千穗谷TCP基因家族生物信息學分析
2022-04-25 13:14:20喻望晨劉寧芳胡龍興
草地學報 2022年4期
周 濤, 彭 輝, 喻望晨, 劉寧芳, 胡龍興, 徐 倩
(湖南農業大學農學院草業科學系, 湖南 長沙 410128)
籽粒莧是莧科莧屬一年生草本粒用植物的統稱,包括千穗谷(Amaranthushypochondriacus)、老鴉谷(A.cruentus)、尾穗莧(A.caudatus)、綠穗莧(A.hybridus)等。千穗谷為籽粒莧的一種,起源于美洲,目前主要分布于溫帶和熱帶地區,在平原和高海拔地區均有分布。千穗谷具有根系發達、吸肥力強、生長快、產量高、營養豐富等特點,是一種糧飼兼用型作物,其莖葉可作蔬菜,種子可食用或釀酒,也可栽培觀賞[1-3]。千穗谷耐酸、耐鹽堿,具有較強的抗旱性和耐受重金屬的能力,可作為土壤修復和生態恢復類型植被[4-6]。
TCP(Teosinte branched1/cycloidea/proliferating cell factor)是一類植物特有的轉錄因子家族,參與植物多種器官的生長發育與形態建成。目前已在擬南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、蒺藜苜蓿(Medicagotruncatula)、番茄(Solanumlycopersicum)、胡楊(Populuseuphratica)和白菜(Brassicarapa)中分別鑒定到24,30,21,30,36和39個TCP成員[7-11]。TCP的命名來自于三個蛋白的首字母縮寫——玉米(Zeamays)中參與頂端優勢調節的TEOSINTE BRANCHED1(TB1)、金魚草(Antirrhinummajus)中控制花不對稱性的CYCLOIDEA(CYC)以及水稻中參與分生組織表達的PROLIFERATING CELL FACTOR1(PCF1)和PCF2[12-14]。TCP蛋白的N端均包含一個高度保守的TCP結構域,由59個氨基酸構成,屬于非典型的bHLH結構[13]。根據該結構域氨基酸序列的差異,該家族可分為兩類:ClassⅠ(PCF)和ClassⅡ[15],其中,ClassⅡ可以進一步分為CIN和CYC/TB1兩個分支[16]。與ClassⅡ亞家族相比,ClassⅠ在bHLH結構的Basic區域有一個四氨基酸的缺失[17]。
