BET蛋白抑制劑JQ?1對系統性紅斑狼瘡患者外周血B細胞分化的調控作用
2022-06-06 03:15:36李昊曾珊邱茜肖游君徐思琪王競男石茂華許韓師梁柳琴
實用醫學雜志 2022年8期
李昊 曾珊 邱茜 肖游君 徐思琪 王競男 石茂華 許韓師 梁柳琴
中山大學附屬第一醫院風濕免疫科(廣州 510080)
系統性紅斑狼瘡(SLE)是一種累及多系統、多器官的異質性自身免疫性疾病,以自身反應性B淋巴細胞多克隆增殖活化及產生大量致病性自身抗體為特點[1]。體內B 細胞的平衡被破壞,是SLE疾病活動的特點,包括幼稚B 細胞減少、IgD/CD27雙陰性細胞增加、記憶性B 細胞增加及漿母細胞及漿細胞增加[2]。目前有多種以B 細胞為靶向的治療策略,如清除B 細胞及漿細胞(如利妥昔單抗)[3]、選擇性清除活性B 細胞(貝利尤單抗)[4]、阻斷病致病性抗體[abetimus(LJP?394)][5]及阻止B細胞活化等(抗IFN[6]、抗IL?6[7])。這些治療策略的有效性提示B 細胞是SLE 的重要治療靶細胞。
BET(bromodomain and extraterminal)家族蛋白是包含溴區蛋白(bromodomain?containing proteins,BRD)的一類,包括BRD2、BRD3、BRD4 和BRDT 四個成員,作為表觀遺傳閱讀器,參與調控多種免疫反應相關基因的表達。BET 家族成員的異常,往往導致生理及病理過程的失調,在免疫及炎癥性疾病中具有重要的致病作用。因而以其為治療靶點,可調控免疫系統的穩態,進而可能在免疫相關性疾病中發揮治療作用[8]。BRD4 作為研究最為廣泛的BET 蛋白,調節不同的生物過程,包括細胞周期[9]、炎癥[10-11]、維持高級染色質結構[12]和DNA損傷信號[13],在多種刺激下于多種固有免疫細胞及非免疫細胞中表達升高,主要通過激活多種免疫和炎癥基因轉錄發揮促炎作用,進而促進自身免疫性疾病的發生發展[14-15]。筆者既往的研究證實BRD4 參與調控SLE 外周血漿細胞分化過程,抑制劑PFI?1 在狼瘡腎炎模型中具有重要的治療作用[16],但第一代合成的BET 抑制劑JQ1,以模擬乙酰化賴氨酸部分與BET 蛋白家族成員的溴結構域結合發揮抑制BET 蛋白尤其是BRD4 蛋白活性的作用[10,17],目前尚未見其對SLE作用的文獻報道。
本研究擬探討BRD4 蛋白在SLE 外周血B 細胞分化中的作用,擬進一步明確BET 蛋白抑制劑JQ?1 對SLE 外周血B 細胞功能的調控作用。
1 材料與方法
