長江源區高寒沼澤草甸植物對增溫的生理響應及適應性評價
2022-04-25 12:40:10張景然房佳辰曾紅麗
草地學報 2022年4期
關鍵詞:植物
馮 月, 白 煒, 張景然, 房佳辰, 曾紅麗
(蘭州交通大學環境與市政工程學院, 甘肅 蘭州 730070)
全球變暖正在改變陸地生態系統的結構和功能,造成的負面和潛在影響已受到世界各國政府和科學家的重點關注。IPCC報告表明,近百年來全球平均地表溫度升高了0.85℃,并預測到2100年將升高0.3~4.8℃,高緯度或高海拔地區的增幅更明顯[1-2]。青藏高原平均海拔超過4 000 m,自然條件嚴酷,生態系統極為脆弱,極易受到氣候變暖和人類活動的影響。青藏高原因自身獨特的自然環境特征被稱為全球氣候變化的觸發器,其隆起和抬升形成了獨特的高原季風系統,并對周邊地區乃至全球的氣候具有強烈影響[3-4]。溫度作為高寒植物生長的主要限制性因子[5],其變化會對多年凍土、活動層土壤水分狀態等造成強烈影響,使得地氣間雪蓋層和植被的水熱平衡發生變化,引起植物生理特性的改變[6],進而影響植物生長,推動植物群落和物種組成發生變化[7],影響高寒生態系統的穩定性[8-9]。沼澤化草甸是一個非常獨特的生態系統,以莎草科等植物為優勢種,涵蓋濕地和草原所具有的結構和功能,是青藏高原分布較為廣泛的草地生態系統[10]。但隨著氣候變暖程度加重,青藏高原草地生態系統的空間分布格局也隨之變化,表現為高寒草原分布面積增加,而高寒沼澤草甸顯著退化[11],因此研究高寒沼澤生態系統對氣溫升高響應特征及其內在機制具有重要的獨特性和前瞻性。
目前,針對本研究關注的沼澤草甸生態系統,相關研究多集中在溫度變化對沼澤草甸植物群落組成[12-13]、物種多樣性[14-15]和生物量[16-18]等方面的影響,而對沼澤草甸植物生理響應關注較少。僅對群落結構和功能等宏觀方面進行研究,很難全面地揭示高寒沼澤草甸適應增溫環境的策略,有必要對增溫條件下高寒沼澤草甸植物生理響應進行研究,以便更好地理解高寒沼澤草甸適應增溫的機制和策略。如石福孫等[19]的研究,認為可通過生理指標定量分析判斷升溫是否對高寒植物生長造成負面影響;付剛等[6]發現高寒植物生理變化決定該區域植被群落結構和生產力演變的方向和程度;喬曉英等[20]認為葉綠素、丙二醛等生理指標含量的高低可指示寸草苔(Carexduriuscula)對環境變化適應能力的強弱。光合色素、滲透調節物質、丙二醛的含量及抗氧化酶活性往往會隨著增溫程度的變化而相應變化,因而成為研究增溫條件下植物生理響應的重要指標[17]。
植物對增溫適應能力的強弱是受多因子影響的綜合表現,簡單的使用單項生理指標難以全面準確地反映各物種的適應能力[21-22]。現常用多元統計分析方法對植物的增溫適應性進行評價,主要采用主成分分析法結合其他數據統計方法,如Logistic回歸分析法、隸屬函數分析法及聚類分析法等。該評價方法目前已經廣泛應用于芍藥屬(Paeonia)等多種植物的對增溫適應性評價,且多元建模分析技術的評價結果與實際熱害觀察結果一致[23]。因此,本研究采用OTCs系統模擬增溫的方式,分析青藏高原高寒沼澤草甸植物矮嵩草(Kobresiahumilis)、藏嵩草(Kobresiatibetica)和青藏苔草(Carexmoorcroftii)光合色素含量、滲透調節物質、丙二醛含量和抗氧化酶活性對短期增溫的響應特征,并運用主成分分析法及隸屬函數法評價3種植物適應性,揭示3種植物對氣溫升高的響應模式及差異,為預測未來氣候變化對高寒沼澤草甸植物以及整個陸地生態系統產生的影響奠定理論基礎。
