一個靈芝新品種的選育與分子鑒定*
2022-06-08 11:48:58叢倩倩唐麗娜李秀梅安秀榮
中國食用菌 2022年5期
叢倩倩,崔 曉,唐麗娜,李秀梅,安秀榮
(泰安市農業科學院,山東 泰安 271000)
靈芝(Ganoderma lingzhi)屬真菌界(Fungi)擔子菌亞門(Basidiomycota)傘菌綱(Agaricomycetes)多孔菌目(Polyporales)靈芝科(Ganodermataceae)靈芝屬(Ganoderma),是一種名貴的藥用真菌,具有抗腫瘤、護肝、抗病毒、抑菌、抗氧化等多種藥效[1-5],被稱為“仙草”。
近年來,隨著靈芝藥用功效被不斷發現,人們對靈芝及其產品的需求量大幅增加,靈芝產業化、規模化發展迅速[6-7]。我國是靈芝出口大國,但栽培品種大多由國外引進或野生菌株分離得到,具有自主知識產權的品種較少[8]。這不僅對我國靈芝產業的發展構成威脅,還會對其他國家的品種產生依賴性,同時也浪費了我國豐富的靈芝種質資源。此外,大多育種的目標更注重產量因素而忽視了功能性指標,導致有效成分含量高的品種較少,且菌種退化的現象比較嚴重,這些問題均制約了我國靈芝產業的發展[9]。因此,選育具有自主知識產權且有效成分含量高的優質高產新品種,提高品種的功能性,對滿足國民營養健康需求,加快推進供給側結構優化和品種換代改良具有重大意義,是促進我國靈芝產業健康發展的重要基礎。
以國家認定品種泰山赤靈芝1號(TL-1)和野生靈芝菌株4895作為親本,通過原生質體單核化雜交育種技術開展靈芝新品種的選育工作,旨在為靈芝生產提供優質高產的新菌株,提升產品的市場競爭力。
1 材料與方法
1.1 供試材料
親本菌株TL-1、4895于泰安市農業科學院食用菌研究所保藏。TL-1為國家認定品種,子實體多糖含量較高,綜合農藝性狀較好;4895由采自泰山的野生靈芝菌株馴化而來,其子實體三萜含量較高。
1.2 親本原生質體的制備與再生
分別制備2個親本的原生質體,用液體再生培養基適當稀釋后涂布于固體再生培養基平板上,至長出再生菌落[10-11]。
1.3 單核菌株的篩選及交配型測定
從親本原生質體再生平板上挑取菌落小、生長緩慢的單菌落,分別轉接到PDA培養基中,待菌落長至3 cm~5 cm時,挑取邊緣菌絲鏡檢,無鎖狀聯合的即為單核菌株。隨機選取一株單核菌株作為標準菌株,將其他單核菌株分別與該標準菌株接種到同一個PDA培養基平板上進行對峙培養,當2個菌株的菌絲相互接觸后,挑取交接處菌絲鏡檢。有鎖狀聯合則說明這2個單核菌株為不同的交配型;無鎖狀聯合則說明兩者不親和,為相同的交配型。