TCP轉錄因子在植物的不同生長發育階段均有重要作用[18-19],包括葉片形態建成[20]、花器官發育[21]、根的生長和分蘗形成[22]等。白菜中的BrTCP7通過促進茉莉酸生物合成基因BrOPR3和葉綠素分解代謝基因BrRCCR的表達,從而促進葉片衰老[23]。毛竹(Phyllostachysedulis)中PeTCP10基因可通過MeJA途徑負調控側根的生長發育[24]。在擬南芥花中,AtTCP15通過調控IAA合成基因YUC1和YUC4影響IAA的合成[25]。近年來,越來越多的研究發現,TCP轉錄因子可以提高植物對多種生物脅迫和非生物脅迫的抗性。比如,香蕉(Musanana)TCP基因家族大部分基因響應低氮脅迫的誘導,并且調節香蕉缺氮時的逆境響應[26]。馬鈴薯(Solanumtuberosum)TCP轉錄因子StTCP13能提高大腸桿菌對高鹽脅迫的耐受性[27]。擬南芥CYC分支成員AtTCP18蛋白能和植原體效應因子SWP1互作,參與植物防御進程[28]。水稻中過表達OsTCP21基因可增強水稻對RRSV病毒的抵抗力[29]。
microRNAs(miRNAs)是一類內源性的小分子非編碼RNA[30],在基因表達過程中起負調控作用[31]。前人研究發現miR319的主要靶基因為TCP基因,且以ClassⅡ亞家族中的CIN分支基因成員為主,通過抑制它們的表達調控植物的生長發育以及對逆境的響應[32]。比如,水稻中miR319通過調控OsTCP21和OsGAmyb基因的表達使水稻分蘗減少、籽粒產量降低[33];梅(Prunusmume)中發現的Pm-miR319a可以通過調控靶基因PmTCP4的表達間接調控梅花雌蕊的發育[34]。
千穗谷最新基因組于2017年發布[35],但關于千穗谷TCP基因家族的系統研究未見報道。本研究擬采用生物信息學手段,對千穗谷TCP基因家族進行鑒定和特征分析,包括基因序列特征、結構域比較、蛋白理化性質預測、近源物種系統進化分析、染色體定位、啟動子順式作用元件預測、組織特異性表達等多個方面,為千穗谷TCP基因的功能研究和千穗谷分子育種提供重要的理論依據。
1 材料與方法
1.1 千穗谷TCP基因家族成員的鑒定
從Plant TFDB v5.0(http://planttfdb.cbi.pku.edu.cn)數據庫中下載千穗谷TCP蛋白序列,并在Phytozome v13(https://phytozome-next.jgi.doe.gov/)數據庫中進行blast搜索,得到基因序列全長。通過在線工具HMMER(https://www.ebi.ac.uk/Tools/hmmer/)分析所有蛋白的結構域,保留所有含有典型TCP結構域的基因。
1.2 千穗谷TCP 基因及蛋白結構分析