1 材料與方法
1.1 研究區概況
研究區域位于青藏高原腹地風火山地區,海拔為4 680~5 360 m,屬于典型多年凍土區,該區年均降水量270 mm,年均蒸發量1 478 mm,且降水主要發生在植物生長季(5—9月)[6]。該區域面積約30%為高寒沼澤草甸,主要分布在海拔3 200~4 800 m的河畔、湖濱、排水不暢的平緩灘地[16],本研究中高寒沼澤草甸樣地位于河畔,坐標為 34°43′57.5″ N,92°53′22.5″ E,海拔4759 m,群落覆蓋度大、物種組成豐富。主要植物種有矮嵩草、藏嵩草、青藏苔草、針茅(Stipaaliena)、羊茅(Festucasp.)等高寒植物,植被蓋度達85%[24]。
1.2 研究方法
2015年7月,分別選取植被蓋度、植物類型以及地勢基本一致的試驗樣地進行試驗布置,建立100 m×100 m樣地,并于樣地內隨機建立5個試驗小區,試驗小區間隔不小于6 m。在小區內隨機布設對照樣方(1.5 m×1.5 m)、低幅度增溫樣方(T1,增溫1.5~2.5℃)和高幅度增溫樣方(T2,增溫3~5℃)各1個,對照和增溫樣方相隔2~3 m,共計15個的樣方。
模擬增溫的方式采用國際凍原計劃中的開頂式生長室(open-top chambers,OTCs)[25]。增溫室采用透明有機玻璃加工成正六邊形圓臺狀,透光率95%,低幅度增溫和高幅度增溫小室高度分別為0.4 m(T1)和0.8 m(T2),底面積分別約為1 m2和2 m2,圓臺斜邊均與地面呈60°角,并保證圓臺開口為0.6 m(圖1)。同時,采用EC-TM溫濕度傳感儀(Decagon Decices Inc.,Pullman,WA)記錄空氣溫度與濕度。對2019年進行全年氣溫監測得出,OTCs內的平均氣溫分別相比CK提高了2.1℃(T1)和4.4℃(T2),其中7月月平均氣溫為全年最大值(圖1)。

圖1 OTCs示意圖(低幅度增溫)及2019年月平均氣溫
1.3 樣品采集
在模擬增溫4年后采樣,即2019年8月22日,在每個樣方內,按對角線法選取矮嵩草、藏嵩草及青藏苔草各約5 g新鮮植株,齊地面剪下各物種植株,然后將重復樣方所采集的矮嵩草、藏嵩草及青藏苔草分別均勻混合,3種處理得到矮嵩草、藏嵩草及青藏苔草新鮮植株各25 g,冷藏于取樣箱迅速帶回實驗室,保存在液氮中待測,每個指標測定3次重復[26]。
1.4 測定指標
1.4.1植物光合色素含量的測定 葉綠素a、葉綠素b及類胡蘿卜素含量的測定根據Lichtenthaler等[27]的乙醇浸提比色法方法,利用可見光分光光度計分別在663,649和470 nm下測定光吸收值,再根據3種色素在該波長下的吸收系數即可求出其含量(mg·g-1)。
