新疆克孜爾河斯氏高原鰍繁殖生物學初步研究
2022-04-13 07:47:42李麗朱霞強壯趙賀魏杰
塔里木大學學報 2022年1期
李麗,朱霞 ,強壯,趙賀,魏杰*
(1塔里木大學生命科學與技術學院,新疆 阿拉爾 843300)
(2塔里木畜牧科技兵團重點實驗室,新疆 阿拉爾 843300)
(3塔里木大學動物科學與技術學院,新疆 阿拉爾 843300)
繁殖是魚類養殖及遺傳育種的重要環節,是魚類生命過程中不可或缺的部分,繁殖生物學是魚類延續種群和生長進化的重要組成部分[1]。魚類繁殖生物學特性的形成主要與其自身遺傳因素和外界環境因素有關[2]。這都是在長期種群進化過程中自然選擇的結果[3-4],它顯示出種群及其后代對環境的適應特征[5],然而這種適應特征與魚類的性腺發育狀況[6]、性成熟時間[7]、產卵類型、產卵場所等息息相關[8]。
斯氏高原鰍(Triplophysa stoliczkae),別名背斑條鰍,隸屬于鯉形目(Cypriniformes)、鰍科(Cobitidae)、條鰍亞科(Noemacheilinae)、高原鰍屬(Triplophysa)、高原鰍亞屬(Triplophysa)[9],是克孜爾河土著魚類。目前對斯氏高原鰍的研究僅限于分布[9]和寄生蟲[10]等方面,對其繁殖生物學的研究尚未見報道。近年來陳生熬[11]、梁祥等[7]、王志堅等[8]、汪帆等[12]學者分別對葉爾羌高原鰍(Triplophysa Yarkandensis)、秀麗高原鰍(Triplophysa venusta)、貝氏高原鰍(Triplophysa bleekeri)、東方高原鰍(Triplophysa orientalis)的繁殖生物學進行研究,對了解高原鰍屬魚類早期生活史,開展其增殖保護等工作提供了基本參考資料。
本研究通過重量比例法估算斯氏高原鰍的個體繁殖力,測定性比和最小性成熟個體,觀察其第二性征和性腺發育時期,并統計卵徑分布確定其產卵類型,旨在探明克孜爾河斯氏高原鰍的繁殖生物學特性,豐富新疆土著魚類繁殖生物學的內容,為其繁殖及增殖放流等工作提供基礎數據。
1 材料與方法
1.1 材料
斯氏高原鰍于2018年6月采集于新疆拜城縣克孜爾河黑英山鄉河段,并對其生存水域的水溫和流速進行現場測量(表1)。捕獲斯氏高原鰍成魚30尾,測定其體長、體質量以及性腺重(表2),將樣本用10%的福爾馬林固定帶回實驗室進一步研究[13]。

表1 黑英山鄉河段水溫、流速指標

表2 斯氏高原鰍成魚常規形態指標
1.2 方法
1.2.1 雌雄差異
對性成熟樣本進行雌雄辨別,先從胸鰭的形狀、體色深淺、背部斑紋分界的清晰度與否以及腹部膨大程度進行區別,然后再從第一性征上進行驗證。
