馬尾松次生林轉換為濕地松和杉木林對土壤細菌群落結構和多樣性的影響
2022-05-24 03:28:14夏開鄧鵬飛馬銳豪王斐溫正宇徐小牛
生態環境學報 2022年3期
關鍵詞:影響
夏開,鄧鵬飛,馬銳豪,王斐,溫正宇,徐小牛
安徽農業大學林學與園林學院,安徽 合肥 230036
建國以來,隨著我國對木材需求的不斷增加,大面積的原始森林逐漸被砍伐,經封山育林形成天然次生林(朱教君,2002)。由于部分天然次生林生產力低,被砍伐轉換成不同樹種的人工林。皖南山區于2009年前后,將馬尾松(Pinus massoniana)低效天然次生林(PM)改造為濕地松(P. elliottii)人工林(PE)和杉木(Cunninghamia lanceolata)人工林(CL)。林型轉換是驅動土壤環境條件發生改變的主要因素之一(Bruun et al.,2015),土壤微生物群落結構和多樣性可能會隨著土壤環境條件變化而發生改變(Chen et al.,2016),進而反映森林生態系統的土壤受損情況或恢復潛力(鄧嬌嬌等,2019),對土壤微生物群落結構和多樣性進行研究,可為森林土壤生態恢復和可持續經營提供理論基礎。
土壤微生物是陸地生態系統地球生物化學循環的重要驅動因素(Liu et al.,2017),其群落特性指標常被作為土壤生態系統生產力的關鍵生物指標(Bell et al.,2015;韓芳等,2021)。細菌是土壤中最豐富和最多樣的微生物類群(Dennis et al.,2013),廣泛參與土壤養分循環和能量轉換過程(Averill et al.,2016),同時可以通過分泌大量次生代謝產物,反饋影響土壤環境和植物生長(De et al.,2015),在維持土壤生態系統生產力和穩定性方面起著重要作用(Felske et al.,2000)。林型轉換是影響土壤細菌組成和多樣性的重要因素之一(Nakayama et al.,2019)。林型轉換改變了林間微氣候、凋落物的組成和分解、植物根系的活動,改變了土壤理化性質和土壤養分狀況,從而影響土壤細菌群落結構和多樣性。Meng et al.(2019)研究發現:亞熱帶地區原始森林向混交林轉換后,土壤細菌群落和多樣性變化不明顯,而向杉木和竹林轉換提高了土壤細菌多樣性,是土壤pH的變化導致了該地區林型轉化引起土壤細菌群落的變化。Sun et al.(2020)研究發現熱帶雨林天然林轉換為橡膠樹人工林后細菌多樣性顯著降低,土壤容重和濕度是影響該地區土壤細菌群落的關鍵因子。不同地區的林型轉換對土壤細菌群落影響結果不同,影響土壤細菌群落的關鍵環境因子也不同。皖南山區PM轉化成PE和CL后,對土壤細菌群落結構和多樣性產生何種影響以及哪些關鍵環境因子影響該地區的土壤細菌群落,目前尚不清楚。因此,研究該地區林型轉換對土壤細菌群落結構和多樣性的影響,有助于揭示其對森林土壤生態系統的影響機制,為該地區林型轉換后森林土壤生態系統的恢復和維持提供科學依據,對土壤生態系統的可持續發展具有重要意義。
基于16S rRNA的新一代高通量測序技術采用最新版QIIME 2分析流程,可以極大提升數據精確度與物種分辨率(Benjamin et al.,2016),更加真實地揭示環境中微生物群落結構組成(Mark et al.,2005)。本研究以安徽旌德縣國營蔡家橋林場立地條件相同的PM以及由PM轉換的PE和CL為對象,分析了林型轉換對土壤的理化性質、細菌群落結構和多樣性的影響以及關鍵環境因子對細菌群落結構的影響,以期為該地區森林土壤生態系統恢復和可持續經營提供科學依據。
1 研究地區與研究方法
1.1 研究地區概況
