4種LED光源對1齡赤點石斑魚生長和發(fā)育的影響
2022-06-08 13:55:54徐文剛劉立明王九龍于文松張建柏征矢野清唐永政
水產(chǎn)科學(xué) 2022年3期
關(guān)鍵詞:生長
徐文剛,劉立明,王九龍,于文松,張建柏,征矢野清,唐永政
( 1.煙臺大學(xué) 海洋學(xué)院,山東 煙臺 264005; 2.煙臺市海洋經(jīng)濟研究院,山東 煙臺 264003; 3.長崎大學(xué) 環(huán)中國東海環(huán)境資源研究所,日本 長崎 851-2213 )

魚類成長周期中,內(nèi)分泌系統(tǒng)中的腦—垂體—性腺軸(BPG軸)分泌的激素能夠調(diào)節(jié)其生長、發(fā)育、繁殖、新陳代謝和能量平衡[12]。研究表明,光質(zhì)能改變魚類晝夜節(jié)律,調(diào)節(jié)BPG軸中與生長相關(guān)激素的分泌而影響生長[12-13]。生長激素(GH)在腦垂體中合成,主要調(diào)節(jié)生長和新陳代謝,生長抑素前體-Ⅰ(PSS-Ⅰ)在腦中合成并能抑制生長激素的合成和分泌,神經(jīng)肽Y(NPY)則能刺激食欲并影響晝夜節(jié)律[12]。光質(zhì)通過調(diào)節(jié)魚類GH、PSS-Ⅰ和NPY基因的合成而影響其生長[14]。Shin等[15]報道,綠光和藍(lán)光比紅光更能有效促進(jìn)克氏雙鋸魚(Amphiprionclarkii)中GH基因的合成而加速其生長。
光質(zhì)不僅能影響魚類的生長,也能通過調(diào)節(jié)BPG軸中與繁殖相關(guān)激素的合成而影響其繁殖。已有研究表明,光質(zhì)能調(diào)節(jié)河鱸(Percafluviatilis)[16]和星點東方鲀(Takifuguniphobles)[17]腦垂體中促性腺激素(GtHs)的分泌而影響其性成熟。GtHs基因中的β亞基單元有兩種類型:促卵泡激素(FSHβ)和黃體生成素(LHβ),F(xiàn)SHβ和LHβ基因受到下丘腦中促性腺激素釋放激素(GnRH)的調(diào)節(jié)[12]。近年研究表明,腦中分泌的Kisspeptin激素處于上游調(diào)控階段,能夠調(diào)節(jié)GnRH和GtHs基因的合成和分泌。Kisspeptin基因中的Kiss1和Kiss2基因與其受體G蛋白偶聯(lián)受體54(GPR54)結(jié)合后直接刺激GnRH神經(jīng)元,從而促進(jìn)GtHs基因的合成,其功能與魚類的BPG軸相對應(yīng)[18-19]。