1.4.2植物滲透調節物質的測定 脯氨酸含量采用水浴浸提法脯氨酸含量的測定根據Bates等[28]的方法,用 3%的磺基水楊酸溶液研磨提前游離脯氨酸后,測得其含量(μg·g-1FW)。可溶性糖含量的測定根據Dhindsa等[29]的方法,利用450 nm下的光吸收值求出可溶性糖含量(mmol·L-1)。可溶性蛋白含量的測定根據Bradford等[30]的考馬斯亮藍染色法,根據在595 nm下的光吸收值,對照標準蛋白標準曲線求出可溶性蛋白含量(mg·g-1)。
1.4.3植物丙二醛含量的測定 丙二醛含量的測定根據Dhindsa等[29]的方法,利用532 nm下的光吸收值減去600 nm下最小吸收值求出丙二醛含量(μmol·g-1)。
1.4.4植物抗氧化酶活性的測定 過氧化物酶活性的測定根據Ekmekci等[31]的方法,利用470 nm下光吸收值在3 min內每15 s的變化表示過氧化物酶活性(U·min-1·g-1FW)。超氧化物歧化酶活性的測定根據Becana等[32]的方法,以每毫克蛋白抑制氮藍四唑光化還原的50%作為一個酶活性單位。
1.5 數據處理
使用Microsoft Excel 2010進行數據處理,運用SPSS 20.0軟件進行數據分析。不同處理間各植物生理指標差異采用單因素方差分析(One-way ANOVA)與Duncan’s多重比較法進行顯著性檢驗,利用Origin2018軟件進行繪圖,圖表中數據為平均數±標準差。將主成分分析和隸屬函數結合使用評價植物對增溫的適應性[33]。利用SPSS 20.0對9項生理指標進行主成分分析,合理設置特征值的限定值,保證主成分累積貢獻率超過85%,獲得特征值、貢獻率、累積貢獻率等。計算各綜合指標主成分得分并將其隸屬函數化,結合各綜合指標權重排序,具體公式如下:
主成分得分(Zi)計算公式為:
(1)
(i=1,2,3,…,n)
式中:Zi為各物種第i個主成分得分;ai為第i個主成分的標準化特征向量值;Xi為各單項生理指標的標準化值。
隸屬函數值公式為:
(2)
(i=1,2,3,…,n)
式中:μ(Zi)為各主成分得分的隸屬函數值;Zi max和Zi min分別為各物種第i個主成分得分內的最大值和最小值。
各主成分的權重計算公式為:
(3)
(i=1,2,3,…,n)
式中:wi為各物種第i個主成分權重;Pi為各物種第i個主成分的貢獻率。
各物種適應性評價值(D)計算公式為:
(4)
(i=1,2,3,…,n)
2 結果與分析
2.1 增溫對3種植物光合色素的影響
矮嵩草和青藏苔草葉綠素a含量在低幅度增溫(T1)處理下顯著增加(P<0.05),在高幅度增溫(T2)處理下則顯著減少,相比對照(CK),T1處理下矮嵩草和青藏苔草葉綠素a含量分別增加22.49%和9.55%,T2處理下則分別下降了34.69%和36.26%;藏嵩草在T1和T2處理下葉綠素a含量均顯著高于CK,分別增加29.03%和25.27%(圖2)。

圖2 增溫對3種植物光合色素的影響
矮嵩草和青藏苔草葉綠素b含量在T1處理下無顯著變化(P>0.05),在T2處理下則顯著減少,相比CK,T2處理下矮嵩草和青藏苔草葉綠素b含量分別下降了19.90%和18.18%;藏嵩草在T1和T2處理下葉綠素b含量均無顯著變化。