試驗地位于皖南山區國營蔡家橋林場,地處118°30′E,30°20′N,氣候屬亞熱帶濕潤季風氣候,光照充足,四季分明,山地氣候特征明顯。年平均氣溫為16 ℃,年平均降水量為1521 mm,無霜期230 d左右。基巖以板巖和花崗巖為主,土壤多為紅黃壤、山地黃壤。林區植被豐富,森林覆蓋率達85%,喬木主要有馬尾松、濕地松、杉木、鵝掌楸(Liriodendron chinense)、檫木(Sassafras tzumu)、苦櫧(Castanopsis sclerophylla)、甜櫧(C.eyrei)、木荷(Schima superba)等。林下植物主要有檵木(Lorpetalum chindensevar.rubrum)、杜鵑(Rhododendron simsii)、白茅(Imperata cylindrica)、蕨類(Pteridophyta)等。
1.2 樣地設計和土壤樣品采集
2020年8月,在蔡家橋林場選擇立地條件類型相同的馬尾松次生林(PM)以及馬尾松次生林伐后營造的濕地松林(PE)和杉木林(CL)3種林分各設置3個樣地,樣地面積均為20 m×20 m,共9塊樣地。調查林分基本情況詳見表1。
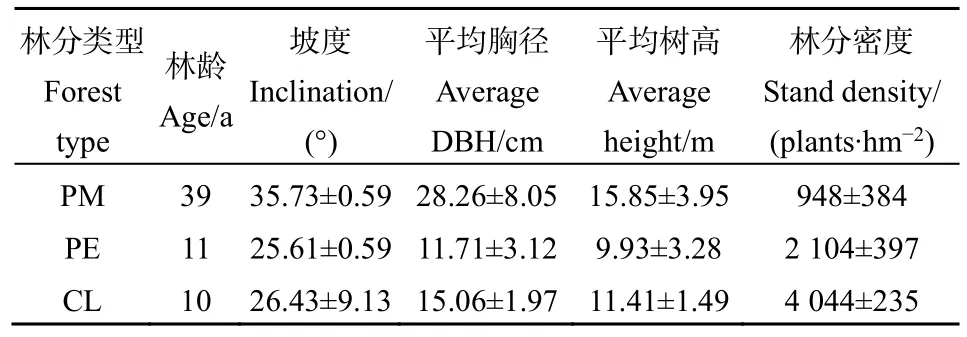
表1 調查林分樣地基本概況Table 1 Basic characteristics of the sampling stands
每個樣地內按機械隨機法設置3個小樣方,樣方面積5 m×5 m,土壤樣品采集之前,去除地表枯枝落葉,用土鉆在小樣方內按“S”形多點取樣,取樣深度為0—10 cm,將所取土壤充分混合成一個土樣,每個土樣分成2份,一份取500 g左右裝入無菌自封袋中,9個樣地共計27份土壤樣品,用于測定土壤理化性質;同時將另一份土壤,過2 mm篩以除去石頭、根系以及土壤動植物后,取10 g左右裝入無菌管中,并儲存于-80 ℃的干冰保溫盒中,共獲取 27份土壤微生物樣品,用于土壤微生物指標的測定。土壤微生物指標的測定委托杭州聯川生物科技有限公司進行16s rRNA高通量測序。
1.3 土壤理化指標的測定
土壤含水率(SWC,%)采用105 ℃烘干法測定;土壤pH按水土比例(2.5∶1)混合后用pH計測定;土壤電導率(EC,μS·cm-1)按水土比例(5∶1)混合后用電導率計測定;土壤有機碳(SOC)和全氮(TN)使用元素分析儀(EA 3000,Vector,Italy)測定;土壤全磷(TP)采用HNO3∶HClO4=3∶1法消煮后,利用連續流動注射分析儀(FIA Star 5000,FOSS,Denmark)測定。土壤速效氮(NH4+-N,NO3--N,mg·kg-1)使用 1 mol·L-1KCl溶液浸提,用流動注射分析儀進行測定。
1.4 土壤細菌16S rRNA測序方法
1.4.1 土壤細菌DNA提取與擴增測序
使用 E.Z.N.A.?Soil DNA Kit試劑盒對土壤細菌 DNA進行提取,并通過瓊脂糖凝膠電泳檢測DNA提取質量,同時采用紫外分光光度計對DNA進行定量。使用引物 341F(5′-CCTACGGGNGGC WGCAG-3′)和 805R(5′-GACTACHVGGGTATC TAATCC-3′)對16S rDNA可變區(V3—V4)進行PCR擴增。PCR產物采用Quant-iT PicoGreen dsDNA Assay Kit在Qbit熒光定量系統上對文庫進行定量,將合格的測序文庫梯度稀釋后,根據所需測序量按相應比例混合,經 NaOH變性為單鏈后使用NovaSeq測序儀進行2×250 bp的雙端測序。
1.4.2 生物信息數據處理