1.1 樣本來源 本研究使用的46 例SLE 患者外周血標本來源于中山大學附屬第一醫院。患者納入標準:(1)女性,年齡19~38 歲;(2)符合1997年修訂的美國風濕病學會(American College of Rheuma?tology,ACR)SLE 分類標準;(3)初發未經激素和/或免疫抑制劑治療,或病情穩定小量激素維持治療的患者即經SLE 疾病活動性指數(SLE disease activity index,SLEDAI[18])?2000 評估為基本無活動或低度活動的患者。排除標準:(1)經SLE 疾病活動性指數(SLE disease activity index,SLEDAI[18])?2000 評估為中度或高度活動的患者;(2)存在其他自身免疫病,心、肝、血液及腫瘤疾病,感染性疾病及其他代謝疾病;(3)妊娠期、哺乳期婦女。實驗符合1975年赫爾辛基宣言(The Declaration of Helsinki)的相關規定,并得到中山大學附屬第一醫院倫理委員會及提供標本的醫院倫理委員會的批準。患者診斷明確,符合各項納入標準,并無排除標準各項情況,自愿同意,簽署知情同意書后納入本項研究,經肘部靜脈采血收集其外周靜脈血標本于肝素抗凝管中。
本研究納入的健康志愿者(HC)為中山大學附屬第一醫院的健康志愿者,其性別、年齡均與納入的SLE 患者匹配,所有參與志愿者均已簽署知情同意書,并得到中山大學附屬第一醫院倫理委員會及提供標本的醫院倫理委員會的批準。
1.2 方法
1.2.1 SLE 外周血PBMC 的提取 采取密度梯度離心(Ficoll?Paque)法分離外周血單個核細胞。取SLE 患者外周靜脈血15~20 mL,PBS(Gibco,美國)稀釋,加入淋巴細胞分離液(GE Healthcare,瑞典)離心,收集白色窄帶層于離心管中,PBS 洗滌2 次,用RMPI?1640 培養基(Gibco,美國)將細胞重懸并定容。
1.2.2 人外周血CD19+B 淋巴細胞磁珠分選純化(陰選) 根據Human B Lymphocyte Enrichment Set?DM(BD,美國)說明書操作,使用BD IMagTMbuffer(BD,美國)重懸細胞,加入Biotinylated Human B Lymphocyte Enrichment Cocktail(BD,美國),室溫孵育15 min,用10 倍體積的BD IMagTMbuffer 洗滌,加入BD IMagTMStreptavidin Particles Plus?DM(BD,美國),室溫孵育30 min,過磁力架6 ~ 8 min,吸取上清(enriched fraction)放入新的流式管,加入1 mL BD IMagTMbuffer 重懸positive?fraction,重新置于磁力架6 ~ 8 min,吸取上清,與enriched fraction 合并,將combined enriched fraction 再過一遍磁力架6 ~ 8 min,吸取上清,放入新的流式管中,為twice?enriched fraction,用于后續實驗。