T1和T2處理下矮嵩草和藏嵩草類胡蘿卜素含量均顯著低于CK,相比CK,T1處理下矮嵩草和藏嵩草類胡蘿卜素含量分別下降了13.30%和26.77%,T2處理下分別下降61.58%和41.73%;青藏苔草類胡蘿卜素含量在T1處理下無顯著變化,T2處理下顯著低于CK,下降了61.97%。
2.2 增溫對3種植物滲透調節物質的影響
矮嵩草和青藏苔草脯氨酸含量在T1處理下均無顯著變化,T2處理下顯著增加,相比CK,矮嵩草和青藏苔草脯氨酸含量分別增加369.84%和181.86%;T1和T2處理下藏嵩草游離脯氨酸含量均顯著高于CK,分別增加27.03%和114.10%(圖3)。
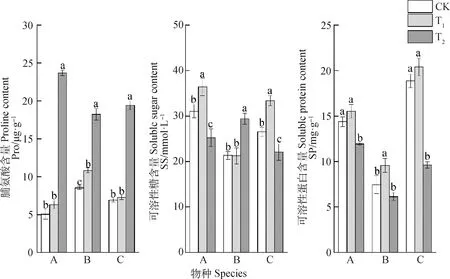
圖3 增溫對3種植物滲透調節物質含量的影響
矮嵩草和青藏苔草可溶性糖含量在T1處理下顯著增加,T2處理下顯著減少,相比CK,T1處理下矮嵩草和青藏苔草可溶性糖含量分別增加了17.26%和25.92%,T2處理下則分別下降了18.84%和16.88%;T1處理下藏嵩草可溶性糖含量無顯著變化,T2處理下則顯著高于CK,增加了37.88%。
矮嵩草和青藏苔草可溶性蛋白含量在T1處理下均無顯著變化,T2處理下則顯著低于CK,分別減少了17.04%和48.99%;藏嵩草在T1處理下可溶性蛋白含量顯著高于CK,增加了28.49%,T2處理下則無顯著變化。
2.3 增溫對3種植物丙二醛含量及抗氧化酶活性影響
矮嵩草在T1和T2處理下丙二醛含量顯著低于CK,分別下降了28.43%和58.61%;藏嵩草在T1處理下丙二醛含量無顯著變化,T2處理下則均顯著低于CK,相比CK,降低了24.05%;青藏苔草在T1處理下丙二醛含量顯著低于CK,相比CK,下降了22.88%,T2處理下則無顯著變化(圖4)。

圖4 增溫對3種植物丙二醛及抗氧化酶活性的影響
矮嵩草在T1和T2處理下過氧化物酶活性均顯著高于CK,分別增加了371.20%和176.48%;藏嵩草則在T1和T2處理下均顯著低于CK,分別減少了46.27%和24.27%;青藏苔草則在T1處理下顯著高于CK,相比CK,增加了105.77%,T2處理下則顯著低于CK,減少了35.26%。
矮嵩草在T1處理下超氧化物歧化酶含量顯著低于CK,減相比CK,下降了6.37%,T2處理下則無顯著變化;藏嵩草在T1處理下顯著高于CK,相比CK,增加了11.74%,T2處理下則顯著減少,下降了6.19%;青藏苔草在T1和T2處理下超氧化物歧化酶含量均顯著高于CK,增加了27.12%和15.46%。
2.4 3種植物增溫適應性評價分析
由表1可知,增溫處理后植物各生理指標之間存在大小不同、顯著性不同的相關性,導致它們所代表的信息存在相互重疊的問題,各單項指標的重要程度也不同。3種植物對增溫的適應性是一個復雜的綜合性狀,從單項指標不能清楚準確地進行適應性評價[34],需要在此基礎上對各指標進行其他多元統計分析。

表1 植物生理指標的相關系數矩陣
由表2可以看出,對9個生理指標進行主成分分析,根據累計貢獻率>85%的原則取前4個主成分進行分析,它們的累計貢獻率為93.6%,表明前4個主成分能代表所有生理指標進行不同物種間對不同增溫幅度的適應性評價。
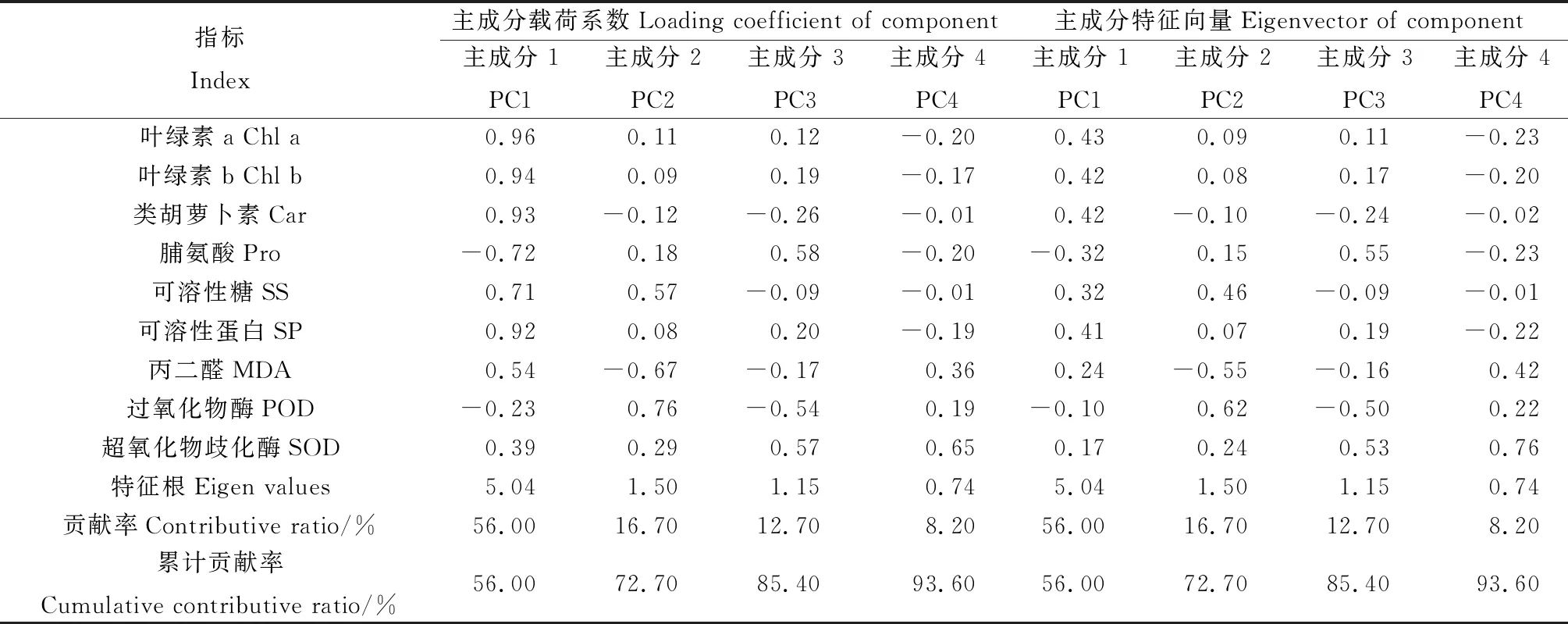