上機測序完成后,得到原始的下機數據,利用overlap將雙端數據進行拼接,并進行質控、嵌合體過濾,獲得高質量的cleandata。采用最新版QIIME 2分析流程,調用 DADA2(Divisive Amplicon Denoising Algorithm)(Benjamin et al.,2016)對序列進行去噪,即不再使用以97%相似度聚類成OTU(Operational Taxonomic Units)的分析方法,而是通過“去重復”(Dereplication,相當于以100%相似度聚類)等步驟,進而獲得單堿基精度的代表序列,大大提升了數據精確度與物種分辨率,然后使用ASVs(Amplicon Sequence Variants)的概念構建類OTU表,獲得最終的Feature特征表以及特征序列。本次測序共獲得23497個特征序列,特征序列所對應總Feature數為1021518個,3種林型共識別出39門,783屬細菌類群。采用Blast進行序列比對,每個代表性序列用 SILVA(Release 132,https://www.arb-silva.de/documentation/release-132/)以及 NT-16S數據庫對特征序列進行注釋,最后基于得到的 Feature特征表進行多樣性分析、物種分類注釋和菌群差異分析等。
1.5 數據處理
本研究分析平臺為QIIME 2平臺,數據計算及其分析作圖在QIIME 2自帶腳本R完成。運用Excel 2013對數據進行整理,計算所有參數的平均值和標準差(SD)。使用R(v 3.6.1)統計分析軟件對土壤理化性質和細菌α多樣性指標分別進行單因素方差分析(ANOVA),并對兩者進行 Pearson相關性分析,對土壤理化性質和細菌群落結構進行Mantel檢驗分析與Pearson相關性分析。
2 結果與分析
2.1 林型轉換對土壤理化性質的影響
PM轉換成PE和CL后,土壤理化性質發生顯著變化(表2)。與PM相比,PE土壤pH和NH4+-N含量顯著增加(P<0.05),而土壤EC和NO3--N含量顯著降低(P<0.05);CL土壤 SWC顯著增加(P<0.05);土壤 EC和 SOC含量均顯著降低(P<0.05)。
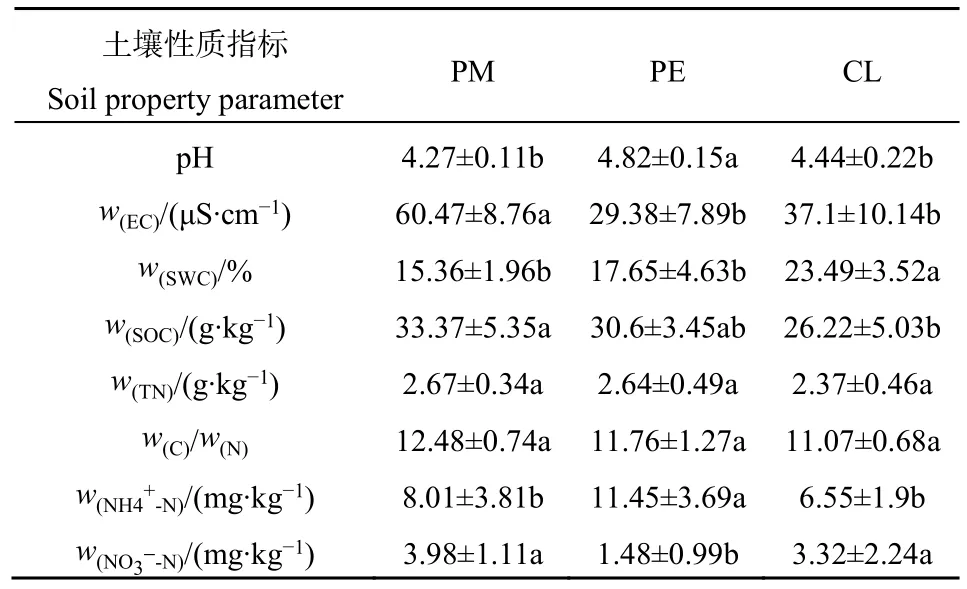
表2 試驗林分土壤理化性質Table 2 Soil basic characteristics of the experimental stands
2.2 林型轉換對土壤細菌群落多樣性的影響
2.2.1 林型轉換對土壤細菌α多樣性的影響
根據對土壤細菌α多樣性指數進行單因素方差分析(表3)可以看出:Chao1指數、Observed_species指數、Shannon指數、Simpson指數的變化趨勢均是PE>CL>PM。PM轉換成PE后,土壤細菌各α多樣性指數均顯著增加(P<0.05);而轉換成CL后,土壤細菌各α多樣性指數無顯著變化(P>0.05)。

表3 土壤細菌α多樣性指數Table 3 Soil bacterial α-diversity indices
對土壤細菌α多樣性指數與土壤理化性質進行Pearson相關分析發現:Chao1指數和Observed_species指數均與土壤 pH呈顯著正相關(P<0.05);Shannon指數、Simpson指數與土壤pH呈極顯著正相關(P<0.01),Simpson指數與EC呈顯著負相關(P<0.05)。
2.2.2 林型轉換對土壤細菌β多樣性的影響