利用ExPASy網站(http://web.expasy.org/compute_pi/)預測千穗谷TCP蛋白的分子量(Molecular weight,MWs)、等電點(Isoelectric point,pIs)、平均親水性(Grand average of hydropathicity)等信息。利用GSDS2.0(http://gsds.gao-lab.org/index.php)在線網站繪制千穗谷TCP的外顯子和內含子結構。利用軟件MEME[36]預測蛋白保守Motif,使用TBtools[37]軟件繪制千穗谷TCP基因家族成員的基因結構圖。
1.3 多序列比對和系統發育樹構建
利用Clustal W[38]軟件對千穗谷、擬南芥、水稻的蛋白序列進行多重序列比對。千穗谷多重序列比對結果使用GeneDoc軟件進行展示,分析不同家族的特征結構域。系統發育樹采用Mega7[39]軟件構建,算法采用鄰近法,使用Evolview(https://www.evolgenius.info/evolview/)展示構建的系統發育樹。擬南芥以及水稻TCP基因家族數據下載于Plant TFDB v5.0(http://planttfdb.cbi.pku.edu.cn)。
1.4 基因染色體定位和蛋白亞細胞定位
通過BioEdit軟件對千穗谷每一個TCP家族成員進行本地BLAST搜索比對,以確定其在染色體中的定位,通過MG2C v2.1(http://mg2c.iask.in/mg2c_v2.1/)在線網站對TCP家族成員染色體定位結果進行圖形化處理。千穗谷全基因組數據下載于Phytozome數據庫(Amaranthushypochondriacusv2.1,DOE-JGI,http://phytozome.jgi.doe.gov/)。通過在線分析工具WoLF PSORT(https://wolfpsort.hgc.jp/)對千穗谷TCP蛋白的亞細胞定位進行預測。
1.5 千穗谷TCP基因啟動子順式作用元件預測
利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在線網站對千穗谷TCP基因的啟動子進行順式作用元件預測,使用TBtools軟件展示預測結果[37]。
1.6 千穗谷TCP基因組織特異性表達分析
從Phytozome數據庫獲取千穗谷TCP基因在不同發育時期不同組織器官的表達情況[40],通過Heml軟件對不同基因的表達量進行分層聚類分析并繪制熱圖。
1.7 調控千穗谷TCP基因的miRNA預測
利用psRNATarget(https://www.zhaolab.org/psRNATarget/home)在線網站對可能調控千穗谷TCP基因編碼區的miRNA進行預測。由于目前并沒有已報道的千穗谷miRNA,因此選擇雙子葉模式植物擬南芥miRNA數據。
2 結果與分析