1.2.2 性比
通過解剖,根據其第一性征鑒別雌雄,并統計雌雄性比。
1.2.3 個體繁殖力
參考《魚類生態學》[14],按照重量比例法來計算斯氏高原鰍的個體繁殖力。選取處于性腺發育至Ⅳ期的10尾雌魚卵巢,測量卵巢總重,然后分別在卵巢前部、中部、后部取0.1 g進行卵粒計數。絕對繁殖力、相對繁殖力計算方法如下:
絕對繁殖力=(樣品卵粒數/樣品重)×卵巢重
相對繁殖力=絕對繁殖力/空殼重
1.2.4 卵徑分布和產卵類型
隨機取卵觀察測量,用吸管隨機吸取處于Ⅳ期的10尾雌魚卵巢,每尾隨機吸取10粒卵粒,共100粒。在體視顯微鏡下測量其卵徑,每個樣本測100粒,共1 000粒。根據結果制作卵徑分布圖,用卵徑頻數分布法和實驗觀察法來判斷產卵類型。卵徑分布圖呈單峰型,則為一次產卵類型;如為雙峰,則為分批產卵類型;如為多峰,則為連續產卵類型[15]。
1.2.5 最小性成熟個體
從性腺發育至Ⅳ期的個體中挑選出體長、體重最小的個體,再結合年齡數據綜合所得。
1.3 數據分析
數據使用SPSS 26.0軟件進行單因素方差分析,采用Duncan方法進行多重比較,顯著性水平設為0.05。卵徑的數據以平均數±標準差(mean±SD)來表示。
2 結果
2.1 雌雄差異
2.1.1 第一性征
對25尾斯氏高原鰍進行解剖,結果顯示雌魚發育至Ⅳ期的卵巢左右連成一片,近似一個長袋狀,能看到大部分已沉積了卵黃的卵粒,見圖1A。雄魚發育至Ⅳ期的精巢,呈白色不規則曲折狀,近似Z字形彎曲,見圖1B。

圖1 發育至Ⅳ期的斯氏高原鰍性腺比較
通過解剖觀察到卵巢發育至Ⅲ期的2尾,發育至Ⅳ期的10尾;精巢發育至Ⅱ期的1尾,Ⅲ期的2尾,Ⅳ期的10尾。處于Ⅲ期的卵巢,透過卵膜肉眼可見卵粒,只有一小部分卵粒能從卵膜上分離下來,大部分都不能分離,且卵粒開始沉積卵黃,能看見卵巢表面的血管。Ⅳ期卵巢體積增大,占據腹腔的1/3左右,呈淡黃色,卵粒相對較大,沉積卵黃的卵粒數比Ⅲ期多,處于Ⅱ期的精巢不透明,細帶狀。Ⅲ期的精巢呈彎曲長條狀,整體呈白色,Ⅳ期的精巢乳白色呈Z字形彎曲。
2.1.2 第二性征
對性腺發育至Ⅳ期的斯氏高原鰍的雌雄差異進行比較,結果如圖2所示。雄魚胸鰭比其他鰭略厚,胸鰭外側第一或第二根鰭條變硬;雌性胸鰭無明顯變化。雄魚的胸鰭略尖,雌魚的胸鰭鈍圓。性成熟雄魚體色較深,背部花紋呈馬鞍型,分界明顯;雌魚體色較淡,身體長形,背部斑紋分界明顯。由于生殖季節的來臨,雌魚的卵巢逐步發育表現出腹部膨大,個體較為豐滿,但此時相比于雄魚,其個體相對偏瘦,且腹部膨大不明顯。

圖2 發育至Ⅳ期的斯氏高原鰍雌雄差異比較
2.2 性比
30尾斯氏高原鰍中雌魚12尾,雄魚13尾,剩余5尾未辨別出雌雄。性比值為1.00:1.08(N=25)。