表2 植物生理指標主成分分析
第1主成分主要有葉綠素a、葉綠素b、類胡蘿卜素、脯氨酸和可溶性蛋白,其特征值為5.04,貢獻率為56.0%;第2主成分主要有可溶性糖、丙二醛和過氧化物酶,其特征值為1.50,貢獻率為16.7%;第3主成分主要有脯氨酸和超氧化物歧化酶,其特征值為1.15,貢獻率為12.7%;第4主成分主要有超氧化物歧化酶,其特征值為0.74,貢獻率為8.2%。可見,3種高寒沼澤草甸植物對增溫的適應性受多因子影響,根據各指標在不同因子上的載荷,相關分析確定,葉綠素a、葉綠素b、類胡蘿卜素、可溶性糖、過氧化物酶和超氧化物歧化酶這6個生理指標為增溫處理下反應敏感的生理指標。
以主成分貢獻率的大小,計算各主成分的權重。如表3所示,4個主成分的權重值不等,分別為0.598,0.178,0.136和0.088。利用隸屬函數計算公式將指標值進行轉化,結合權重計算得到適應性評價值D。根據D值評價3種高寒沼澤草甸植物對不同增溫幅度的適應性,依次為:自然條件下,矮嵩草>青藏苔草>藏嵩草;低幅度增溫下,青藏苔草>矮嵩草>藏嵩草;高幅度增溫下,青藏苔草>矮嵩草>藏嵩草。

表3 植物的主成分得分(Zi)、權重、隸屬函數值、D值及適應性排序
3 討論
3.1 增溫對高寒沼澤草甸植物光合色素的影響
葉片中的葉綠素是植物進行光合作用的物質基礎,其含量能表征植物光合能力的強弱[35]。本研究中,3種植物葉綠素對增溫的響應存在顯著的互作效應,低幅度增溫下3種植物葉綠素含量均顯著增加,這可能是因為適度增溫使得葉片細胞內葉綠素合成相關酶活性升高,促進了3種植物葉綠素合成和積累,這與陳翔等[8]和王瓊等[36]研究結論一致。而高幅度增溫下矮嵩草和青藏苔草葉綠素含量的降低,與張麗霞等[37]研究類似,分析其原因一方面可能與較高的溫度造成溫室內水熱狀況改變有關,不適于葉綠素的形成,使其分解大于合成,另一方面高溫引起植物體內活性氧產生量的上升導致的氧化破壞,也可能使得葉綠素降解速度加快[38]。
類胡蘿卜素既可參與植物的光合作用,還可吸收細胞內剩余的能量,淬滅活性氧,從而降低膜脂過氧化程度[39-40]。袁嫚嫚等[41]研究表明,溫度升高顯著增加了水稻(Oryzasativa)生育后期的類胡蘿卜素含量。而本實驗結果表明,增溫處理下3種植物類胡蘿卜素與對照相比均顯著下降,與王義婧等[42]研究結果類似,分析類胡蘿卜素含量下降的原因可能是因為溫度在促進光合作用的同時,作為光合和呼吸作用副產物的活性氧也增加,類胡蘿卜素需參與活性氧猝滅,防止膜被氧化。
3.2 增溫對高寒沼澤草甸植物滲透調節物質的影響
植物會通過調節滲透物質來避免或減輕環境因子變化對其可能造成的傷害[43],在增溫處理下,植物細胞內滲透調節物質代謝的水平和途徑發生一系列適應性和抵抗性變化[44]。脯氨酸被認為是植物體內理想的有機溶質滲透調節物,多數植物受到脅迫后均能大量積累,適應環境變化所造成的影響[45]。董瑜等[46]研究結果表明,不同濕地植物葉片脯氨酸含量對氣溫升高的響應存在差異性。本研究中,相比對照,3種植物脯氨酸含量在高幅度增溫處理下大幅度增長,表明脯氨酸對高幅度增溫做出積極響應,以迅速積累的方式調節增溫對細胞滲透壓的影響,進而減少細胞受到的傷害,與楊春勐等[47]模擬增溫對滇楊(Populusyunnanensis)和川楊(Populusszechuanica)葉片脯氨酸的研究結果一致。