1.2.3 SLE B 細胞體外培養實驗 將分離純化的B 細胞懸液(1 × 106/mL)以2 × 105/200 μL/孔接種于96 孔平底細胞培養板中,按實驗分組在培養體系中加入(co?stimulate)或不加入(nil)Goat anti?IgM F(ab′)fragment(Jackson,美 國)(5 μg/mL)、Monoclonal mouse anti?CD40L(RD,美 國)(1 μg/mL)、CpG ODN2006(Invivogen,美國)(5 μmol/L)、Recombinant Human IL?4(Pepro Tech,美國)(40 ng/mL),Recombinant Human IL?2(Pepro Tech,美國)(10 ng/mL)共刺激,按實驗分組在co?stimulate 培養體系中不加入(control 組)或加入JQ1(300 nmol/L)(JQ1 組)分別培養7 d 后收集細胞備用。
1.2.4 流式細胞術檢測細胞凋亡 根據凋亡試劑盒(BD,美國)說明書操作,將B 細胞制備成單細胞懸液,收集于5 mL 流式管中,每個樣品加入Annexin V/PI 各50 μL,室溫避光孵育15 min,加入200 μL binding buffer 重懸,30 min 內流式細胞儀檢測(Cyto FLEX,BD,美國)。
1.2.5 流式細胞術檢測細胞增殖 在避光條件下加入CFSE(eBioscience,美國),孵育15 min,終止染色,離心棄上清,保留沉淀,重懸細胞,流式細胞儀檢測。
1.2.6 人外周血漿母細胞體外誘導分化 將分離純化的B 細胞接種于96 孔平底細胞培養板,按實驗分組加入或不加入Goat anti?IgM F(ab′)fragment(Jackson,美國)(5 μg/mL)、Monoclonal mouse anti?CD40L(RD,美國)(1 μg/mL)、CpG ODN2006(Invi?vogen,美國)(5 μmol/L)、Recombinant Human IL?10(Pepro Tech,美國)(50 ng/mL),Recombinant Hu?man IL?15(Pepro Tech,美國)(10 ng/mL)共刺激分別培養7 d 后收集細胞備用。