對土壤細菌群落進行基于 weighted_unifrac的主坐標分析(Principal coordinates analysis,PCoA)和相似性分析(Analysis of similarities,Anosim),結果表明(圖 1):PCoA中兩個維度分別解釋了36.42%和 21.13%,共解釋了 57.55%(r=0.4084,P=0.001),代表PM、PE和CL的點可以分別聚在一起,且PE、CL與PM可以明顯區分開,3種林型組間差異明顯大于組內差異。
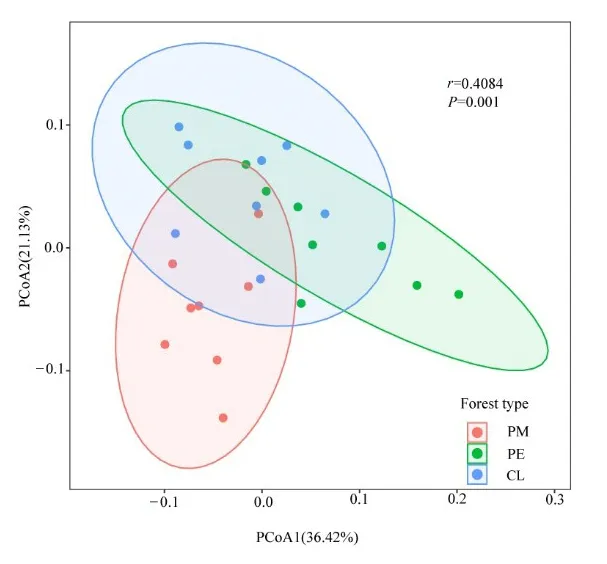
圖1 土壤細菌群落基于weighted UniFrac距離的主坐標分析Figure 1 Principal coordinate analysis of soil bacterial community based on weighted UniFrac distance
2.3 林型轉換對土壤細菌群落結構的影響
2.3.1 林型轉換對土壤細菌群落結構在門水平的影響
在門水平上,PM、PE、CL林分土壤共獲得39個細菌門。將平均相對豐度<1%歸為其他(Others),得到11個類群(圖2)。3種林型土壤細菌主要優勢菌群(平均相對豐度>10%)為酸桿菌門(Acidobacteria)(39.31%、36.70%、41.18%),其次是變形菌門(Proteobacteria)(31.77%、30.45%、27.11%)和放線菌門(Actinobacteria)(10.69%、10.73%、7.75%),總體相對平均豐度達到78.56%以上。其他7個優勢菌群分別為:綠彎菌門(Chloroflexi)、疣微菌門(Verrucomicrobia)、浮霉菌門(Planctomycetes)、芽單胞菌門(Gemmatimonadetes)、WPS-2,己科河菌門(Rokubacteria)未識別門(Unclassified)和其他(Others)。
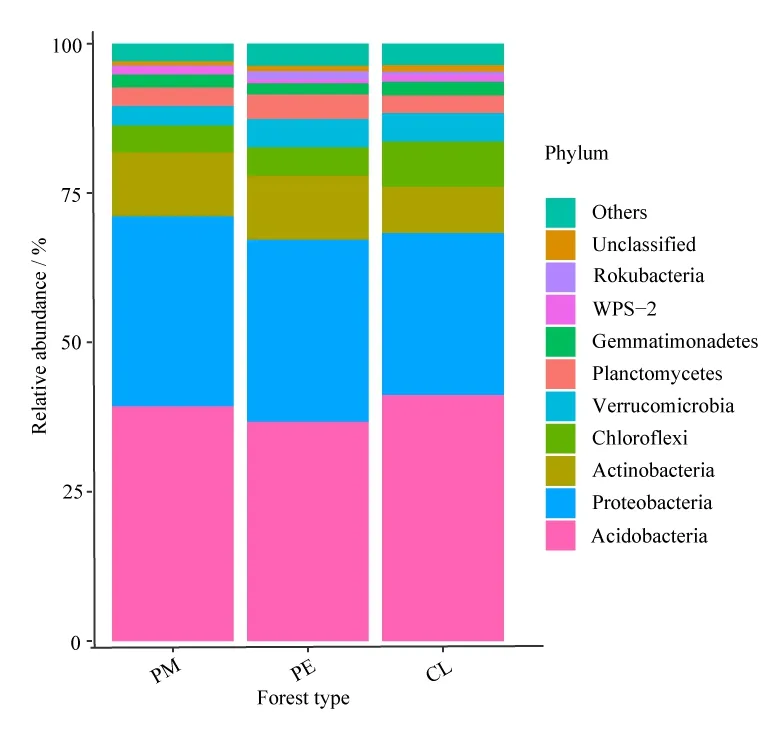
圖2 門水平上的細菌群落相對平均豐度堆疊柱狀圖Figure 2 Stack histogram of relative average abundance of bacterial community at phylum level
采用Wilcoxon方法對PE、CL分別和PM進行兩兩比較檢驗(表 4)發現:在三大優勢菌群中,酸桿菌門相對豐度在PM轉換為PE和CL后均未發生顯著變化(P>0.05),變形菌門和放線菌門相對豐度在PM轉換為CL后顯著降低(P<0.05)。其他主要菌群的平均相對豐度在 PM轉換為 PE和 CL后也發生了變化,PE與 PM 相比,己科河菌門和WPS-2均有極顯著差異(P<0.01);CL與PM相比,綠彎菌門和疣微菌門有極顯著差異(P<0.01),己科河菌門和未識別門有顯著差異(P<0.05)。
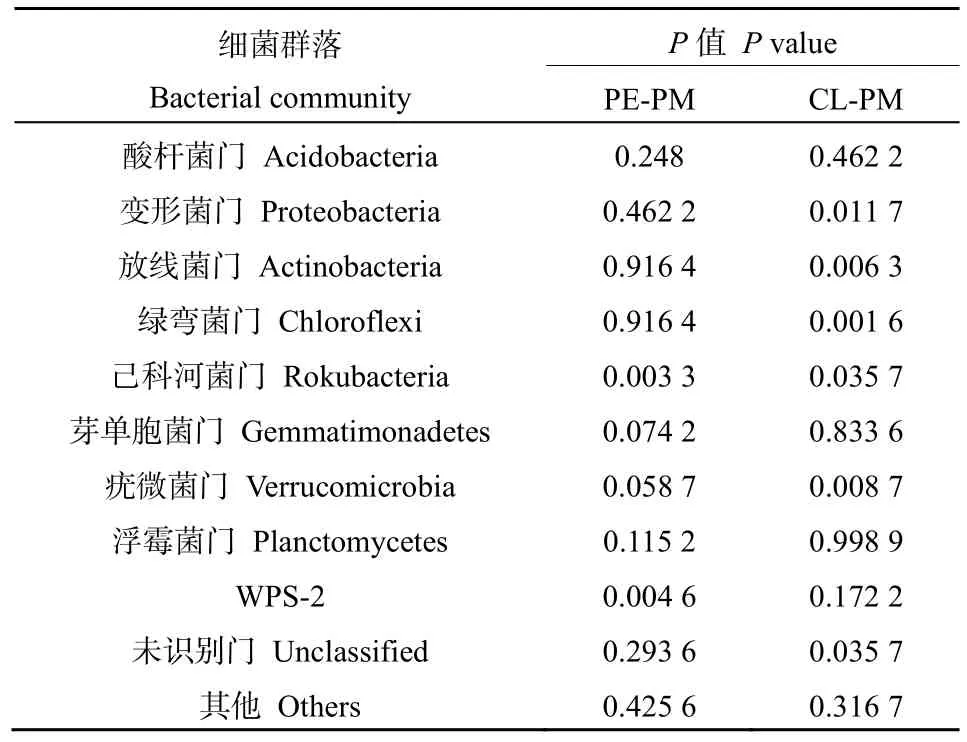
表4 PE、CL分別和PM土壤細菌門水平的Wilcoxon兩兩比較檢驗結果Table 4 Statistical results of Wilcoxon pair-to-pair comparison test at soil bacteriophyta levels among PE, CL and PM
2.3.2 林型轉換對土壤細菌群落結構在屬水平的影響
在屬水平上,3種林型共獲得783個類群。將PM、PE、CL中平均相對豐度<1%細菌類群歸為其他(Others),共有 20個類群(圖 3),占比超過37.21%,主要優勢菌屬(平均相對豐度>5%)有 3個,分別為酸桿菌門 Subgroup_2未定屬(Subgroup_2_unclassified)(18.45%、12.66%、14.11%)、酸桿菌目未定屬(Acidobacteriales_unclassified)(7.72%、6.69%、7.35%)、變形菌門Elsterales目未定屬(Elsterales_unclassified)(8.56%、3.80%、5.51%)。