受到脅迫植物體內的可溶性糖含量也會積累,維持細胞正常滲透壓,且可溶性糖含量越高,緩解逆境所帶來傷害的能力越強。張秋芳等[48]研究表明,杉木(Cunninghamialanceolata)在外界環境影響下產生自我保護,可在短時間內通過調控可溶性糖等維持細胞正常功能。本研究中,低幅度增溫下矮嵩草和青藏苔草可溶性糖含量相比對照顯著增加,適當增溫可促進植物光合產物可溶性糖增加;而高幅度增溫下相比對照顯著下降,其原因可能在于增溫導致矮嵩草和青藏苔草光合作用失調,正常生理代謝和維持細胞滲透勢均需要大量可溶性糖[40]。藏嵩草相比矮嵩草和青藏苔草對增溫更為敏感,在低幅度增溫時已通過可溶性糖累積調節葉片細胞滲透勢,防止蒸騰失水,維持水分代謝平衡,進而提高自身對環境的適應力,與張玥等[49]研究結果一致。
與脯氨酸和可溶性糖一樣,可溶性蛋白也是一種重要的滲透調節物質,具有較強的親水能力,能夠顯著增加細胞親水能力,參與植物的滲透調節過程[45,50]。本研究中,3種植物可溶性蛋白在低幅度增溫下相比對照無顯著變化,且在高幅度增溫下顯著降低,這與曾貞等[51]得出增溫導致桑樹(Morusalba)葉片可溶性蛋白含量顯著增加的結論不一致。原因可能是,低幅度增溫使得植物光合速率增加,植物體需要更多的光合酶參與光合作用,從而使得可溶性蛋白含量降低[52],但光合產物可溶性糖含量增加,為合成蛋白質提供足夠碳源,保證蛋白質供需平衡。而高幅度增溫一方面引起的葉溫增幅較大,使蛋白水解酶活性增加,體內碳水化合物的供應受阻造成可溶性糖和可溶性蛋白含量減少[53];另一方面,增溫導致植物體細胞中活性氧自由基的積累,加速了蛋白質的分解。
3.3 增溫對高寒沼澤草甸植物丙二醛及抗氧化物酶活性的影響
生長在青藏高原高寒環境中的植物,抗氧化能力的高低是決定植物能否在高原復雜環境生存的重要因素[54]。丙二醛是細胞膜脂過氧化產物之一,其含量越多表明葉片受毒害作用越強,可間接反映植物組織的抗氧化能力[48]。本研究中,增溫處理下3種植物丙二醛含量整體維持于正常水平,并未出現大幅度增加,這與Martinez等[53]和吳久赟等[55]研究結果類似。分析其原因可能是低幅度增溫下植物光合速率較高會導致細胞中活性氧自由基積累增多,膜脂過氧化加劇使丙二醛增高,并激活了抗氧化系統,但3種植物葉片中積累的氧自由基并沒有超過細胞的控制力引發氧自由基代謝失衡,抗氧化酶協同作用清除過剩氧化性物質、抑制丙二醛生成或維持正常水平以減輕膜結構的損傷程度[56]。高幅度增溫下藏嵩草抗氧化酶盡管受到抑制,但并未發生氧自由基代謝失衡,矮嵩草和青藏苔草則受溫度影響,相比低幅度增溫產生大量氧自由基積累,同時升溫造成保護酶類的可溶性糖減少,進一步造成了丙二醛的增加。