1.2.7 流式細胞術表面分子染色 收集單細胞懸液,PBS 洗滌,根據實驗設計分別加入0.5 μL Anti?human CD19?APC(Biolegend,美國)、Anti?human IgD?PE?Cy7(Biolegend,美國)、Anti?human CD27?PE(Biolegend,美國)、Anti?human CD38?FITC(Bioleg?end,美國)、Anti?human CD138?Brilliant Violet510(Biolegend,美國),于4 ℃避光孵育30 min,終止染色,離心棄上清,保留沉淀,重懸細胞,流式細胞儀進行檢測。
1.2.8 RT?qPCR 總RNA的提取采用Takara Prime?Script RT Reagent Kit(Takara Bio,日本),并在LC480系統中完成RT?qPCR(Roche,美國)。用2?△△CT檢測目的基因相對于甘油醛?3?磷酸脫氫酶(GAPDH)的表達。BRD4 及GAPDH 引物序列見表1。
1.3 統計學方法 所得數據用SPSS 18.0 軟件進行統計學分析,計量資料用()表示,兩組間計量資料比較使用獨立樣本t檢驗,多組之間使用單因素方差分析,P<0.05 表示差異有統計學意義。
2 結果
2.1 BRD4 蛋白在健康對照組(HC)及SLE 患者外周血B 細胞中的表達 利用RT?qPCR 對HC 及SLE 患者B 細胞中BRD4 蛋白的mRNA 水平進行檢測。結果顯示SLE 外周血B 細胞中BRD4 的表達水平顯著高于HC 組(圖1)。
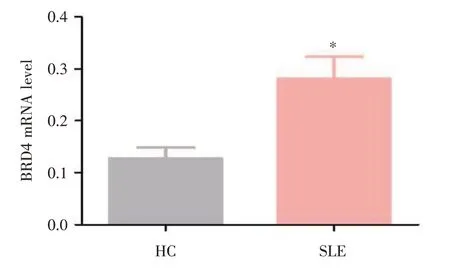
圖1 BRD4 蛋白在HC 及SLE 患者外周血B 細胞中的表達Fig.1 Expression of BRD4 in human peripheral B cells of HC and patients with SLE
2.2 BET 蛋白抑制劑JQ?1 對SLE 外周血B 細胞凋亡的影響 體外培養SLE 外周血B 細胞,通過Annexin V/PI 染色流式細胞術,檢測B 細胞中早期凋亡細胞(Annexin V+PI?)的比例。觀察到在培養7 d 時與陰性對照組(nil)相比,給予共刺激培養條件下B 細胞的早期凋亡細胞的比例同樣明顯減少,而加入300 nmol/L JQ1對細胞的凋亡無影響(圖2)。提示抑制BRD4 蛋白同樣對SLE 外周血活化的B細胞凋亡無影響。
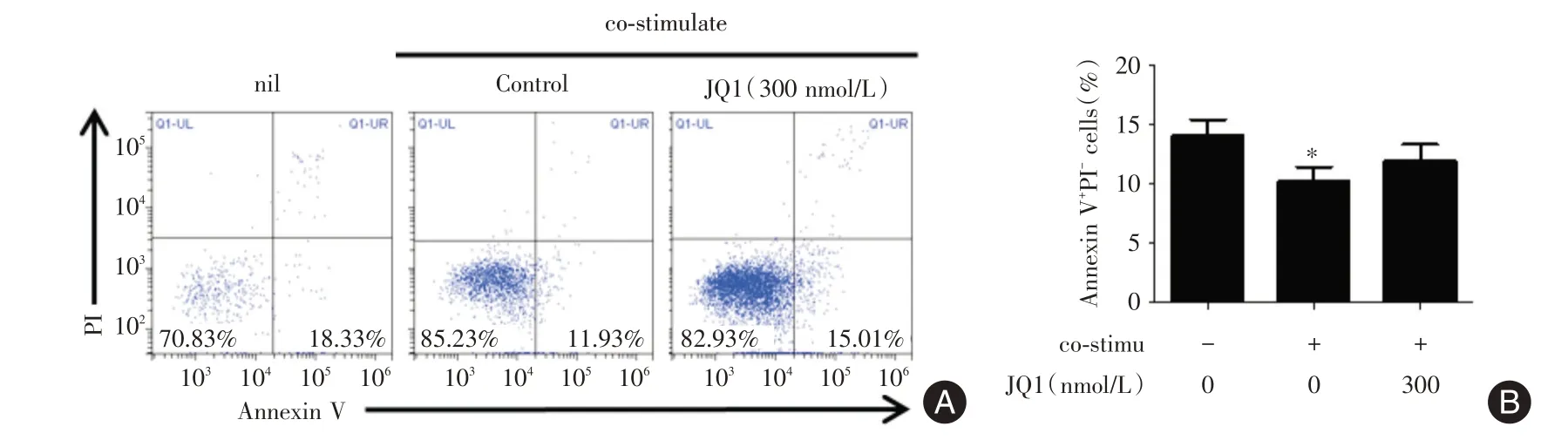
圖2 BET 蛋白抑制劑JQ?1 對SLE 外周血B 細胞凋亡的影響Fig.2 Effect of JQ?1 on the apoptosis of human peripheral activated CD19+B cells
2.3 抑制BRD4 蛋白對SLE 外周血B 細胞增殖的影響 在培養7 d 時與陰性對照組(nil)相比,給予共刺激培養條件下B 細胞的增殖率明顯增加,JQ1處理組對細胞的增殖率無影響(圖3),提示抑制BRD4 蛋白對SLE 外周血活化的B 細胞增殖同樣無影響。