2.1 千穗谷TCP基因家族的鑒定及特性分析
從Plant TFDB v5.0數據庫共檢索得到16個千穗谷TCP基因,分別命名為AhTCP1~AhTCP16(表1)。AhTCP基因長度介于270~1 410 bp之間,氨基酸序列長度在90~470之間,最長為AhTCP12,最短為AhTCP10。蛋白分子量介于9 790.9~50 986.9kD之間;理論等電點介于6.26~10之間,且僅有AhTCP2,AhTCP4,AhTCP8,AhTCP9,AhTCP11的理論等電點小于7,其他蛋白等電點均在堿性范圍內;平均親水性指標均為負值,表明AhTCP成員均是親水性蛋白。亞細胞定位預測的結果顯示,所有千穗谷TCP蛋白在細胞核中定位的概率最高,這與其轉錄因子的功能相符合。
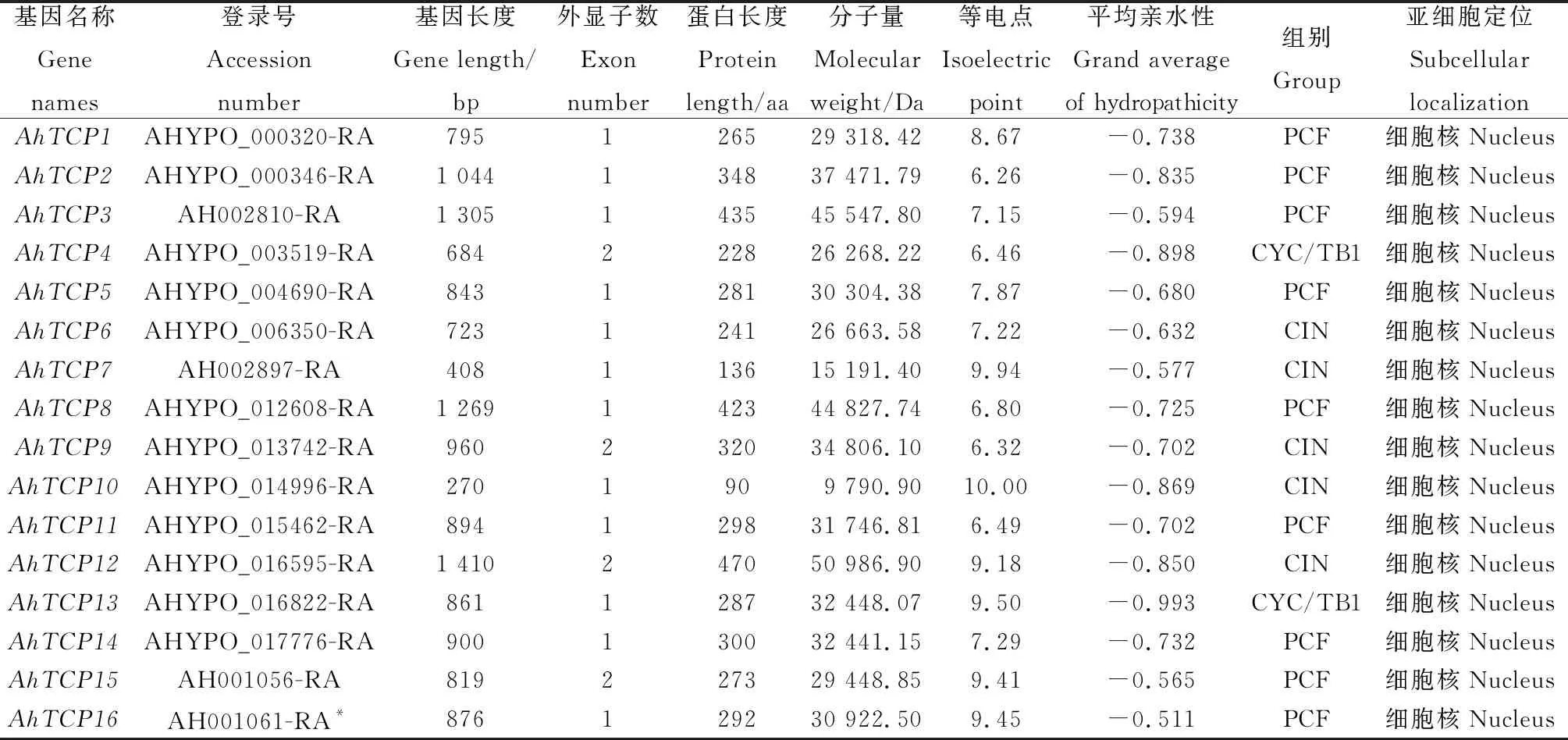
表1 千穗谷TCP 家族成員基本信息
2.2 千穗谷TCP蛋白多重序列比對與系統進化樹構建
對千穗谷TCP蛋白序列進行多重序列比對(圖1),發現16個成員的TCP結構域呈現不同的特點。典型的TCP結構域屬于螺旋-環-螺旋(bHLH)結構,其中AhTCP10在helixII區域有部分缺失,與擬南芥的AtTCP6相似。千穗谷TCP家族中有9個成員在Basic區域缺失了4個氨基酸,與其他物種TCP家族中的PCF亞家族類似,根據這一特征可以將千穗谷16個TCP基因分為兩個亞家族:ClassⅠ和ClassⅡ。根據Helix和Loop區域的序列差異(圖2),可以進一步將ClassⅡ分為CYC/TB1和CIN兩個亞類。在千穗谷中,CYC/TB1亞類特有的保守氨基酸包括Basic區域和HelixI區域之間的亮氨酸,HelixI區域的異亮氨酸和精氨酸,以及Loop區域的苯丙氨酸、賴氨酸和丙氨酸等;而CIN亞類中特有的保守氨基酸包括Basic區域的賴氨酸,HelixⅠ區域的蘇氨酸、異亮氨酸、谷氨酰胺和精氨酸,以及Loop區域的脯氨酸等。不同的亞類在HelixII區域的序列差異較大,多態性較高。
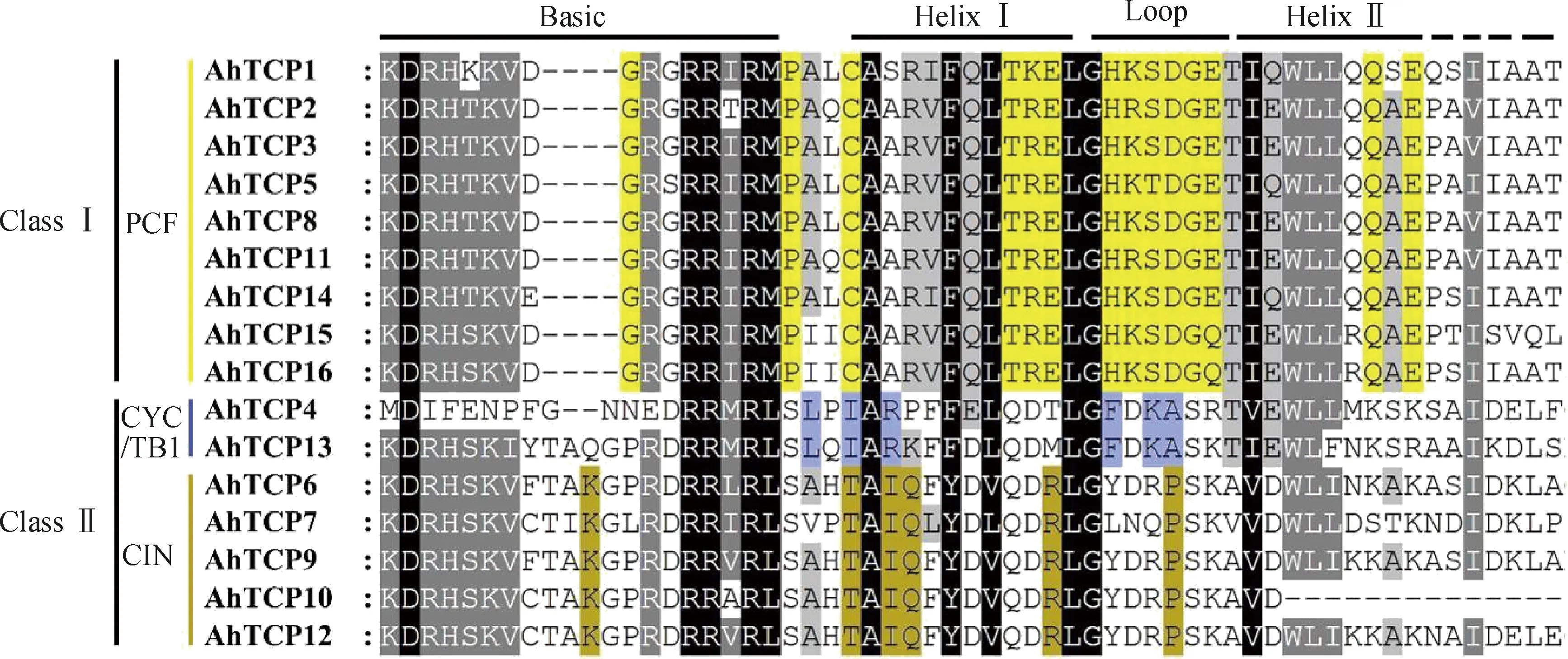
圖1 千穗谷TCP蛋白保守結構域序列對比
對千穗谷、擬南芥、水稻三個物種的TCP轉錄因子構建系統發育樹(圖2),發現TCP家族成員可分為三大類。根據序列特征,將三大類分別命名為PCF,CYC/TB1和CIN,如圖2所示。其中,PCF亞類包含了16個擬南芥基因、10個水稻基因和9個千穗谷基因;CYC/TB1亞類包含了5個擬南芥基因、3個水稻基因和2個千穗谷基因;CIN亞類包含了12個擬南芥基因、10個水稻基因和5個千穗谷基因。千穗谷的TCP家族中一半以上成員為PCF亞類,而擬南芥和水稻中ClassⅠ和ClassⅡ兩個亞類的成員數目幾乎相同。