1.4 雜交菌株的選育過程
用直徑0.5 cm的金屬打孔器分別將具有不同交配型的2個單核菌株接種到同一個PDA培養基平板上,兩者相距2 cm,25℃恒溫箱對峙培養7 d;待菌絲相互接觸后,挑取交接處的菌絲鏡檢,有鎖狀聯合則說明雜交成功;將雜交成功組合進行拮抗反應驗證,共獲得雜交菌株5個。2018年,將5個雜交菌株分別進行栽培比較試驗,發現編號為TL-3的雜交菌株菌蓋中等偏大且厚,形態較飽滿,產量、子實體有效成分含量均高于親本菌株TL-1。2019年至2021年,進行了連續3年的栽培試驗,發現TL-3菌株遺傳穩定性較好,綜合農藝性狀優良,具有較高的應用推廣價值。
1.5 優良菌株的ISSR分子標記鑒定
分別提取親本菌株TL-1、4895和雜交菌株TL-3的菌絲體基因組DNA[12],參考NY/T 1730-2009食用菌菌種真實性鑒定ISSR法[13]中推薦的28條引物分別對供試菌株進行ISSR-PCR擴增。PCR產物進行1.5%的瓊脂糖凝膠電泳,根據擴增條帶的穩定性和多態性篩選出適合用于ISSR分析的引物。從篩選出的適宜引物中挑選擴增效果較好的引物作為ISSR分子標記驗證優良雜交菌株的引物。
1.6 優良菌株的菌絲生長特征分析
取大小約0.5 cm×0.5 cm的親本和雜交株的菌絲塊,分別接種于3個PDA平板培養基上,置于25℃恒溫箱中避光培養5 d;觀察親本和雜交株的菌落形態,并測量菌絲的生長速度,每個處理3次重復。
1.7 優良菌株的綜合農藝性狀分析
發菌期,觀察和記錄親本和雜交菌株的栽培袋菌絲生長情況。子實體生長期,拍照記錄不同生長時期的子實體形態特征。待菌蓋邊緣白色生長點消失,菌蓋背面開始彈射霧狀紅褐色孢子時,開始采收。采收后,親本和雜交菌株各取30個靈芝子實體,分別測量菌蓋長直徑、菌蓋厚度、菌柄長度;將子實體曬干,統計產量并計算生物學效率,生物學效率為子實體干重(g)與培養料干重(g)之比乘以100%,每個處理3次重復。
1.8 優良菌株的有效成分含量檢測
各取10個曬干的靈芝子實體,分別檢測親本和雜交株子實體中多糖和三萜的含量[14-15],每個處理3次重復。
1.9 數據分析
采用DPS軟件對以上數據進行統計學分析。
2 結果與分析
2.1 親本原生質體的制備與再生