超氧化物歧化酶作為防御的第一條線,它能特異性地將超氧陰離子(O2-)歧化為H2O2和O2,再由過氧化物酶,過氧化氫酶等將H2O2進一步催化分解成H2O和O2[57]。而本研究中,3種植物過氧化物酶和超氧化物歧化酶活性變化不一,且并未與活性氧含量表現一致,這與查倩等[58]研究結果一致。分析其原因可能是增溫破壞了抗氧化酶系統的平衡,使得細胞膜脂類成分發生過氧化作用,植物體內積累丙二醛,但溫度對酶類的影響又會促進抗氧化酶的生成,一定程度清除體內的活性氧,導致丙二醛含量有所下降[19],進一步印證了植物具有自我調節能力。3種植物抗氧化系統的差異可能是因為研究區域植物自身活性氧的產生與清除機制的差異,但3種植物均表現出在增溫處理下抗氧化酶需要一定程度的增強以抑制體內的活性氧的毒害。
3.4 高寒沼澤草甸3種植物增溫適應性評價
環境變化時,植物體內會發生一系列的生理生化反應,涉及滲透脅迫、活性氧積累等,是一個復雜的反應過程,其適應能力的大小是受多因子影響的表現[21-22]。本研究運用主成分分析法及隸屬函數法評價了3種植物對增溫的適應性,分析得出,3種高寒沼澤草甸植物對增溫的適應性,青藏苔草增溫適應性隨著溫度增加,優勢顯現,排序優于矮嵩草。3種植物對增溫的適應性排序發生變化,主要是因為不同物種對溫度的敏感性不同,因此增溫后表現出不同于自然條件下的適應模式。一方面,矮嵩草屬于耐寒旱中生植物,具有耐寒、耐旱等特點,能夠適應增溫導致的土壤含水量的減少;藏嵩草和青藏苔草均對水分變化極為敏感,但已有研究表明,青藏苔草既存在一般植物常有的適應生境升溫的機制,又因其生理整合現象,即營養物質和水分可以通過地下根莖進行分株間的運輸和轉移,實現整個無性系的資源共享,從而提高青藏苔草對高溫環境的適應性[59-61];另一方面,適度增溫有利于莎草科植物的生長發育的同時,促使植物發育速率加快,進而導致部分高寒植物的成熟過程提早,縮短實際生長期,生物量的減少,喪失優勢地位。李英年等[62]通多對比20年間植被結構數據,認為在氣候變暖的情況下,高寒濕地植物群落組成會發生變異,原生適應寒冷、潮濕生境的藏嵩草為主的草甸植被類型可能會被寒冷濕中生為主的典型草甸類型替代,部分結論與本次評價一致。此外,楊凱等[63]從形態學與化學計量學角度出發,分析得出增溫促進凍土融化提供一定營養供給,促進藏嵩草沼澤化草甸生物量的積累,提高其資源掠奪能力,加強自身生態地位,可見僅從植物生理層面評價植物對溫度的適應性,對于全面準確的預測增溫背景下沼澤草甸植物群落結構和功能的演變方向仍有所欠缺,后續需結合植物形態特征與生物量生產等方面內容進一步探究高寒沼澤生態系統對氣候變暖的響應機制。
4 結論
綜上所述,短期模擬增溫對3種高寒沼澤草甸植物生理方面造成了影響,3種植物在一定程度上通過自身調節來抵御增溫環境所帶來的影響,具體表現為低幅度增溫促進植物光合作用,通過滲透調節物質和抗氧化系統協同作用維持植物正常生長;高幅度增溫下,植物光合作用受到抑制,通過滲透調節物質增多和抗氧化酶活性增加,使得活性氧的破壞作用減輕,維持植物正常生理活動。同時,不同增溫幅度下,3種植物對增溫的適應性相對自然條件發生改變,適應性從強到弱依次為青藏苔草>矮嵩草>藏嵩草。3種植物生理響應及對其增溫適應性評價表明,高寒沼澤草甸不同增溫幅度下植物能通過自身生理特性的改變,對增溫環境做出有利于自身生長的反饋調節,增溫適應能力最強的物種出現分異,物種競爭能力可能會因此發生改變。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