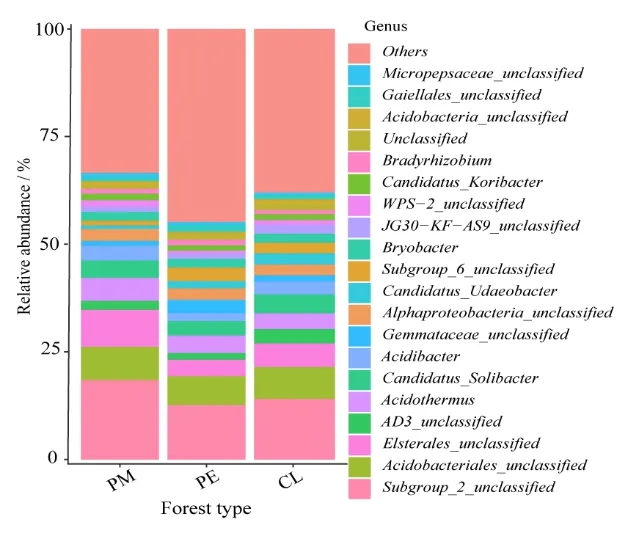
圖3 屬水平上的細菌群落相對平均豐度堆疊柱狀圖Figure 3 Stack histogram of relative average abundance of bacterial community at the genus level
采用Wilcoxon方法對PE、CL分別和PM進行兩兩比較檢驗發現:林型轉換顯著改變土壤細菌在屬水平的結構,優勢菌群(平均相對豐度>1%)的平均相對豐度在PM轉換為PE后有8個屬產生了顯著差異(表 5),分別為:酸桿菌門的 3個屬[Subgroup_6綱未定屬(Subgroup_6_unclassified),Candidatus_Solibacter屬(Candidatus_Solibacter),Subgroup_2目未定屬(Subgroup_2_unclassified)],變形菌門的 2個屬[Elsterales目未定屬(Elsterales_unclassified),黃色桿菌科未定屬(Xanthobacteraceae_unclassified)],放線菌門的Gaiellales科未定屬(Gaiellales_unclassified),浮霉菌門的出芽科未定屬(Gemmataceae_unclassified)以及疣微菌門的 Candidatus_Udaeobacter屬(Candidatus_Udaeobacter)。PM 轉換為 CL后有 8個屬產生了顯著差異(表5),分別為:酸桿菌門的Subgroup_6綱未定屬(Subgroup_6_unclassified),放線菌門的熱酸菌屬(Acidothermus),變形菌門里的 2個屬(Elsterales目未定屬(Elsterales_unclassified),α-變形桿菌綱未定屬(Alphaproteobacteria_unclassified),綠彎菌菌門的2個屬(JG30-KF-AS9科未定屬(JG30-KF-AS9_unclassified),AD3綱未定屬(AD3_unclassified)和疣微菌門的 2個屬[Candidatus_Udaeobacter屬(Candidatus_Udaeobacter),ADurb.Bin063-1 屬(ADurb.Bin063-1) ]。

表5 PE、CL和PM土壤細菌屬水平的Wilcoxon兩兩比較檢驗結果Table 5 Statistical results of Wilcoxon pair-to-pair comparison test at soil bacterial genera levels among PE, CL and PM
2.4 關鍵環境因子與細菌群落結構的相關性
對細菌群落結構和土壤理化性質進行 Mantel分析檢驗(表6)土壤pH對本研究地區土壤細菌群落結構影響最大(r=0.4992,P=0.001),同時 EC(r=0.3347,P=0.002)對細菌群落結構也有極顯著影響。
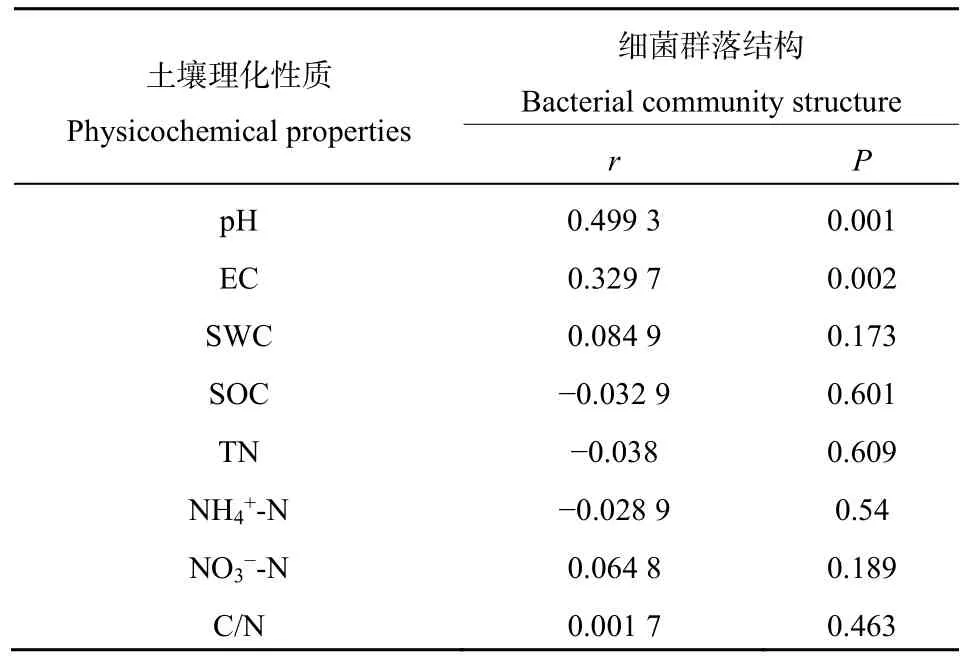
表6 Mantel檢驗細菌群落結構與土壤環境因子的相關性Table 6 Correlation between bacterial community structure and soil environmental factors tested by Mantel