Shin等[20-22]報道,光質(zhì)不僅能影響金魚(Carassiusauratus)繁殖系統(tǒng)中各激素的合成和分泌,也能影響紅鰭東方鲀(T.rubripes)的生長和星點東方鲀[17]的成熟。因此,筆者以日本海域較具代表性的赤點石斑魚為研究對象,探究不同光質(zhì)LED光源對其1齡個體生長和性腺發(fā)育的影響;同時也探究不同光質(zhì)對試驗魚腦和腦垂體中與生長相關(guān)的基因如GH、PSS-Ⅰ和NPY,以及與繁殖相關(guān)的基因如Kiss1、Kiss2、GPR54、sbGnRH、FSHβ和LHβ表達(dá)量的影響,以期為未來石斑魚工業(yè)化高效養(yǎng)殖、資源保護(hù)及人工繁殖方面提供基礎(chǔ)生物學(xué)資料和理論依據(jù)。
1 材料與方法
1.1 試驗材料和設(shè)計
取168尾1齡左右赤點石斑魚[體質(zhì)量(42.29±0.77) g,體長(15.6±0.07) cm],其中8尾作為試驗開始前的對照樣本,余下160尾飼養(yǎng)在4個裝有開放式流水系統(tǒng)和曝氣裝置的500 L水槽中,每個水槽放置40尾試驗魚,待其適應(yīng)水溫和攝食等條件后開始試驗,試驗時間為2018年7月24日至9月23日。各組試驗魚暴露在白光色溫3000 K、紅光590 nm、藍(lán)光480 nm、綠光520 nm 4種不同光質(zhì)LED光源下飼養(yǎng)2個月,并在四周覆蓋黑色塑料布以避免自然光干擾。每個水槽放置8個0.72 W的LED燈,固定在距水面22~25 cm處,光照度約580 lx,光照周期為12L∶12D。試驗期間水溫為22.3~28.8 ℃(圖1),鹽度29~31,pH 7.9~8.3,溶解氧>5 mg/L,24 h增氧。試驗期間每日早晚各投喂1次,觀察試驗魚的攝食情況并記錄每日攝食量和水溫的變化,所用飼料為Ohitome EP2 (日清丸紅飼料,日本)。試驗期間無個體死亡。

圖1 7月24日至9月23日試驗期間海水水溫的變化
1.2 采樣方法
1.2.1 體長和體質(zhì)量的測定
試驗開始時、試驗1個月和2個月后,分別測量各水槽中每尾魚的體長和體質(zhì)量,并按下式計算肥滿度(CF)、體質(zhì)量增加率(wWGR,%)和特定生長率(RSG,%/d):
CF=100m/L3
(1)
wWGR=(mt-m0)/m0×100%
(2)
RSG=(lnmt-lnm0)/t×100%
(3)
式中,L為體長,m為體質(zhì)量,mt為終末平均體質(zhì)量,m0為初始平均體質(zhì)量,t為飼育時間。
1.2.2 樣品的采集