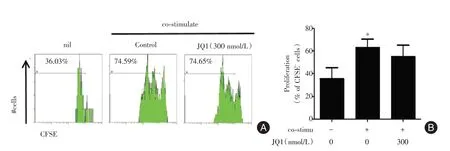
圖3 BET 蛋白抑制劑JQ?1 對SLE 外周血B 細胞增殖的影響Fig.3 Effect of JQ?1 on the proliferation of human peripheral activated CD19+B cells
2.4 BET蛋白抑制劑JQ?1對SLE外周血B細胞各亞群分化的影響 體外培養SLE 外周血B 細胞,通過流式細胞術檢測B 細胞各亞群細胞(幼稚B 細胞、記憶B 細胞、漿母細胞及漿細胞)的百分比。觀察到在培養7 d 時與陰性對照組(nil)相比,給予共刺激培養條件下CD19+IgD+CD27-幼稚B 細胞、CD19+IgD-CD27+記憶B 細胞、CD19+IgD-CD27++漿母細胞、CD19+CD38++CD138+漿細胞亞群的比例均有所升高,而JQ?1 組使升高的漿母細胞、漿細胞亞群的比例受到抑制,且顯著抑制記憶性B 細胞分化,使幼稚性B 細胞亞群的比例顯著增加(圖4),提示抑制BRD4 蛋白同樣能顯著穩定地抑制SLE 外周血漿母細胞途徑介導的漿細胞的分化。
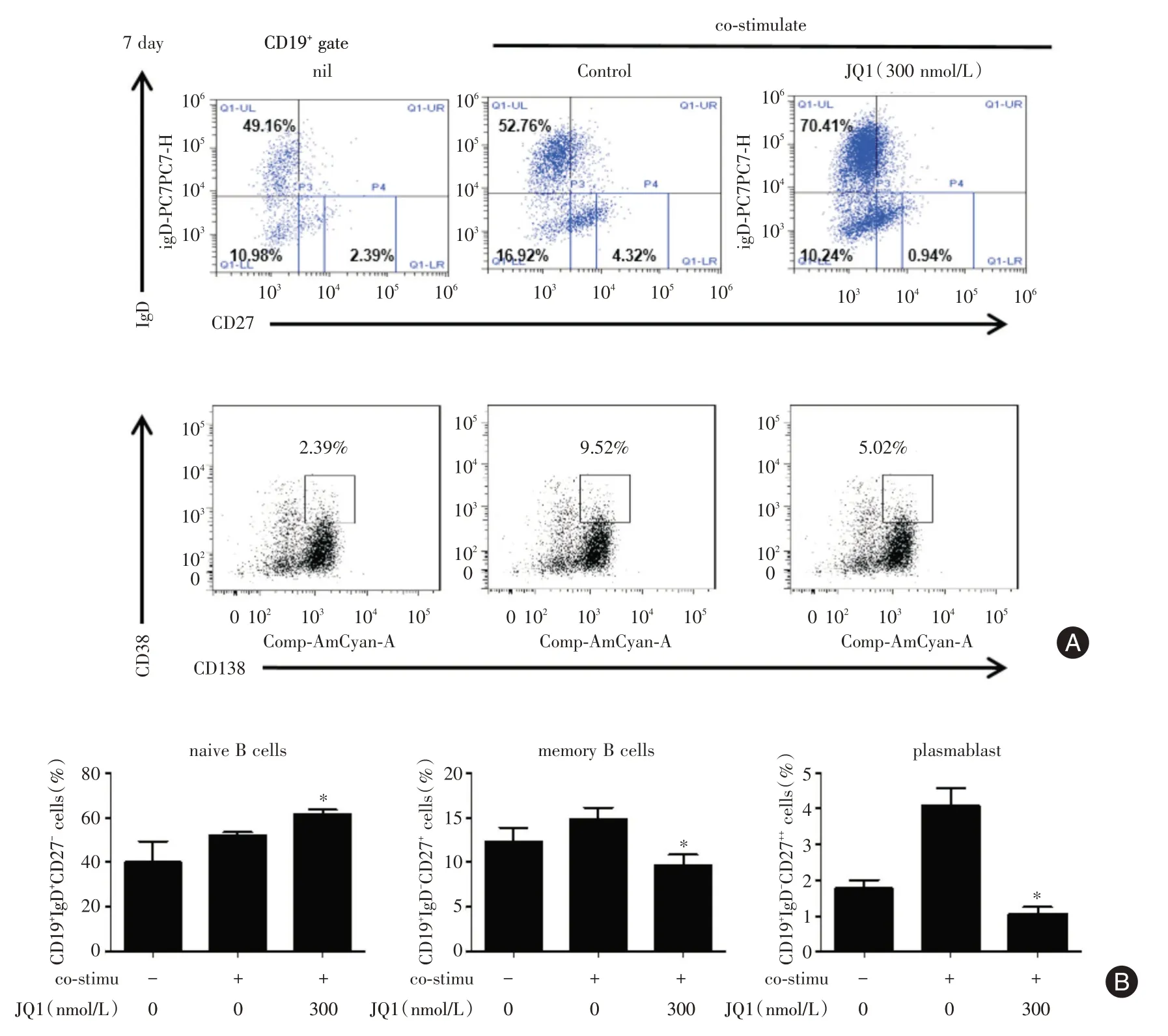
圖4 BET 蛋白抑制劑JQ?1 對SLE 外周血B 細胞各亞群分化的影響Fig.4 Effect of Brd4 inhibition on the percentages of na?ve,memory,plasmablast,and plasma cells in CD19+B cells from patients with SLE
2.5 抑制BRD4 蛋白對SLE 外周血漿細胞分化的影響 為了進一步闡述抑制BRD4 蛋白對SLE 外周血漿細胞分化的研究,利用外周血B 細胞定向誘導分化為大量漿母細胞,通過流式細胞術檢測CD19+IgD-CD38hi漿母細胞產生的比例。觀察到在培養7 d 時與陰性對照組(nil)相比,加入共刺激培養條件后導致SLE 中CD19+IgD-CD38hi漿母細胞比例大量增加,而JQ1 組使漿母細胞的誘導分化作用受到顯著抑制(圖5),進一步提示抑制BRD4 蛋白可以顯著地抑制SLE 外周血漿母細胞途徑介導的漿細胞分化。

圖5 BET 蛋白抑制劑JQ?1 對SLE 外周血漿細胞分化的影響Fig.5 Effect of JQ?1 on differentiation of plasmablasts and plasma cells from SLE patients
3 討論