2.3 繁殖力
選卵巢發育于Ⅳ期的雌魚,計算斯氏高原鰍個體繁殖力,結果顯示絕對繁殖力范圍為964~4 095粒,平均為2 361粒,相對繁殖力范圍為172~493粒/g,平均為301粒/g。
2.4 卵徑分布和產卵類型
測量10尾雌魚性腺發育至Ⅳ期卵巢中卵的卵徑,依據測量結果作出卵徑分布圖,結果如圖3所示。本次共測量1 000粒卵的卵徑,卵徑均值為(0.15±0.03)mm,屬于卵徑小的魚類。卵徑處于0.13~0.15 mm居多,為優勢組,占總測量卵量的40%,其他組共占60%。因圖中顯示只有一個峰值,且卵巢中的卵屬于同一時期性成熟,推測為一次產卵類型。
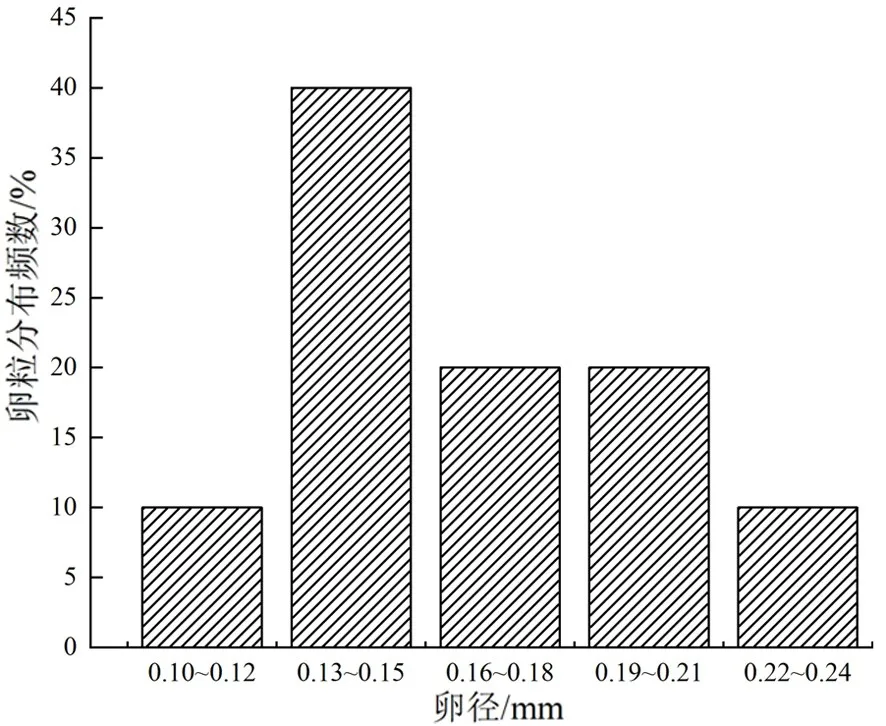
圖3 斯氏高原鰍Ⅳ期卵巢中卵徑分布圖
2.5 最小性成熟個體
斯氏高原鰍雌魚和雄魚的最小性成熟個體標準如圖4所示,雌魚的最小性成熟個體年齡為2齡,體長46.19 mm,體質量1.02 g;雄魚的最小成熟個體年齡為2齡,體長49.55 mm,體質量1.16 g。

圖4 斯氏高原鰍最小性成熟個體
3 討論
3.1 第二性征
一般來說,當達到性成熟之后,在外部形態上顯著發現雌魚較雄魚整體上飽滿,在解剖過程中也發現由于魚體內臟中存在脂肪積累過多而導致其體型飽滿的狀況。并且在生殖季節大多數鯉科和鰍科魚類的第二性征會在吻端、鰓蓋和胸鰭等部位出現珠星,對比發現高原鰍屬魚類性成熟時只有東方高原鰍胸鰭上出現珠星[8,16]。在本研究中,斯氏高原鰍性成熟個體未發現珠星的出現,同時在雄魚的頰部和胸鰭上也均未發現有肉墊或刺突。繁殖季節雄性胸鰭顯著增厚,雄性胸鰭外側第一或二根鰭條變硬,與賽里木湖高原鰍的研究結果一致[17]。斯氏高原鰍雄魚胸鰭略尖,雌魚胸鰭鈍圓,與葉爾羌高原鰍的研究結果一致[2]。雌雄體色深淺這一特征與同屬于鰍科的東方高原鰍和中華沙鰍描述相符[8,18]。對6種不同流域高原鰍第二性征的比較見表3。結果發現,生活在不同水系中的相同種類之間雄性第二性征也不盡相同,這表明外界生活環境也會影響到高原鰍的第二性征。

表3 6種不同流域高原鰍魚類第二性征的比較
3.2 個體繁殖力
斯氏高原鰍相對繁殖力的平均值為301粒/g,而同屬于高原鰍屬的賽里木湖的新疆高原鰍(Triplophysa strauchii)其相對繁殖力的平均值為568粒/g[16],柴窩堡的新疆高原鰍其相對繁殖力的平均值為408粒/g[21],秀麗高原鰍其相對繁殖力的平均值為420粒/g[22],葉爾羌高原鰍其相對繁殖力的平均值為982粒/g[2],相比較而言斯氏高原鰍相對繁殖力水平最低。一方面,可能是由于其長期適應外界環境的結果,克孜爾河發源于天山南部的汗騰格里峰,主要以冰雪融水補給[23],流速較大,水質偏瘦,營養物質較少;另一方面這也可能是由其遺傳特性所決定,具體原因還有待進一步探究。