試驗開始前對對照樣本中的8尾魚進(jìn)行采樣,試驗結(jié)束時各光質(zhì)組再分別取8尾魚作為試驗樣本。使用2-苯氧乙醇將魚麻醉后,測量其體長和體質(zhì)量,然后取出性腺,稱量質(zhì)量后將其立即置于波恩試劑中,固定24 h后于70%乙醇中保存,用于組織學(xué)分析。隨后打開頭蓋骨,取腦和腦垂體。由于所采集的腦部樣品較大,將腦短暫保存于液氮后移至-80 ℃冰箱中長期保存,腦垂體置于RNAlater試劑(Ambion Inc.,Invitrogen Life Technologies,Japan)中4 ℃低溫保存,1周后廢棄RNAlater試劑,移至-80 ℃冰箱中長期保存,用于分子生物學(xué)試驗。
1.2.3 性腺的組織學(xué)觀察
組織學(xué)分析步驟參照文獻(xiàn)[23]的方法,對赤點石斑魚性腺進(jìn)行常規(guī)梯度乙醇脫水、二甲苯透明、石蠟包埋和連續(xù)切片,厚度為5 μm。采用蘇木精—伊紅染色,中性樹膠封片,Olympus FX380型光學(xué)顯微鏡觀察并拍照,以個體中卵母細(xì)胞最發(fā)達(dá)階段為性腺發(fā)育階段。參照文獻(xiàn)[24-25]的方法對性腺發(fā)育進(jìn)行分期。
1.2.4 總RNA的提取與反轉(zhuǎn)錄
取凍存的赤點石斑魚腦和腦垂體,參照TRIzol試劑盒(Life Technologies Corp.,USA)說明書進(jìn)行總RNA提取,經(jīng)1%瓊脂凝膠電泳檢測RNA完整度,用NanoDrop 2000分光光度計(Thermo Scientific Inc.,美國)檢測RNA濃度。根據(jù)Transcriptor first strand cDNA synthesis(Roche Diagnostic GmbH,Mannheim,德國)說明書操作,取1 μg腦及腦垂體總RNA為反轉(zhuǎn)錄模板合成第一鏈cDNA,用于PCR擴增試驗。
1.2.5 腦和腦垂體中各基因?qū)崟r熒光定量PCR的檢測
腦中PSS-Ⅰ、NPY、Kiss1、Kiss2、GPR54和sbGnRH基因以及腦垂體中GH、FSHβ和LHβ基因表達(dá)量的測定方法為根據(jù)基因庫中各基因序列用Primer 3 Plus軟件設(shè)計熒光定量引物,由Fasmac公司(Kanagawa,日本)合成,引物序列見表1。各基因的測定參照EvaGreen 2×qPCR MasterMix-Rox(Abm Inc.,加拿大)試劑盒上的說明書操作。PCR反應(yīng)體系:cDNA模板2 μL,5 μL EvaGreen 2×qPCR MasterMix,上下游引物(10 pmol/μL)各0.3 μL,2.4 μL滅菌超純水,總計10 μL。