親本菌絲體通過溶壁酶降解除去細胞壁后,離心收集原生質體,并在顯微鏡下分別拍照記錄原生質體的形態,見圖1。
圖1 親本菌株TL-1及4895的原生質體Fig.1 Protoplasts of parental strains TL-1 and 4895
2.2 單核菌株的篩選及交配型測定
通過鏡檢親本原生質體再生菌落中鎖狀聯合的有無,可以篩選到2種交配型的單核菌株。篩選獲得TL-1單核菌株共21個,其中2種不同交配型單核體的比例為16∶5;篩選獲得4895單核菌株23個,其中僅獲得1種交配型單核體。
2.3 優良雜交菌株的鑒定
對菌株進行拮抗試驗,結果見圖2。

圖2 親本菌株TL-1、4895及雜交株TL-3的拮抗現象Fig.2 Antagonism of parental strains TL-1,4895 and hybrid strain TL-3
如圖2所示,雜交菌株TL-3與親本菌株TL-1、4895之間均存在明顯的拮抗現象,證明TL-3為真正的雜交株。
菌株TL-1、4895和TL-3的ISSR擴增圖譜見圖3。
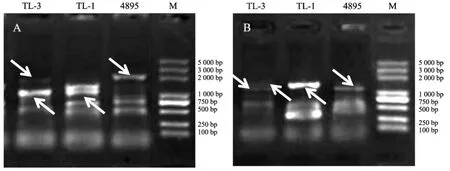
圖3 親本菌株TL-1、4895及雜交株TL-3的ISSR擴增圖譜Fig.3 ISSR amplification map of parental strains TL-1,4895 and hybrid strain TL-3
如圖3所示,ISSR分子標記驗證中,引物P4、P9對雜交株和親本菌株的擴增效果較好,親本菌株與雜交株間的特異性條帶較明顯。雜交株TL-3擴增條帶中同時包含了親本菌株TL-1和4895的特異性條帶,從而在分子水平上進一步證明了TL-3雜交株的真實性。
2.4 優良菌株的菌絲生長特性分析
對親本菌株TL-1、4895及雜交株TL-3進行培養,其菌落形態見圖4,菌絲生長情況見表1。
由圖4和表1可知,雜交菌株TL-3菌絲潔白,呈絨毛狀,濃密,長勢強壯。TL-3平板培養的菌絲生長速度介于2個親本之間,為0.74 cm·d-1,快于親本菌株4895,慢于親本菌株TL-1,與2個親本菌株相比差異極顯著(P<0.01)。

表1 親本菌株TL-1、4895及雜交株TL-3的菌絲生長情況比較Tab.1 Comparison of mycelial growth between parental strains TL-1,4895 and hybrid strain TL-3