進一步對門水平的優勢菌群與土壤理化性質進行 Pearson相關性分析(圖 4),酸桿菌門(P<0.05)、WPS-2(P<0.01)相對豐度與土壤 pH 呈顯著負相關,浮霉菌門(P<0.05)、己科河菌門(P<0.01)與土壤pH呈顯著正相關;己科河菌門、疣微菌門與土壤EC呈顯著負相關(P<0.05),WPS-2與土壤EC呈顯著正相關(P<0.01)。除了關鍵因子對菌群相對豐度影響外,還有其他環境因子也對部分菌群相對豐度產生影響。如:變形菌門(P<0.05)、放線菌門(P<0.05)的相對豐度與SWC呈顯著負相關,綠彎菌門與 SWC呈顯著正相關(P<0.01);酸桿菌門與TN呈顯著負相關,己科河菌門、浮霉菌門與TN呈顯著正相關(P<0.05)。酸桿菌門、WPS-2與NO3--N呈顯著正相關(P<0.05),浮霉菌門與NO3--N呈顯著負相關(P<0.05)。放線菌門與NH4+-N呈顯著正相關(P<0.01)。
對屬水平有顯著差異的優勢菌群與土壤理化性質進行 Pearson相關性分析(圖 4),酸桿菌門Subgroup_6綱未定屬、變形菌門的黃色桿菌科未定屬、放線菌門Gaiellales科未定屬、浮霉菌門出芽科未定屬相對豐度均與土壤 pH呈極顯著正相關(P<0.01),酸桿菌門 Subgroup_2綱未定屬和Candidatus_Solibacter屬(P<0.05)、變形菌門Elsterales目未定屬(P<0.01)與土壤pH呈顯著負相關。變形菌門Elsterales目未定屬與EC呈顯著正相關(P<0.05),酸桿菌門 Subgroup_6綱未定屬(P<0.01)、變形菌門黃色桿菌科未定屬(P<0.01)、放線菌門Gaiellales科未定屬(P<0.01)、疣微菌門ADurb.Bin063-1屬(P<0.05)、浮霉菌門出芽科未定屬(P<0.01)與EC呈顯著負相關。此外NO3--N也顯著影響了優勢菌群相對豐度的差異。變形菌門Elsterales目未定屬與NO3--N呈顯著正相關,酸桿菌門Subgroup_6綱未定屬(P<0.05)、變形菌門黃色桿菌科未定屬(P<0.01)、放線菌門Gaiellales科未定屬(P<0.01)、疣微菌門Candidatus_Udaeobacter屬(P<0.05)、浮霉菌門出芽科未定屬(P<0.05)與NO3--N呈顯著負相關。
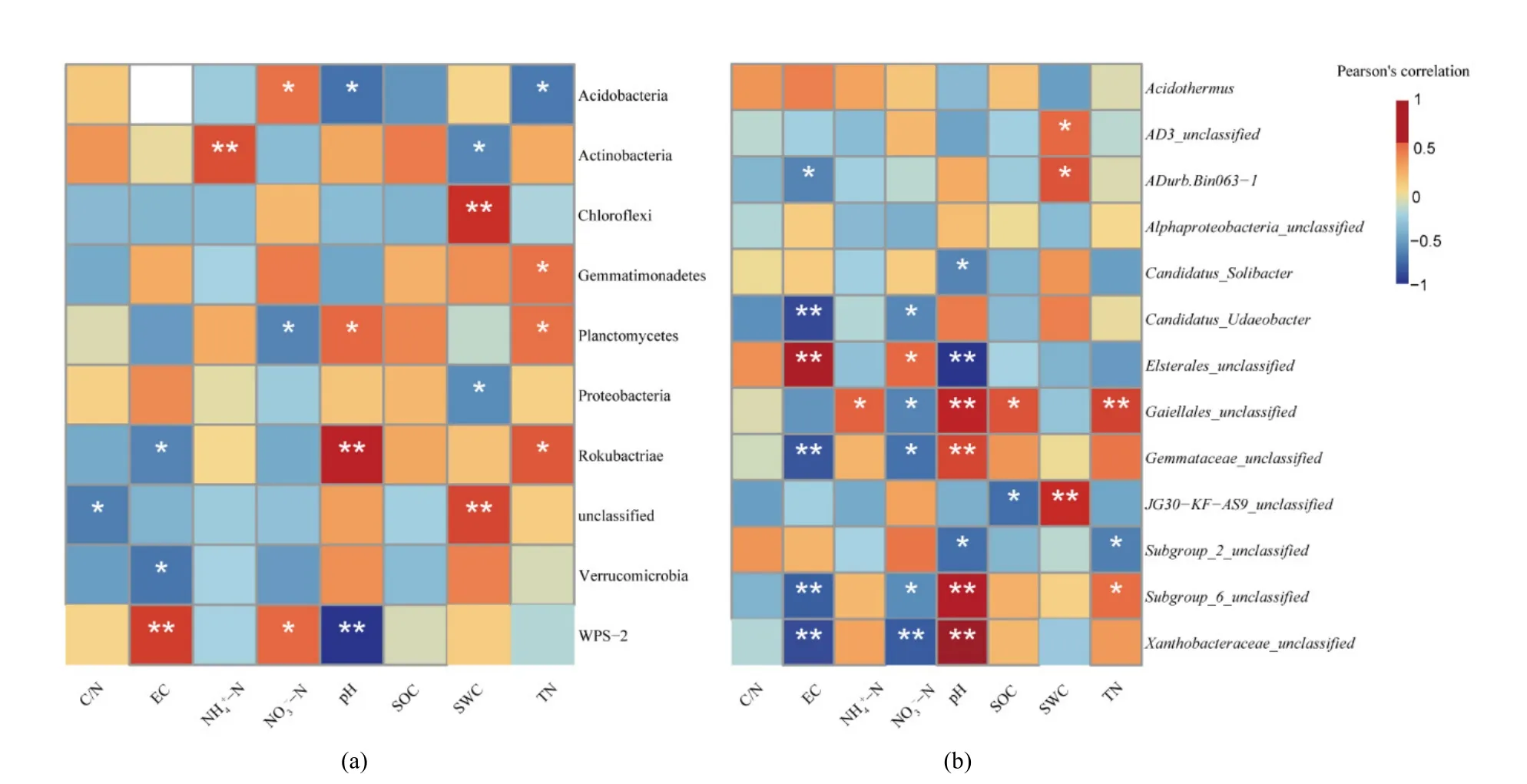
圖4 優勢門菌群相對豐度(a)、有差異的優勢屬菌群相對豐度(b)和土壤理化性質相關性熱圖Figure 4 Heat map of correlation between relative abundance of dominant phylum (a), relative abundance of dominant genera with differences (b) and soil physical and chemical properties
3 討論
3.1 林型轉換對細菌群落多樣性的影響