在CFX96 TouchTMReal-Time系統(tǒng)上(BioRad,美國)進(jìn)行PCR反應(yīng),條件為:95 ℃預(yù)變性 10 min;94 ℃變性 45 s,58 ℃退火45 s,72 ℃延伸1 min,40個循環(huán);95 ℃ 5 s,65 ℃ 1 min熔解;最后50 ℃冷卻30 s。PCR反應(yīng)后獲得熔解曲線,檢測數(shù)據(jù)用相對定量法進(jìn)行統(tǒng)計分析,β-actin作為內(nèi)參基因。
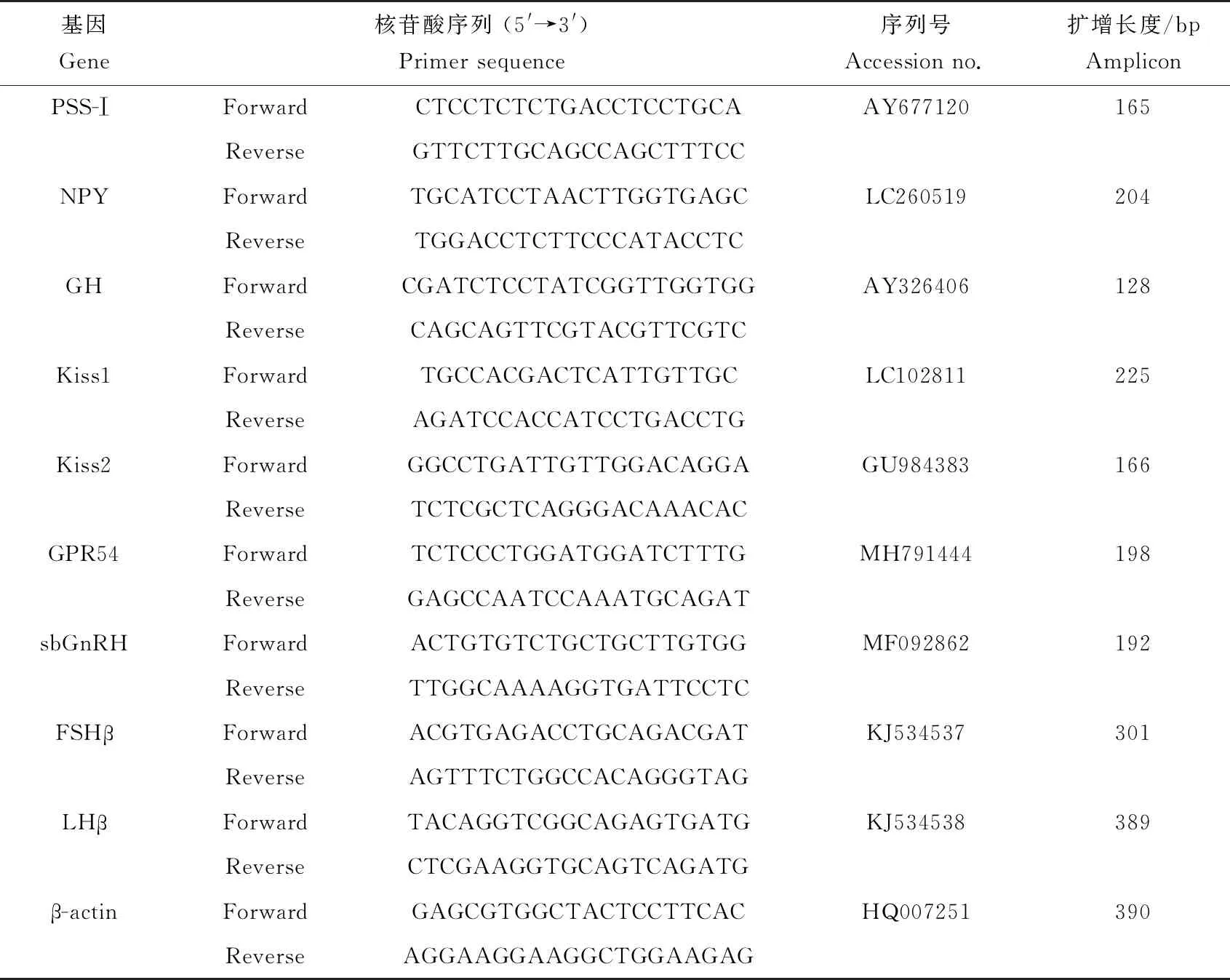
表1 實時熒光定量PCR反應(yīng)中引物序列
1.3 數(shù)據(jù)處理
試驗數(shù)據(jù)以平均值±標(biāo)準(zhǔn)差表示。采用SPSS 22.0軟件對試驗數(shù)據(jù)進(jìn)行比較分析。并用Kolmogorov-Smirnov法進(jìn)行正態(tài)分布檢測,利用單因素方差分析,檢驗體長、體質(zhì)量、肥滿度以及各基因表達(dá)量的差異,運用Tukey′s HSD多重比較檢驗各組數(shù)據(jù)間的差異顯著性,P<0.05時認(rèn)為差異顯著。
2 結(jié)果與分析
2.1 不同光質(zhì)條件下赤點石斑魚生長指數(shù)的差異
試驗期間不同光質(zhì)條件下赤點石斑魚各生長指數(shù)的變化見表2。由表2可見,試驗1個月后白光組魚的體長顯著高于藍(lán)光和綠光組(P<0.05),體質(zhì)量和肥滿度顯著高于其他光質(zhì)組(P<0.05)。試驗結(jié)束時,白光組魚的體長顯著高于藍(lán)光和綠光組(P<0.05),體質(zhì)量顯著高于綠光組(P<0.05),但肥滿度差異不顯著(P>0.05)。試驗結(jié)束時白光組魚的體質(zhì)量增加率高于其他光質(zhì)組。

表2 試驗期間不同光質(zhì)條件下赤點石斑魚各生長指數(shù)的變化
2.2 不同光質(zhì)條件下試驗魚性腺發(fā)育的變化
試驗開始時,試驗魚性腺發(fā)育處于染色質(zhì)核仁期卵母細(xì)胞階段(圖2a)。卵巢內(nèi)可見染色質(zhì)核仁期卵母細(xì)胞,其直徑略大于卵原細(xì)胞。卵母細(xì)胞核較大,占據(jù)卵母細(xì)胞大部分面積。試驗結(jié)束時,各光質(zhì)條件下試驗魚性腺發(fā)育均處于周邊核仁期卵母細(xì)胞階段(圖2b1~b4)。卵母細(xì)胞細(xì)胞質(zhì)體積增加且呈強烈嗜堿性;卵黃核出現(xiàn)且直徑增加,核內(nèi)有少量核仁分布。
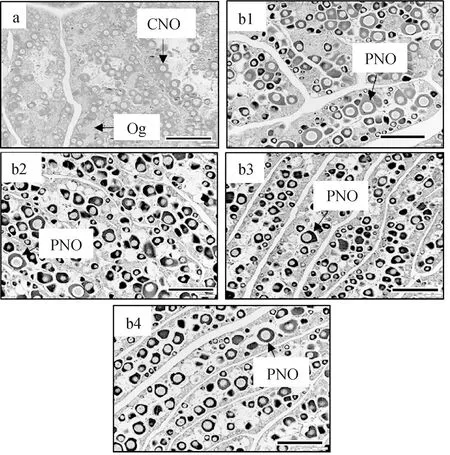
圖2 試驗開始和結(jié)束時各光質(zhì)條件下赤點石斑魚的性腺發(fā)育