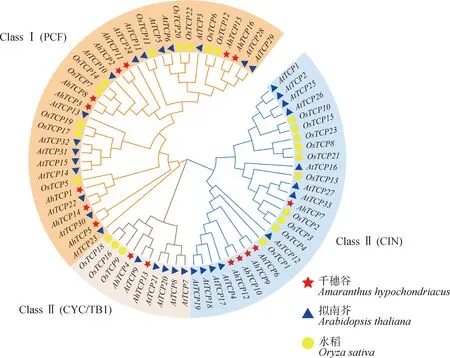
圖2 千穗谷、擬南芥和水稻的系統發育樹
2.3 千穗谷TCP基因結構和保守基序分析
千穗谷TCP家族成員的基因結構比較簡單,大部分AhTCP基因不含內含子,只有4個成員(AhTCP4,AhTCP9,AhTCP12,AhTCP15)含有1個內含子,占總數的25%。不同基因的內含子序列長度差異很大,AhTCP15的內含子長度僅為80 bp,而AhTCP9內含子序列長達791 bp。結合系統進化樹的結果可以看出,千穗谷TCP基因的內含子數目在不同亞家族之間具有一定的特異性:ClassⅠ亞家族中,除AhTCP15外的其他成員均不含內含子,而ClassⅡ亞家族中將近一半的成員含有1個內含子和2個外顯子(圖3)。

圖3 千穗谷TCP家族成員基因結構分析圖
千穗谷TCP蛋白的保守基序分析顯示,在進化樹中同一分支的AhTCP蛋白通常具有相似的基序組成。除了TCP結構域所在的motif1外,TCP家族不同成員之間還存在多個保守的DNA區段。比如,除AhTCP7外,Class Ⅱ的CIN亞類中都存在Motif3,Class Ⅰ的PCF分支中的AhTCP蛋白都具有Motif2和Motif5;有的Motif只在個別成員中存在,比如Motif6和Motif10僅存在于AhTCP3和AhTCP8中。
2.4 染色體定位分析
如圖4所示,千穗谷的16個TCP基因僅分布在Chr1,Chr2,Chr3,Chr4,Chr6和Chr11等6條染色體上,其他10條染色體無TCP基因分布。其中,Chr2染色體上共分布有5個TCP基因,且包含三個不同亞類;而Chr4和Chr6染色體上僅分布有1個TCP基因,均位于染色體端粒的位置。部分TCP基因在染色體上的分布呈現出成簇的現象,比如Chr1染色體上的AhTCP15,AhTCP16,相距僅0.17Mb;AhTCP1和AhTCP2兩個基因,均分布在Chr2染色體端粒部分34.27~34.54 Mb的區域內。
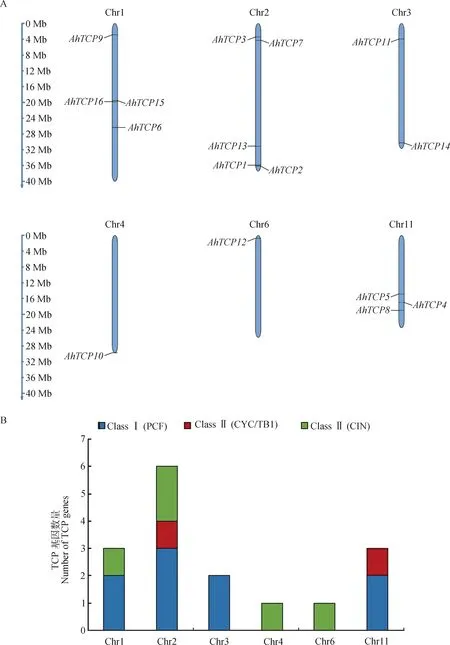
圖4 千穗谷TCP家族成員染色體定位信息
2.5 順式作用元件預測