本研究中PM轉換成PE后,土壤細菌α多樣性顯著增加,而PM轉換成CL后,未發生顯著改變,同時β多樣性分析顯示,3種林型間的細菌群落差異顯著大于各林型內部差異,表明林型轉換顯著改變了土壤細菌群落的多樣性。PM轉換成PE和CL后,對土壤細菌群落多樣性影響不同,這與Meng et al.(2019)研究結果相一致。他們發現亞熱帶地區原始森林向混交林轉換,土壤細菌多樣性變化不大,向杉木和竹林轉換提高了土壤細菌多樣性。主要原因可能是不同類型的林型轉換形成了不同的林分植被結構,影響了林分凋落物組成、分解過程和根系活動,改變了土壤 pH等理化性質(Sun et al.,2020;梁國華等,2015;曾婷婷等,2016),從而影響土壤細菌多樣性。研究地區的土壤細菌Chao1指數、Observed_species指數、Shannon指數和 Simpson指數與土壤 pH均呈顯著正相關,與Hartman et al.(2008)研究的結果相一致,他們發現在pH小于6.5的酸性土壤中,微生物多樣性隨著土壤pH的降低而降低。凋落物是森林生態系統的重要組成部分,其分解可以產生多種化合物直接改變土壤pH(萬春紅等,2015),植物可通過根系活動向土壤中分泌不同的有機酸、糖類和酚類等次生代謝產物改變土壤pH理化性質,對微生物的生長和增殖產生影響(Zhao et al.,2012;Liu et al.,2020;吳林坤等,2014)。低土壤pH值的土壤中,較高的H+濃度會破壞細菌細胞膜的通透性和穩定性(Feng et al.,2014),抑制細菌對土壤養分的利用效率,進而影響土壤細菌多樣性(Shen et al.,2013;張坤等,2017)。PM轉換成PE后,土壤pH顯著增加,提升了土壤細菌對土壤養分的利用效率,從而導致土壤細菌多樣性顯著增加。
3.2 林型轉換對細菌群落結構的影響
林型轉化顯著改變了土壤細菌群落結構。PM轉換成PE后,己科河菌門、WPS-2和8個優勢屬相對豐度發生顯著變化;轉換成CL后,變形菌門、放線菌門、綠彎菌門、疣微菌門、己科河菌門和 8個優勢屬的相對豐度發生顯著變化。林型轉換改變了林間微氣候影響了土壤含水率;森林通過凋落物的歸還與分解實現土壤養分循環,其組成和分解速率在很大程度上影響土壤養分積累(鄧嬌嬌等,2019);不同林型的凋落物與根系分泌物組成不同,這些綜合因素導致了土壤理化性質和土壤養分含量的差異(Sun et al.,2020)。經Pearson相關性分析可知,研究區域優勢菌群的相對豐度與土壤pH、EC、SWC、SOC、TN、C/N、NO3--N、NH4+-N之間存在著一定的相關性。不同的研究也證實了土壤pH、EC、SWC和土壤養分指標是影響土壤微生物群落的重要因素(Zhang et al.,2014;Liu et al.,2020;Sheremet et al.,2020),這說明林型轉換是通過改變了土壤理化性質和土壤營養狀況,影響了土壤細菌的群落結構。
研究地區屬酸性土壤,不同研究表明酸桿菌門、變形菌門和放線菌門偏好土壤pH較低的土壤(Lauber et al.,2009;Sun et al.,2014)。3 種林型土壤優勢菌群均為酸桿菌門、變形菌門和放線菌門,與鄧嬌嬌等(2019)對遼東山區人工針葉林的土壤細菌群落研究結果一致。由于不同細菌的生活方式不同,可以被用作衡量土壤營養狀況的指標(Hartman et al.,2008)。酸桿菌門大多屬于貧營養型類群(Li et al.,2016),變形菌門和放線菌門屬于富營養型類群,主要參與有機質分解(Fazi et al.,2005)。PM轉換為PE后,三大優勢菌門的相對豐度未發生顯著變化,而轉換為CL后,變形菌門和放線菌門相對豐度顯著降低。有研究表明變形菌門的相對豐度與土壤碳含量呈正相關關系(Fierer et al.,2007),其豐度隨著有機質的增加而增加(Navarrete et al.,2008),放線菌門的相對豐度與NH4+-N含量呈顯著正相關(黃慶陽等,2021)。