2.3 不同光質(zhì)條件下試驗魚腦和腦垂體中與生長和繁殖相關(guān)各基因表達(dá)量的變化
試驗魚腦和腦垂體中PSS-Ⅰ、NPY和GH與生長相關(guān)基因表達(dá)量的變化見圖3。試驗結(jié)束時白光組試驗魚腦中PSS-Ⅰ基因表達(dá)量顯著高于試驗開始時的表達(dá)量(P<0.05);試驗結(jié)束時各光質(zhì)組NPY基因表達(dá)量均顯著高于試驗開始時的表達(dá)量(P<0.05);GH基因表達(dá)量在試驗開始和結(jié)束時差異不顯著(P>0.05)。

圖3 試驗開始和結(jié)束時不同光質(zhì)條件下赤點石斑魚腦和腦垂體中PSS-Ⅰ(a)、NPY(b)和GH(c)基因表達(dá)量的差異
試驗魚腦和腦垂體中Kiss1、Kiss2、GPR54、sbGnRH、FSHβ和LHβ與繁殖相關(guān)基因表達(dá)量的變化見圖4。試驗結(jié)束時,各光質(zhì)條件下試驗魚腦中GPR54基因表達(dá)量均顯著高于試驗開始時的表達(dá)量(P<0.05),白光和藍(lán)光組sbGnRH基因表達(dá)量顯著高于試驗開始時的表達(dá)量(P<0.05),但各光質(zhì)條件下腦中Kiss1基因表達(dá)量在試驗開始和結(jié)束時差異不顯著(P>0.05)。試驗開始時,試驗魚腦中Kiss2基因表達(dá)量太低未檢測到,但試驗結(jié)束時各光質(zhì)條件下的表達(dá)量均有所增加。試驗開始和結(jié)束時,各光質(zhì)條件下試驗魚腦垂體中FSHβ和LHβ基因表達(dá)量差異不顯著(P>0.05)。
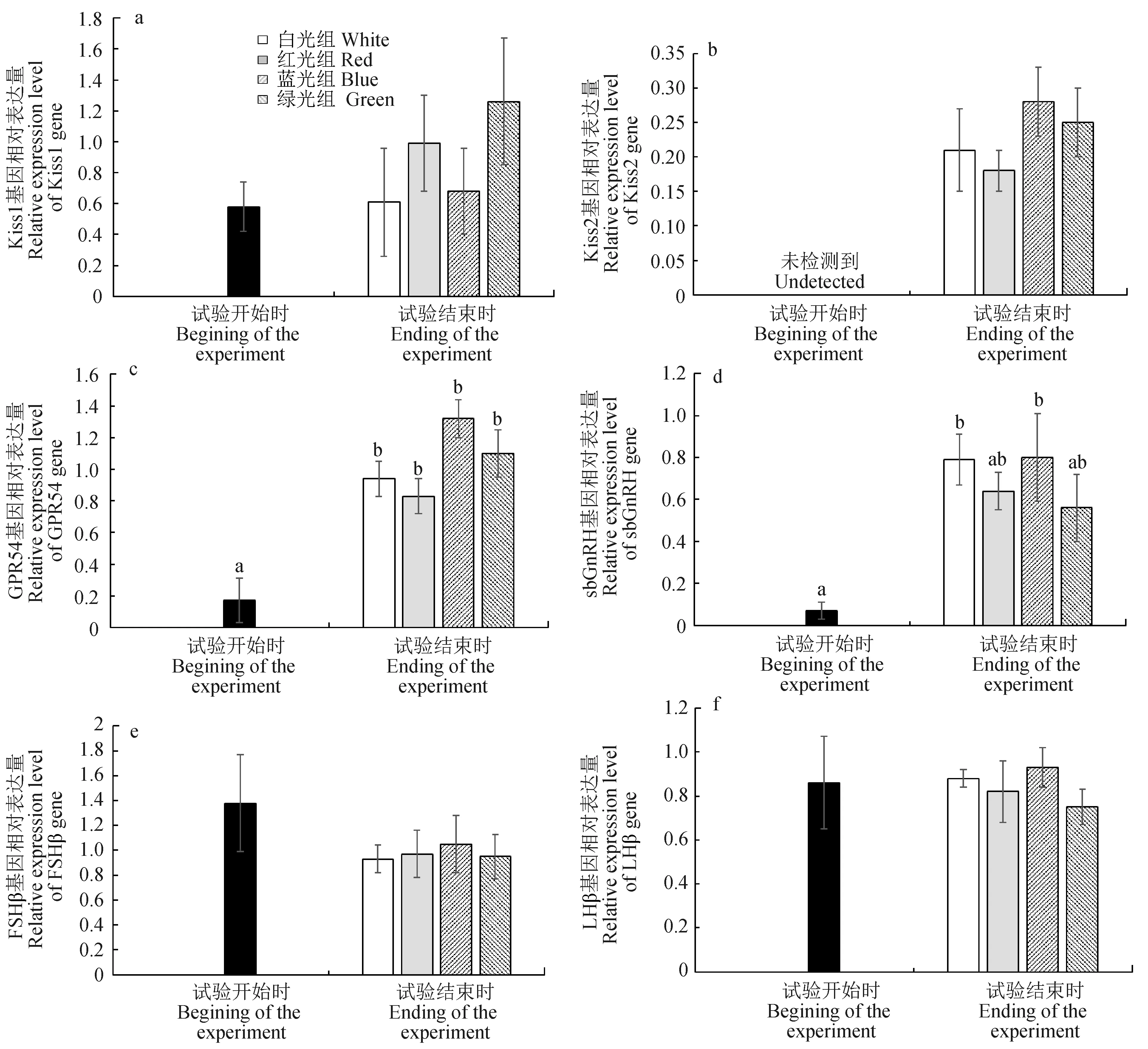
圖4 試驗開始和結(jié)束時不同光質(zhì)條件下赤點石斑魚腦和腦垂體中Kiss1(a)、Kiss2(b)、GPR54(c)、sbGnRH(d)、FSHβ(e)和LHβ(f)基因表達(dá)量的差異
3 討 論
3.1 光質(zhì)對試驗魚生長的影響
本試驗中,赤點石斑魚暴露在不同光質(zhì)下2個月后,其體長、體質(zhì)量和體質(zhì)量增加率順序為白光組>紅光組>藍(lán)光組>綠光組,其中白光組最高,綠光組最低。已有研究表明,水溫13.6~16.7 ℃時,赤點石斑魚暴露在藍(lán)光下10周后比其他光質(zhì)更能促進(jìn)其體質(zhì)量的增加[26]。此外,褐石斑魚(E.bruneus)[27]暴露在綠光下生長率最高,而點帶石斑魚(E.coioides)[28]暴露在紅光下死亡率最低。這些研究結(jié)果表明,當(dāng)石斑魚暴露在不同光質(zhì)下,由于水溫、環(huán)境和物種等因素的差異,光質(zhì)對其生長的影響也有所不同。