對千穗谷TCP基因上游1 000 bp序列進行順式作用元件預測,發現千穗谷TCP基因的啟動子區域包含多個與植物生長發育和逆境響應相關的調控元件(圖6)。比如,G-Box,Box 4,GT1-motif,MRE等光響應元件,在所有AhTCPs啟動子區域均有發現;而響應無氧環境的ARE也存在于大部分的AhTCPs中。此外,部分AhTCP基因上游區域含有響應低溫的元件LTR,比如AhTCP7,AhTCP13和AhTCP14等;部分基因包含響應干旱脅迫的順式作用元件MBS,比如AhTCP5,AhTCP6和AhTCP10等。很多AhTCPs啟動子含有響應激素的順式作用元件,比如AhTCP5,AhTCP7,AhTCP15,AhTCP16中包含響應生長素的AuxRR-core或TGA-element元件,AhTCP4,AhTCP5,AhTCP6,AhTCP7中包含響應赤霉素的P-box或GARE-motif等元件。部分順式作用元件僅在少數AhTCP基因中存在,例如響應晝夜節律的Circadian僅在AhTCP5中存在,參與胚乳特異性表達的GCN4-motif僅在AhTCP10和AhTCP15中存在。
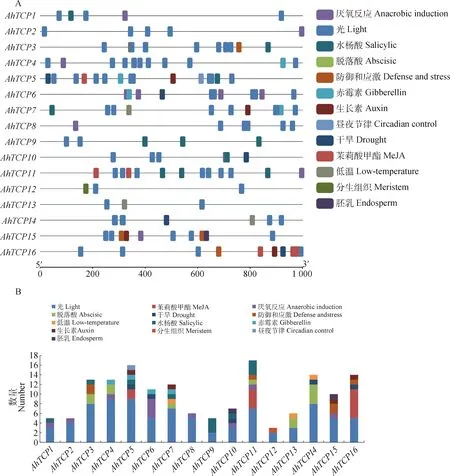
圖5 千穗谷TCP基因啟動子順式作用元件預測
2.6 組織特異性表達
基因的組織特異性表達通常與其功能密切相關。根據Phytozome數據庫中的信息可知,16個千穗谷TCP基因分別在根、莖、子葉、葉、花、未成熟種子、成熟種子和混合組織中檢測到不同水平的表達量(圖6)。不同亞家族的千穗谷TCP基因表達模式相似,比如PCF亞家族中的AhTCP2,AhTCP3,AhTCP5,AhTCP8,AhTCP14,AhTCP15,AhTCP16的表達模式被聚為一類;CIN亞家族中的AhTCP10和AhTCP12的表達模式被聚為一類且在大部分組織中未表達。其中,AhTCP4僅在花中表達;AhTCP13僅在根和莖中表達。AhTCPs在不同組織和器官中的表達模式不同,暗示不同成員在千穗谷的生長發育不同階段具有不同的功能。
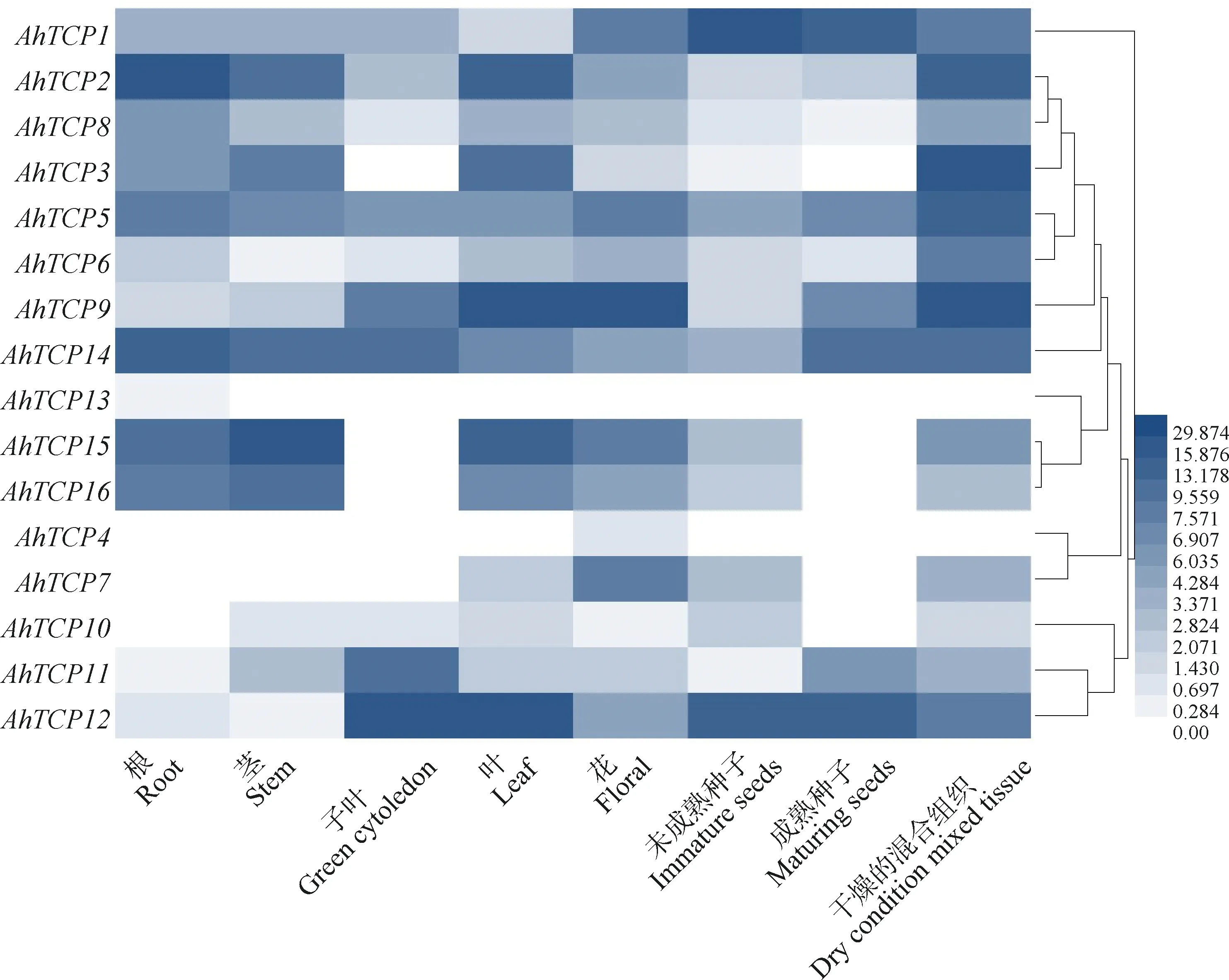
圖6 不同組織千穗谷TCP基因表達量分析
2.7 千穗谷TCP基因的miRNA預測
以AhTCP基因的編碼區為靶序列預測能夠與其互作的miRNA,發現多達47個類型的miRNA與AhTCP基因存在結合位點。其中,AhTCP12編碼區存在的結合位點最多,高達16個,AhTCP2存在的結合位點最少,只有1個(圖7A)。大部分的miRNA只跟1個AhTCP基因互作,但部分miRNA可以同時靶向不同的AhTCPs。比如,miR159,miR5658和miR834在千穗谷TCP基因中存在4個靶基因,miR472,miR838和miR319存在3個靶基因,另有11個miRNA存在2個靶基因(圖7B)。
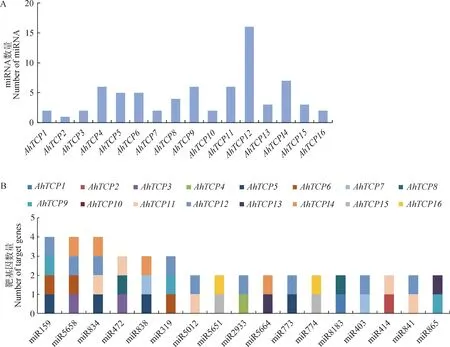
圖7 AhTCP基因的互作miRNA預測
