海拔梯度對賀蘭山巖羊主要活動區植物群落特征的影響
2022-06-11 01:59:34趙朋波邱開陽謝應忠劉王鎖李小偉陳林王繼飛孟文芬黃業蕓李小聰楊浩楠
草業學報 2022年6期
趙朋波,邱開陽*,謝應忠,劉王鎖,李小偉,陳林,王繼飛,孟文芬,黃業蕓,李小聰,楊浩楠
(1. 寧夏大學農學院,寧夏 銀川 750021;2. 寧夏大學生態環境學院,寧夏 銀川 750021;3. 寧夏大學西北土地退化與生態恢復省部共建國家重點實驗室培育基地,寧夏 銀川 750021;4. 寧夏賀蘭山國家級自然保護區管理局,寧夏 銀川 750021)
賀蘭山地處阿拉善高原和銀川平原之間,是我國重要地理分界線,區隔著我國季風區與非季風區、溫帶草原與荒漠草原[1?2],也是青藏高原、蒙古高原及華北平原植物區系的重要樞紐[3],是干旱內陸地區為數不多的富有植物多樣性的地區之一[4]。賀蘭山作為我國西北地區一道重要的生態屏障,既阻礙了騰格里沙漠的東進,又攔截了來自烏蘭布和沙漠的風沙,有效地保護了銀川平原[3]。因此,賀蘭山對于維護中國西北乃至整個西部地區的生態安全具有重要的戰略意義。
在賀蘭山,由于雪豹等天敵消失[5]、禁牧措施等原因導致種間競爭減弱[6],使得巖羊(Pseudois nayaur)數量快速增長[6],這種單一物種種群快速增長誘發了諸多問題,造成了與同等生態位上其他物種的關系失衡[7],嚴重影響到植被的恢復和其他動物的發展[8]。巖羊作為植物?巖羊?雪豹這一食物鏈中重要的一環,一旦超過自然界的承載能力,勢必引起賀蘭山生態環境的破壞,如草場面積銳減、土地沙漠化加劇等問題[9]。
賀蘭山自然保護區的植被垂直分異特征明顯,其植被類型沿著海拔上升依次為荒漠草原、山地疏林草原、山地針葉林和高山?亞高山灌叢草甸[4,10?11]。賀蘭山東坡是巖羊的主要分布區[6]。賀蘭山巖羊主要分布在海拔為1100~2800 m 的地區,這些地區多是以灰榆(Ulmus glaucescens)等喬木為主的山地疏林草原,或無高大喬木,或接近水源,并遠離人類活動干擾,距離裸巖2~5 m,位于半陰半陽坡的下坡位[12?13]。另外還有高山?亞高山灌叢草甸[14?15]也是巖羊偏愛的取食地和棲息地。
在山地生態系統中,海拔對其生態?水文過程有著強烈的影響,這也導致了植物群落特征隨海拔變化而有所不同[16]。同時,與海拔密切相關的氣候的變化也對植物群落有制約作用,也是植物多樣性變化的主要驅動因子[17]。現有研究表明,山地植物物種豐富度的模型主要有兩種:一是物種豐富度隨海拔升高而升高[18],二是隨海拔的升高,物種豐富度先升高后降低[19]。這是與海拔相關的水分、溫度等環境因子的變化所導致。在不同海拔地區,水分主導了植物的生長,水分對植物的限制性隨著海拔的降低而增強。總的來說,海拔變化囊括了水分、溫度、光照、濕度和土壤等環境因子的變化,對植物群落的多樣性存在顯著影響[20]。
為了探明賀蘭山巖羊主要活動區的植物群落特征,進而為理清巖羊與植被的平衡關系奠定基礎,以期加強對賀蘭山地區巖羊種群的合理管護,從而維護賀蘭山生態平衡,本研究以位于賀蘭山東坡的巖羊主要活動區植被群落為對象,通過對不同海拔的巖羊主要活動區進行植被調查,研究其物種組成、物種多樣性變化,試圖說明:1)賀蘭山巖羊主要活動區不同植被類型的物種組成;2)植物物種多樣性沿海拔梯度的變化規律。
1 材料與方法
1.1 研究區概況
研究區位于寧夏賀蘭山國家級自然保護區蘇峪口。賀蘭山橫亙于阿拉善高原和銀川平原之間,東臨黃河,西靠騰格里沙漠,屬于干旱半干旱地區。地理坐標為38°13′?39°30′ N,105°41′?106°41′ E,是我國季風區與非季風區、內流區與外流區的地理分界線。賀蘭山大體為西南?東北走向,東西寬20~40 km,南北約180 km,東坡山勢陡峭,西坡相對平緩,從山麓到山頂垂直分化明顯,高度相差2000 m 以上,主體海拔大多在2000~3000 m 之間,主峰敖包疙瘩高3556.1 m。目前建立了寧夏賀蘭山國家級自然保護區(東坡)和內蒙古賀蘭山國家級自然保護區(西坡)。