除石斑魚外,光質(zhì)也能影響其他魚類的生長。研究表明,綠光有利于豹紋鰓棘鱸(Plectropomusleopardus)[13]的攝食、生長和存活,而黃光對紅鰭東方鲀仔稚魚[29]的生長和發(fā)育具有促進(jìn)作用,紅光和綠光不適合其生存。Luchiari等[30]報道,紅光可能對尼羅羅非魚(Oreochromisniloticus)的生長有害;而Volpato等[31]的研究表明,紅光能刺激其進(jìn)食,也有報道表明,藍(lán)光更能促進(jìn)其體長和體質(zhì)量的增加[32]。以上的試驗結(jié)果表明,不同LED光質(zhì)能夠影響魚類的生長,同時,魚類的生長還受魚種、年齡、生理階段、飼養(yǎng)條件和溫度等條件差異的影響。
本試驗中,白光組幼魚肥滿度在第1個月顯著高于其他光質(zhì)組,但2個月后差異卻不顯著。試驗中第1個月的平均水溫為26.7 ℃,此水溫為赤點石斑魚的最適生長條件,而第2個月的平均水溫僅為23.4 ℃。已有研究表明,恒溫26 ℃條件有助于加速赤點石斑魚的生長[10]。對斑點叉尾(Ictaluruspunctatus)[33]和大口黑鱸(Micropterussalmoides)[34]的研究表明,在試驗條件下,高水溫能促進(jìn)其生長。本試驗中可能由于較高的水溫和白光的綜合作用使白光組幼魚在第1個月獲得較高的肥滿度。
3.2 光質(zhì)對試驗魚腦和垂體中與生長相關(guān)基因表達(dá)量的影響
試驗結(jié)束時,各光質(zhì)條件下試驗魚腦中的NPY基因表達(dá)量均顯著高于試驗開始時,表明4種光質(zhì)均能有效促進(jìn)赤點石斑魚的食物吸收、新陳代謝和能量平衡。本試驗中,雖然試驗魚暴露在白光下2個月后其體長和體質(zhì)量均顯著高于其他光質(zhì)條件下,但白光組的GH基因表達(dá)量與其他光質(zhì)條件下和試驗開始時均無顯著差異,而PSS-Ⅰ基因表達(dá)量卻顯著高于試驗開始時。研究表明,GH基因主要用于調(diào)節(jié)魚類的生長,而PSS-Ⅰ基因能夠抑制GH基因的分泌[35]。本試驗中,雖然白光組試驗魚腦中PSS-Ⅰ基因表達(dá)量顯著增加,但其可能對GH基因的分泌沒有很強的抑制作用,因此對試驗魚生長的抑制作用有限。已有研究表明,魚類的生長受到內(nèi)分泌因素如皮質(zhì)醇、甲狀腺激素以及與繁殖相關(guān)激素的綜合影響,導(dǎo)致生長激素復(fù)雜的調(diào)節(jié)機制[36]。因此,進(jìn)一步的研究需明確GH和PSS-Ⅰ基因具體的調(diào)節(jié)機制。
3.3 光質(zhì)對試驗魚性腺發(fā)育以及腦和垂體中與繁殖相關(guān)基因表達(dá)量的影響
