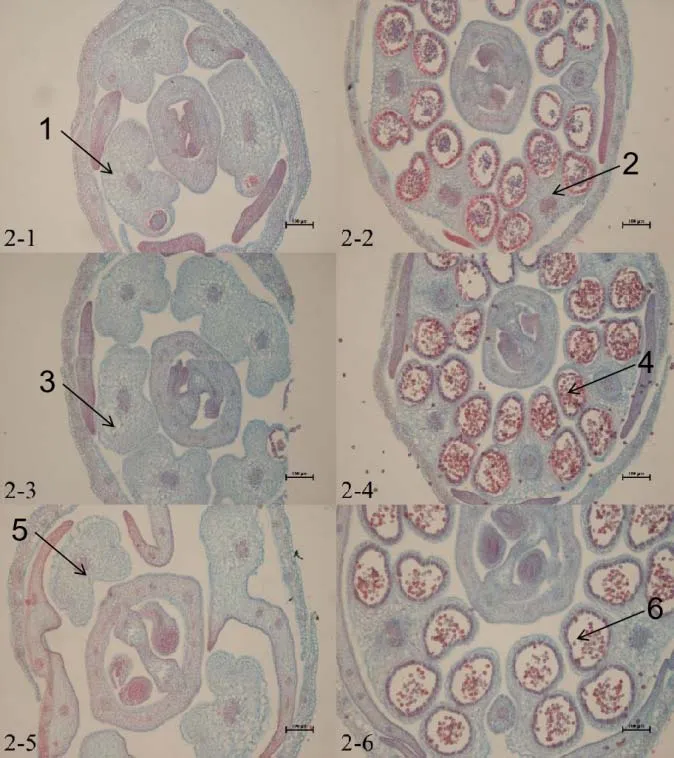
圖2 I 晉毛不育系和IB 晉毛保持系細胞學觀察
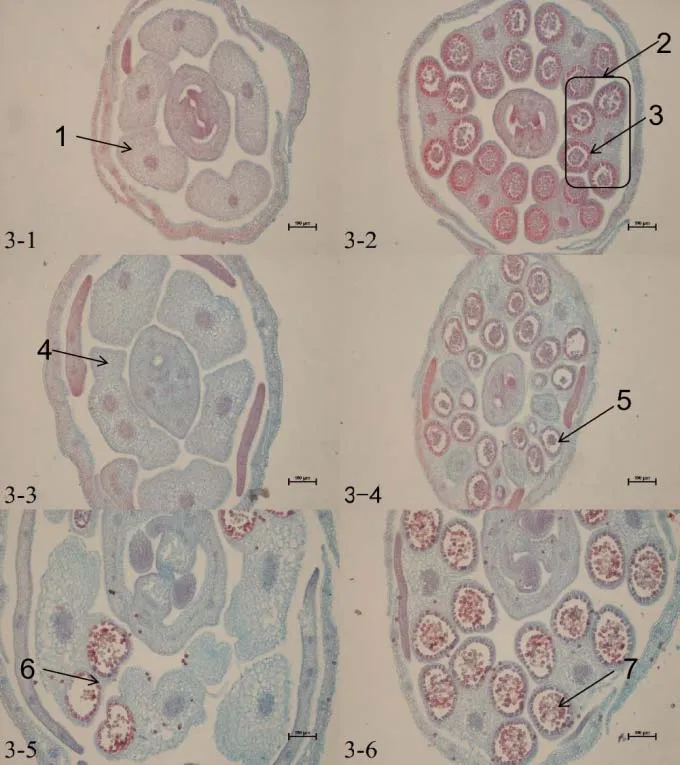
圖3 II 苔不育系和IIB 苔保持系細胞學觀察
當花蕾長度為1 mm <L≤2 mm 時,保持系材料花藥中藥室、藥隔更加清晰,絨粘層整齊的排列在小孢子囊母細胞的周圍,可以看見清楚的藥室由外到內分別為表皮、藥室的內壁、中層、絨氈層(圖2-4,3-4,4-4)。但不育系I 晉毛不育系(圖2-3)和II苔不育系(圖3-3)在這個時期仍然沒有形成花粉囊,III 紫不育系雖然有花粉囊的形成,但花粉囊內的花粉較少,個別花藥內不能正常形成4個藥室,還有個別花粉囊內絨氈層膨大而擠壓小孢子形成一條縫隙,造成小孢子發育不良,呈現敗育的現象(圖4-3)。當花蕾長度為2 mm<L≤3 mm 時,保持系材料孢間層逐漸溶解,細胞逐漸分開,變成圓球形,個別細胞呈四邊體形,著色較深(圖2-6,3-6,4-6)。不育系材料I 晉毛不育系雌蕊發育正常,從此時雌蕊發育的程度以及IB晉毛保持系的發育程度來看,花粉發育應該和它的保持系發育一樣,但此時I 晉毛不育系仍然沒有藥室形成,細胞呈半透明狀,是典型的無花粉囊(圖2-5)。II苔不育系發育較晚,只有角隅處可以形成2~3 個藥室,而且花蕾橫切面不能呈正常的蝶形(圖3-5)。而可以觀察到III紫不育系花粉囊內的細胞染色較淺,花粉囊內的花粉粘連無法形成單個的圓形花粉粒,個別出現無藥隔的現象(圖4-5)。
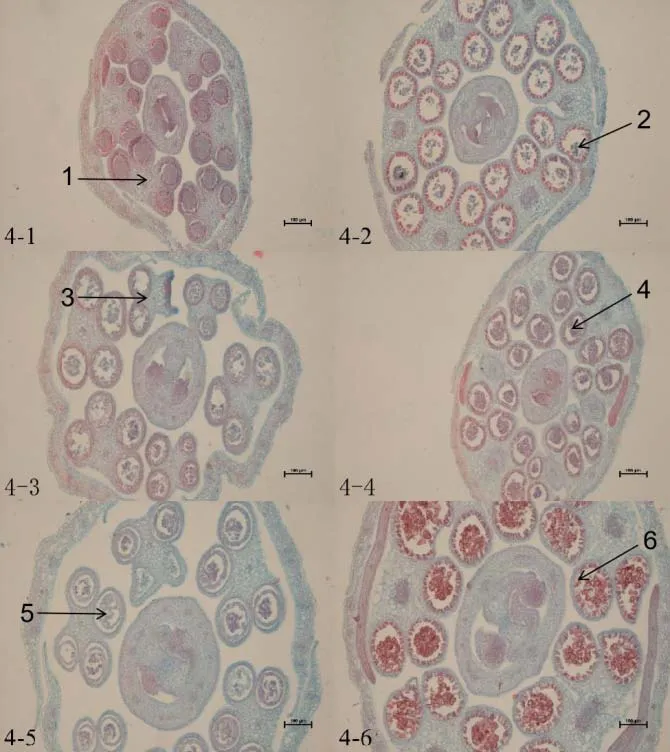
圖4 III 紫不育系和IIIB 紫保持系細胞學觀察
3 討論與結論
近20年來,在大白菜細胞質雄性不育性研究方面,已報道有Ogura CMS、Polima CMS 和nap CMS等多種細胞質雄性不育系類型。Polima CMS 是最有生產價值的細胞質雄性不育系,不僅在大白菜CMS不育系的選育中被廣泛利用,而且在其他十字花科作物雄性不育系的選育中也被廣泛應用。