轉換后的CL中幾乎無林下植被,凋落物少,杉木凋落物的枝和葉常常連在一起,很難與土壤充分接觸,分解緩慢,導致土壤養分含量下降嚴重(何斌等,2002),CL的土壤SOC和NO3--N含量顯著下降,NH4+-N含量在3種林分中最低,從而導致富營養性菌的相對豐度顯著降低,表明轉換成CL后林分土壤營養狀況下降,不利于森林土壤系統的可持續發展。另外,WPS-2的相對豐度在貧瘠土壤中較高(Sheremet et al.,2020),綠彎菌門主要以CO2為碳源產生能量,能在 SOC含量較低的土壤中具有競爭能力(王鵬等,2017),轉換后的CL中WPS-2和綠彎菌門相對豐度均顯著增加,也證明了CL林分土壤營養狀況的下降。
3.3 關鍵環境因子對細菌群落的影響
PM轉換成PE后,土壤pH顯著增加,EC顯著降低,轉換成CL后,EC顯著降低。通過對土壤細菌群落結構和環境因子的Mantel分析檢驗得出,土壤pH是影響皖南山區3種林型土壤細菌群落的最重要驅動因子,土壤EC是影響該地區土壤細菌群落的另一個重要環境因子。趙鳳艷等(2019)研究發現土壤pH和EC是影響土壤微生物結構和多樣性的重要因子。全球尺度上,土壤pH被認為是影響土壤細菌群落結構和多樣性的關鍵因子(Bárcenas-Moreno et al.,2016;張紅霞等,2019;劉海洋等,2021)。土壤細菌能夠在一定pH范圍內生存,且存在最適pH,過高或過低的pH會破壞細菌細胞膜的通透性和穩定性(Feng et al.,2014),從而抑制細菌的生長和增殖(Zhou et al.,2020)。土壤環境中的pH變化會改變土壤營養物質分子的電離狀態,降低其被土壤細菌利用的有效性(潭洪治,1988)。土壤pH還會通過影響細菌群落之間以及與其他微生物之間的競爭與共生關系,從而影響細菌群落結構和多樣性(Shen et al.,2013)。本研究中優勢菌群有酸桿菌門等4個門與土壤pH顯著相關;在屬水平上,有酸桿菌門Subgroup_2綱未定屬等7個有差異的優勢菌群與土壤pH顯著相關;土壤細菌α多樣性指數與土壤pH均呈顯著正相關。土壤EC這一指標包含了反映土壤品質和物理性質的多項信息,如:土壤含水、鹽分、質地結構和有機質含量等(Rhoades et al.,1990)。本研究中,在門水平上,有疣微菌門等3個優勢菌群與EC顯著相關;在屬水平上,有變形菌門Elsterales目未定屬等6個有差異的優勢菌群與EC顯著相關;Simpson多樣性指數與EC呈顯著負相關。不同研究表明:SWC、SOC、TN、C/N、NO3--N、NH4+-N是影響不同地區土壤細菌群落的關鍵環境因子(Zhang et al.,2016;李明等,2020;于少鵬等,2020;姜雪薇等,2021),Pearson相關分析結果顯示這些土壤理化指標都與本研究地不同的細菌群落有著顯著相關性,但并不是影響該地區土壤細菌群落的關鍵環境因子。主要原因可能是這些土壤環境因子共同作用,綜合影響了該地區土壤細菌群落,而土壤EC是這些理化性質的綜合反映,成為影響該地區土壤細菌群落的重要環境因子。
4 結論
林型轉換是影響土壤細菌群落結構和多樣性的重要因素,土壤pH和EC是影響該地區土壤細菌群落結構和多樣性的關鍵環境因子。馬尾松次生林轉換成濕地松人工林后,土壤細菌多樣性顯著增加,細菌群落結構變化不明顯;而轉換成杉木林后土壤細菌多樣性無顯著變化,富營養型菌的相對豐度顯著降低。綜上所述,馬尾松次生林轉換為濕地松人工林比轉換為杉木人公林有利于土壤細菌群落發育和土壤營養狀況的維持,有利于森林土壤生態系統的可持續發展。另外,由于喬木根系較深,對深層土壤微生物活性也有一定的影響,如果做土壤剖面進行取樣,將會得到更為全面的結果。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