試驗結(jié)束時,各光質(zhì)條件下試驗魚性腺從染色質(zhì)核仁期卵母細(xì)胞發(fā)育至周邊核仁期卵母細(xì)胞,且各組試驗魚的GPR54基因表達(dá)量以及白光和藍(lán)光組sbGnRH基因表達(dá)量均顯著高于試驗開始時,而FSHβ和LHβ基因在試驗開始和結(jié)束時的表達(dá)量無顯著差異。研究表明,內(nèi)分泌系統(tǒng)中的Kiss、GnRH和GtHs基因在赤點石斑魚[10]、星點東方鲀[37]、歐洲舌齒鱸(Dicentrachuslabrax)[38-39]和海鱸(Moronesaxatilis)[40]成熟和繁殖過程中起重要作用。此外,在鮭魚[41]、星康吉鰻(Congermyriaster)[42]、鰻鱺(Anguillajaponica)[43]和歐洲舌齒鱸[44]中,F(xiàn)SHβ基因?qū)π韵僭缙诎l(fā)育如精子和卵子的生長具有重要影響,而LHβ基因主要調(diào)控性腺發(fā)育的成熟階段如排卵和排精等行為,表明FSHβ和LHβ基因與多數(shù)魚類的性腺發(fā)育密切相關(guān),且主要作用于青春期和成熟期。Tanaka等[45]報道,在人工養(yǎng)殖條件下,赤點石斑魚在2齡進(jìn)入青春期,3齡達(dá)到性成熟。而本試驗中的赤點石斑魚僅為1齡,性腺尚處于初級卵母細(xì)胞發(fā)育階段,兩者均遠(yuǎn)離成熟期。因此,與BPG軸功能相對應(yīng)的Kiss、GnRH和GtHs基因的角色和功能在此階段可能并不明顯,即使GPR54和sbGnRH基因表達(dá)量顯著增加,可能也無法有效地誘導(dǎo)FSHβ和LHβ基因的合成與分泌,導(dǎo)致FSHβ和LHβ基因表達(dá)量無顯著差異。類似結(jié)果也出現(xiàn)在對河鱸的研究中,當(dāng)未成熟的雄性和雌性河鱸暴露在白光、紅光、綠光或藍(lán)光下10 d后,F(xiàn)SHβ和LHβ基因表達(dá)量無顯著差異,原因可能是魚的繁殖周期尚未開始[16]。然而,當(dāng)金魚暴露在白光、紅光和綠光下4個月后,綠光下金魚腦和垂體中Kiss1、GPR54、GnRH、FSHβ和LHβ基因表達(dá)量卻顯著增加,其性腺也處于成熟卵母細(xì)胞發(fā)育階段,表明Kiss1、GPR54、GnRH、FSHβ和LHβ基因在其繁殖系統(tǒng)中起重要作用[20]。因此,進(jìn)一步的研究需明確不同光質(zhì)LED光源對成熟期赤點石斑魚繁殖系統(tǒng)發(fā)育的影響。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