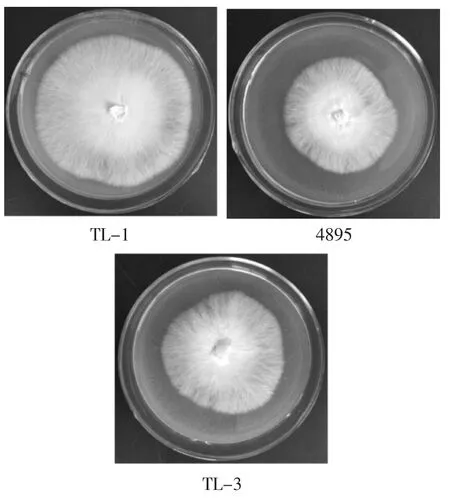
圖4 親本菌株TL-1、4895及雜交株TL-3的菌落形態Fig.4 Mycelial morphology of parental strains TL-1,4895 and hybrid strain TL-3
2.5 優良菌株的綜合農藝性狀分析
統計親本菌株TL-1、4895及雜交株TL-3的農藝性狀,結果見表2。
由表2可知,TL-3栽培袋中菌絲的發菌速度為0.62 cm·d-1,介于2個親本之間;菌絲長勢強壯、濃密,發菌情況與雙親相比均無顯著性差異。菌蓋中等偏大,平均長直徑為10.33 cm,小于2個親本菌株,與親本TL-1之間有顯著性差異(P<0.05),與4895之間無顯著性差異。TL-3菌蓋厚度為2.79 cm,比雙親的菌蓋厚,與TL-1之間差異極顯著(P<0.01),與4895之間無顯著性差異。TL-3菌柄長度為2.06 cm,短于雙親,但無顯著性差異。TL-3平均單產和生物學效率比2個親本菌株高,與親本TL-1之間有顯著性差異(P<0.05),與4895之間無顯著性差異。
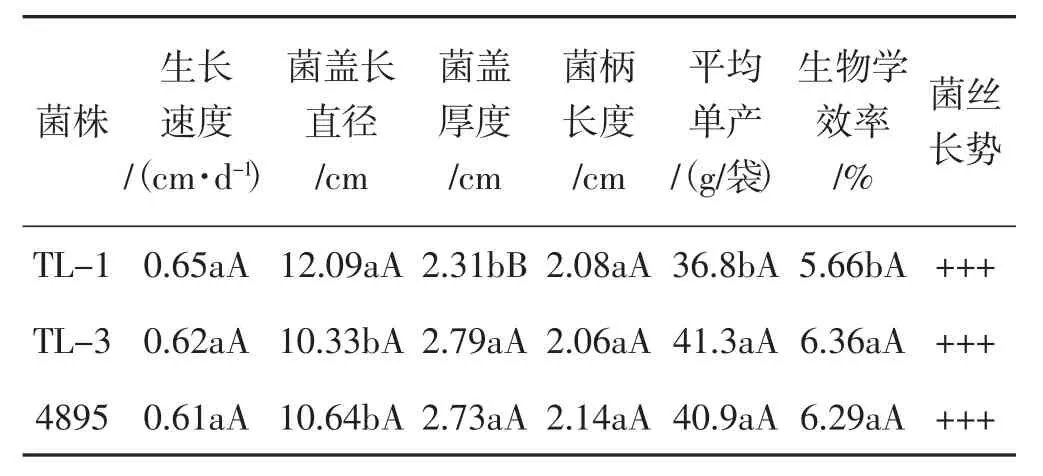
表2 親本菌株TL-1、4895及雜交株TL-3的農藝性狀比較Tab.2 Comparison of agronomic characters between parental strains TL-1,4895 and hybrid strain TL-3
對比親本菌株TL-1、4895及雜交株TL-3不同生長時期的子實體形態,結果見圖5(左側圖為正面,右側圖為背面)。
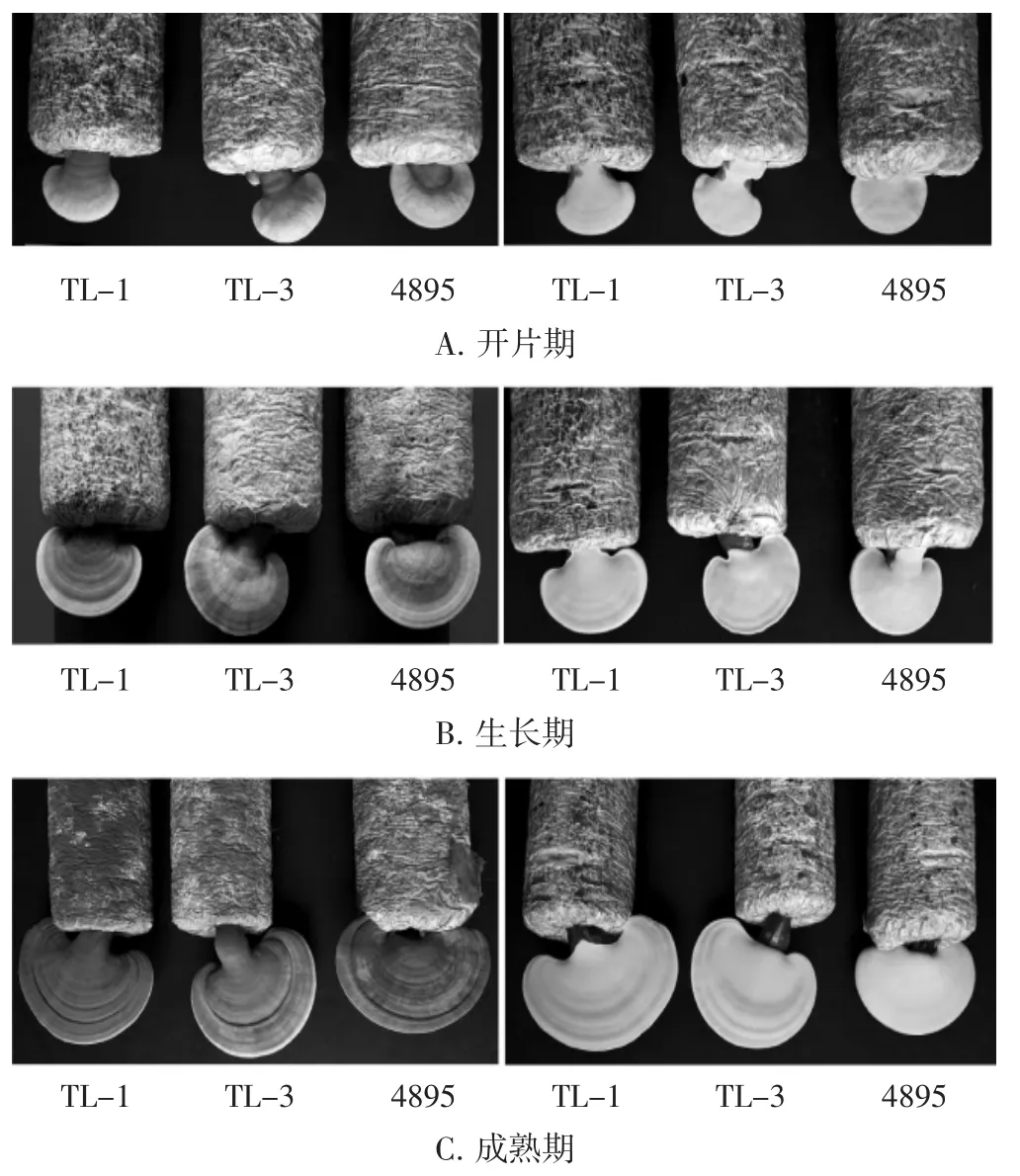
圖5 親本菌株TL-1、4895及雜交株TL-3不同生長時期的子實體正反面形態Fig.5 Positive and negative morphology of fruit bodies of parental strains TL-1,4895 and hybrid strain TL-3 at different growth stages
如圖5所示,TL-3子實體腎圓形,正面黃褐色到紅褐色,有光澤,表面有明顯的放射性縱脊和同心環紋,反面黃色,菌蓋邊緣較圓鈍;菌肉淡褐色;菌柄扁圓柱狀,光滑且亮,紅褐色至紫褐色。
2.6 優良菌株的有效成分含量檢測