3 討論
千穗谷TCP基因家族不同成員的基因長度、氨基酸大小、分子量和理論等電點差異較大,均編碼堿性氨基酸且定位于細胞核;根據氨基酸序列的差異,TCP基因家族可分為Class Ⅰ和Class Ⅱ兩大類,其中Class Ⅱ又分為CYC/TB1和CIN兩個分支[16]。千穗谷TCP基因家族也同樣包含三個亞類。序列比對發現,AhTCPs兩個螺旋區的保守氨基酸殘基中非極性氨基酸含量更多,例如丙氨酸(A)、亮氨酸(L)、異亮氨酸(I)、苯丙氨酸(F);環狀區域則包含許多親水性氨基酸殘基,例如賴氨酸(K)、精氨酸(R)、絲氨酸(S)、天冬氨酸(D)等,這與前人的報道相似[41]。氨基酸序列對比和保守基序分析均顯示不同的亞家族具有一定的保守性,例如在PCF,CYC/TB1,CIN亞類中均含有特定的保守的氨基酸(圖1)、Class Ⅰ亞家族中均含有Motif2和Motif5。基因結構分析顯示千穗谷TCP基因結構比較簡單,僅AhTCP4,AhTCP9,AhTCP12和AhTCP15含有一個內含子,大多數成員均沒有內含子。這與其他物種中的研究結果一致[41-42],表明TCP基因家族序列具有較強的保守性。染色體定位分析表明AhTCP基因僅定位在千穗谷的6條染色體上,基因在染色體上隨機分布且存在成簇的現象,這可能和基因的復制有關。
生物信息學分析可以為預測家族成員功能提供多方面依據。很多研究表明,直系同源基因通常具有相似的功能[43]。Danisman研究發現,AtTCP9通過改變茉莉酸代謝從而促進根的伸長[44],本研究顯示,千穗谷的AhTCP13基因和AtTCP9在系統進化樹中聚在同一個分支,且AhTCP13主要在根和莖中表達,因此AhTCP13很可能也具有與AtTCP9相似的功能和調控機制,在千穗谷的根莖發育中起重要作用。Sun等人發現擬南芥CIN亞類的AtTCP4具有激活次級細胞壁生物合成和細胞凋亡的功能,在花器官發育過程中起關鍵作用[21]。千穗谷的AhTCP9與AtTCP4聚在同一個分支,遺傳距離較近且組織特異性表達顯示其在花中表達量較高,推測AhTCP9基因很可能參與了千穗谷花的發育。前人研究表明,蓮藕(Nelumbonucifera)和豌豆(Pisumsativum)中CYC/TB1分支的TCP蛋白參與調控花的不對稱性[45-46],同樣屬于CYC/TB1分支的AhTCP4僅在花中表達,推測其很可能參與了千穗谷花的形態建成。