2005 年趙利民建立了一套大白菜新品種灰色關聯度多維綜合評判方法,為客觀、準確地從眾多大白菜組合中篩選出優良新品種提供了科學依據,提高了新品種選育的準確性和選擇效率。2018 年崔冰等對大白菜細胞質雄性不育形成的機制進行研究。2020 年楊紅麗等培育出耐熱抗根腫病蘿卜胞質雄性不育多基因聚合新品種。隨著科學技術的發展,準確快速鑒定樣本所歸屬的雄性不育系的類型,以便為開展細胞質雄性不育系在育種中的應用奠定基礎,并為實現大白菜優勢雜交育種提供分子依據。筆者對6 個大白菜材料CMS 的不育類型進行鑒定,發現其中Ⅰ晉毛不育系、Ⅱ苔不育系為Polima 型雄性不育系類型;Ⅲ紫不育系為Ogura 型雄性不育系類型;因其基因序列中沒有與本試驗CMS 不育相關的開放閱讀框序列,所以orf224、orf138和orf222引物均未擴增出清晰條帶。
通過石蠟切片法進行觀察發現本試驗中大白菜保持系花藥均發育正常,可以形成4 個蝶形藥室從而產生可育的花粉。I晉毛不育系與可育材料IB晉毛保持系比較,I 晉毛不育系的敗育花藥一直處于孢原細胞之前,在整個時期內都沒有形成花粉囊,屬于典型的無花粉囊型敗育,它的藥室異常出現較早,當花蕾長度為L≤1 mm時差異顯著;II苔不育系藥室異常出現較晚,在花蕾長度為1~2 mm 時還未形成花粉囊,發育較晚,在花蕾長度為2~3 mm時只有2~3個花粉囊可以形成,且花藥畸形,無法形成正常的蝶形藥室,個別花藥內還出現多藥隔現象;III 紫不育系,在后期可以明顯地觀察到不育系花粉囊內的花粉因相互粘連無法形成單個的花粉粒。2016 年郭瑛琪等大白菜核不育花藥敗育起始于減數分裂期絨氈層結構異常,小孢子在四分體時期開始敗育;不育發生后,伴隨著絨氈層細胞異常膨大,嚴重空泡化,提前降解,小孢子變形干癟而死亡。2012 年許明等研究表明不育花蕾長度在2.5 mm后就已經敗育,敗育時期可能發生在減數分裂后期到四分體時期。本試驗與其研究結論一致,花蕾中花粉敗育的時期較早,當L≤1 mm時,它就決定了花藥能否形成花粉囊從而產生花藥。