3.3 產卵類型
產卵類型的確定一般會采用性腺組織學觀察法、實驗觀察法、成熟系數(GSI)周年變化法、卵徑頻率分布法綜合判斷來確定[14]。本研究發現,黑英山河段的斯氏高原鰍卵巢中的卵多數是屬于同一時期發育成熟且卵徑分布圖呈單峰,判斷為一次產卵類型,該結果與葉爾羌高原鰍的研究結果一致[2],每年的6月份是其產卵繁殖高峰期。但同屬于高原鰍屬的貝氏高原鰍,其一年產卵兩次[12](卵巢中的卵不是一起成熟,背部和尾部先成熟,腹部后成熟);秀麗高原鰍、東方高原鰍都為分批產卵類型[7-8],同屬鯉形目的裂腹魚類,如昆明裂腹魚(Schizdthorax grahami)[24]、神農架齊口裂腹魚(Schizothorax prenati)[19],其產卵類型均為單峰一次產卵類型。
3.4 最小性成熟個體
魚類種間自身的遺傳因素及其所生活的外界環境是影響魚類性腺成熟的兩大主要原因[22]。因此,對于特定的物種來說,研究魚類的性成熟,一方面要考慮遺傳的穩定性,另一方面還要考慮外界因素中的不確定性。外界因素在一定程度上起到決定性作用,魚類之間身體上的相互碰撞、無意間的撞擊石頭以及水質的污染等均會不同程度的影響最小性成熟個體[25]。本研究中,斯氏高原鰍雌雄最小性成熟個體年齡均為2齡,這與秀麗高原鰍和大寧河貝氏高原鰍最小性成熟年齡均為2齡的研究結果一致[12,22],且都屬于同齡性成熟。而在其它研究中,很多魚類則不屬于同齡性成熟,如葉爾羌高原鰍雌性最小性成熟年齡為3齡,雄性為2齡[2],神農架齊口裂腹魚雌性最小性成熟年齡為4齡,雄性為3齡[19];金沙江圓口銅魚(Coreius guichenoti)雌性最小性成熟年齡雌為4齡,雄性為3齡[20]。推測魚類性成熟是否同齡可能與魚類自身遺傳因素有關,同時也與環境中餌料種類及豐度、過渡捕撈以及水利工程建設等密切相關[26-27]。
4 結論
經研究發現,斯氏高原鰍在性成熟時雄魚體色較深,輕壓腹部有精液擠出;雌魚體色較淡,腹部隆起,生殖孔微凸。除5尾未辨性別外,雌雄魚性比為1.00:1.08(N=25)。雌魚雄魚最小性成熟年齡均為2齡,雌性最小性成熟個體體長46.19 mm,體質量1.02 g;雄性最小性成熟個體體長49.55 mm,體質量1.16 g。斯氏高原鰍屬于一次產卵類型,其絕對繁殖力為(2 362±1 066)粒,相對繁殖力為(301±112)粒/g。該魚繁殖力較低,資源瀕危,因此,開展對斯氏高原鰍的基礎研究和人工繁殖并進行增殖放流,恢復其資源,尤為重要且迫在眉睫。