轉錄因子通過與下游靶基因啟動子上順式作用元件的相互作用,在植物發育過程和非生物脅迫反應中發揮重要作用[47]。本研究對千穗谷TCP啟動子順式作用元件進行預測,發現其啟動子區域含有大量與激素調控和脅迫反應有關的順式作用元件,暗示千穗谷的TCP基因可能參與了多種發育事件和脅迫反應。不同的千穗谷TCP基因啟動子區域的順式作用元件類型不同,顯示了家族基因間功能上的分化。比如,擬南芥中PCF分支的TCP14和TCP15通過促進幼嫩節間的細胞分裂來影響植株高度,還可調節葉片發育和花組織的細胞增殖[48],而千穗谷中與該基因同屬PCF分支的AhTCP15和AhTCP16在莖中表達水平較高,且啟動子中包含響應生長素的順式作用元件,推測AhTCP15和AhTCP16基因可能通過響應生長素進而調控千穗谷莖的生長(圖6)。此外,ClassⅠ亞家族的AhTCP3,AhTCP11,AhTCP15,AhTCP16在其啟動子區域包含多個與防御和應激相關的順式作用元件(TC-rich repeats),暗示其可能參與了植物的防御機制,這與擬南芥ClassⅠ亞家族中的AtTCP14,AtTCP15,AtTCP19等基因的功能相似[48]。
對AhTCP基因的編碼區進行miRNA作用位點預測,結果顯示有47個miRNA在AhTCP基因上存在結合位點,其中,miR319和miR5658的預測準確度最高。多個物種中的研究均表明,miR319主要靶向作用于TCP基因,且主要為ClassⅡ亞家族中CIN分支成員[32,49]。比如,水稻中的Osa-miR319bc通過調控OsPCF6和OsTCP21的表達調節水稻對冷的應激反應[50];雞爪槭(Acerpalmatum)中的miR319通過靶向ApTCP2影響植株開花、葉片形態和衰老[20]。本研究中,AhTCP6,AhTCP9和AhTCP12基因中均含有miR319的結合位點,且三者都屬于ClassⅡ亞家族中CIN分支成員,這與其他物種類似[51]。進一步分析顯示,AhTCP12與ath-miR319在互補配對區有4個堿基的差異,AhTCP6和AhTCP9與ath-miR319在互補配對區僅有2個堿基的差異(圖8)。

圖8 miR319靶向AhTCP基因的結合位點
4 結論
根據TCP結構域特點,千穗谷的16個TCP基因分為PCF,CYC/TB1和CIN三個分支,與其他物種的亞家族分類相似。染色體定位結果顯示,AhTCPs不均勻的分布在6條染色體上,部分基因呈簇狀分布。順式作用元件預測和組織特異性表達分析均顯示千穗谷TCP基因家族不同成員存在功能上的分化:不同AhTCPs的啟動子所包含的順式作用元件存在較大差異,暗示其響應不同的植物激素或者環境信號;16個AhTCPs基因在不同組織和器官中的表達模式不同,系統進化樹中同一分支的成員表達模式相似,不同分支的成員存在較大差異。miRNA預測結果顯示,miR319在千穗谷TCP家族中的靶基因也主要為CIN亞類成員。