檢測親本菌株TL-1、4895及雜交株TL-3的多糖、三萜含量,結果見表3。

表3 親本菌株TL-1、4895及雜交株TL-3子實體有效成分含量比較Tab.3 Comparison of effective components in fruit bodies of parental strains TL-1,4895 and hybrid strain TL-3
由表3可知,雜交菌株TL-3子實體多糖含量為1.09%,均高于親本,比TL-1提高1.87%,比4895提高29.76%,與親本TL-1之間無顯著性差異,與親本4895之間有極顯著差異(P<0.01)。子實體三萜含量為0.22%,介于2個親本之間,比親本菌株TL-1提高37.5%,與2個親本之間均有極顯著性差異(P<0.01)。
2.7 優良菌株的遺傳穩定性分析
將雜交株TL-3進行連續3年的栽培試驗,并記錄每年的栽培農藝性狀,統計結果見表4。

表4 雜交株TL-3的遺傳穩定性分析Tab.4 Genetic stability analysis of hybrid strain TL-3
由表4可知,雜交株TL-3菌絲生長速度為0.60 cm·d-1~0.65 cm·d-1,平均單產為 40.5 g/袋~43.7 g/袋,生物學效率>6%,子實體中多糖和三萜的含量分別為1.07%~1.13%和0.21%~0.27%。每年的栽培農藝性狀之間及與初代之間在平均生長速度、平均單產、生物學效率、子實體多糖及三萜含量等方面基本一致,說明雜交菌株TL-3遺傳穩定性較好。
3 討論
親本TL-1為國家認定品種,其綜合農藝性狀較好,子實體多糖含量較高,但三萜含量較低;且TL-1作為主栽品種之一,經過連續多年的栽培,菌種退化現象比較嚴重。親本4895子實體片厚且三萜含量較高,但產量低。經原生質體單核化雜交選育的新品種TL-3較好地遺傳了雙親的優良性狀,其出芝整齊、片厚、產量高、子實體多糖和三萜含量均較高,綜合農藝性狀和功能性指標均優于國家認定品種TL-1,具有較高的推廣應用價值。此外,新品種的選育還有效解決了菌種退化問題,為生產企業替換老化、退化的品種提供了備選種質資源,進一步滿足了市場需求。
通過分別提取親本和新品種的基因組DNA,利用ISSR法,從28條引物中篩選出2條擴增效果較好的引物作為ISSR分子標記驗證雜交菌株的引物。進一步從分子水平上證明了新品種的真實性,為將來申報新品種權提供了依據。