系統性紅斑狼瘡是一個異質性很強的自身免疫性疾病,其免疫耐受被打破,多種因素導致固有免疫和適應性免疫不同程度障礙,導致免疫系統異常激活及大量致病性自身抗體產生[19]。既往曾有文獻發現,在SLE 患者妊娠晚期存在外周血淋巴細胞亞群的比例失調,SLEDAI 分數與B 細胞比率具有顯著相關性,但與T 淋巴細胞的比率并不具有顯著相關性,提示B 細胞在SLE 發病機制中具有獨特的重要作用[20];此外亦有研究表明SLE風險等位基因主要在髓系或B 細胞中表達,在B細胞靶向療法中,尤其是阻斷漿細胞分化,可能可使SLE 患者獲得長期效益[1]。因此以B 細胞作為靶向細胞是SLE 未來治療的重要方向。
近年研究發現,抑制BRD4 蛋白已被證實在多種腫瘤的臨床前小鼠和異種移植模型中具有顯著的抗癌活性[17,21-22]。除了對腫瘤的調控作用,最近發現BRD4蛋白參與調控人和小鼠體內Th17細胞的分化及活化,調節Th17 相關的細胞因子產生[23-24]。這些研究提示BRD4 蛋白很可能參與調控免疫細胞的活化和分化。但關于BRD4 是否參與B 細胞功能的調控目前研究甚少,曾有學者提出BRD2 蛋白促進小鼠B 細胞擴增和有絲分裂[25];筆者曾發現BRD4 蛋白參與調控SLE 患者外周血B 細胞分化,且抑制劑PFI 對狼瘡腎炎具有治療作用[16];這些提示BET 蛋白尤其是BRD4 蛋白可能在B 細胞介導的自身免疫性疾病發病機制中具有重要的作用,但是BET 蛋白抑制劑JQ?1 對SLE 外周血B 細胞的調控作用,目前尚未見文獻報道。
筆者前期結果發現SLE 患者外周血B 細胞中BRD4 的表達顯著高于健康對照組(HC),提示BRD4 可能參與SLE 的發病機制。為此進一步探討SLE 外周血B 細胞中異常升高的BRD4 蛋白在SLE 漿細胞的分化過程中的作用,觀察發現BET蛋白抑制劑JQ1,對B 細胞的增殖及凋亡沒有顯著的抑制作用,但其可有效降低SLE 外周血漿母細胞和漿細胞的比例,提示JQ?1 可調控SLE 中漿母細胞途徑介導的漿細胞分化過程。
本研究不足之處在于:盡管JQ?1 抑制劑對BRD4 活性的抑制作用具有一定偏好,但其仍可對BRD2、BRD3 及BRDT 蛋白發揮作用,因此不排除JQ?1 通過調控其它BET 蛋白活性發揮作用,因此選擇BRD4 高選擇性抑制劑具有重要意義;此外沒有探究JQ?1 對SLE 外周血B 細胞的調控機制,且未通過動物模型進一步證實其可能的潛在治療作用。
總之,BRD4 在系統性紅斑狼瘡患者外周血B細胞中表達增加,提示BRD4 蛋白在SLE 的發病機制中發揮重要作用,JQ?1 可抑制SLE 外周血漿細胞分化,從而發揮對系統性紅斑狼瘡的治療作用,但精確的調控機制有待進一步深入研究,JQ?1 有望成為SLE 新的潛在治療藥物。