賀蘭山屬于典型的大陸性季風氣候,冬季漫長且寒冷,夏季短暫而炎熱,氣候較為干燥,全年盛行西北風,年平均風速7.5 m·s?1,大風天數達157.7 d,最大風速38.7 m·s?1。賀蘭山的水熱條件隨著海拔上升有著明顯的差異,東坡年平均氣溫8.2 ℃,年平均降水量183.3 mm,西坡年均氣溫7.6 ℃,年均降水量430 mm,降水主要集中在6?9月,按水熱條件,賀蘭山地區可被劃分為4 個垂直氣候帶:溫暖干旱草原荒漠氣候帶(<1500 m)、較溫涼半干旱草原氣候帶(1600~2000 m)、溫寒半干旱偏濕潤森林氣候帶(2000~3000 m)、寒冷半濕潤高山?亞高山氣候帶(>3000 m)。主要植被類型隨著海拔升高逐漸由山地草原帶(1000~1600 m)、山地疏林草原帶(1600~2000 m)過渡到山地針葉林帶(2000~2500 m)和高山?亞高山灌叢、草甸帶(>2500 m)[21]。
1.2 樣地設置
于2020年8月在賀蘭山國家級自然保護區蘇峪口開展試驗。結合以往在賀蘭山開展的與巖羊相關的研究[6,12]及實地勘察,并根據植被類型情況,確定賀蘭山巖羊主要活動區4 個,從山麓到山頂分別為荒漠草原、山地疏林草原、山地針葉林、高山?亞高山灌叢草甸(表1)。在每個活動區設置3 個樣地,每個樣地布設具有代表性的套樣方,即3 個喬木樣方(20 m×20 m)內布設灌木樣方(10 m×10 m)5 個,每個灌木樣方內各設置1 個草本樣方(1 m×1 m),樣方均設置在陽坡。運用植物群落學調查技術,調查植物種類、高度、蓋度、頻度、多度以及地上生物量,灌木和喬木植物補充測定其冠幅、胸徑等[22]。
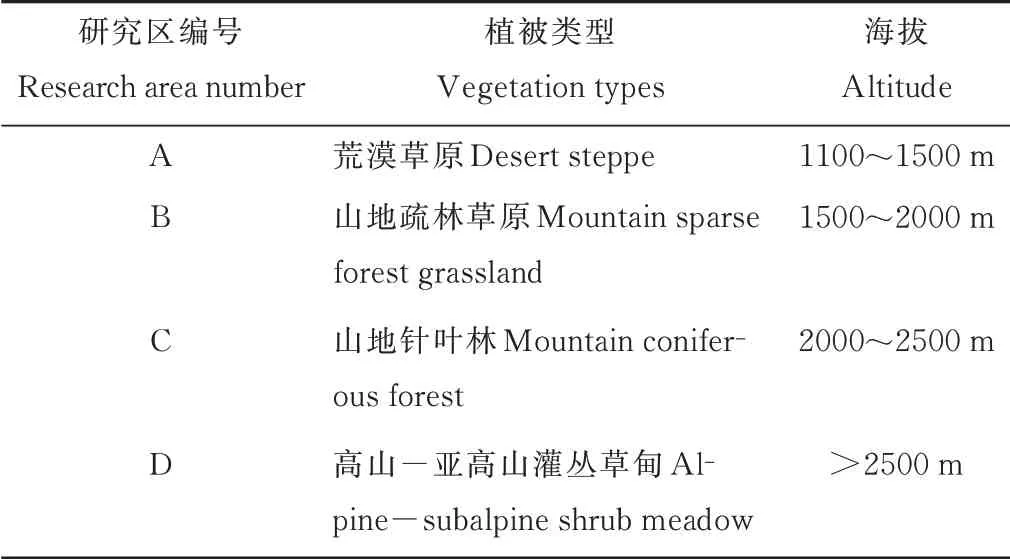
表1 研究區植物群落類型及海拔高度Table 1 Plant community types and the altitude of research areas
1.3 數據分析
物種多樣性指數采用物種豐富度指數(R)、Simpson 優勢度指數(D)、Shannon?Wiener 多樣性指數(H′)和Pielou 均勻度指數(E),根據以下公式計算不同植物群落數量特征:

式中:S為群落中物種總數;Pi為種i的相對重要值,i=1,2,……,S。
本研究首先使用Excel 2019 對數據進行初步整理、計算各研究區內植物的重要值和多樣性指數,然后利用SPSS 19.0 進行單因素分析(One-way ANOVA)和回歸分析,對不同海拔的數據進行差異顯著性檢驗、多重比較和回歸分析等,所有圖表均在Excel 2019 中完成。
2 結果與分析
2.1 賀蘭山巖羊主要活動區植被組成及區系特征
賀蘭山巖羊主要活動區共記錄到30 科62 屬87 種植物。其中被子植物84 種,裸子植物3 種。賀蘭山巖羊主要活動區所有植物中有3 個科所含種的數量在10 種以上(表2),其中含種數量最多的是禾本科(Poaceae),有20種,占總種數比例為22.98%,其次是薔薇科(Rosaceae)(11 種)和菊科(Asteraceae)(10 種),兩科合計占總種數比24.14%,然后依次是豆科(Fabaceae)(7 種)、石竹科(Caryophyllaceae)(4 種)、傘形科(Apiaceae)(3 種)、玄參科(Scrophulariaceae)(3 種)、莧科(Amaranthaceae)(3 種)、石蒜科(Amaryllidaceae)(2 種)、旋花科(Convolvulaceae)(2種)、毛茛科(Ranunculaceae)(2 種)、唇形科(Lamiaceae)(2 種)。另外還有18 個單種科,包括遠志科(Polygalaceae)、鳶尾科(Iridaceae)、忍冬科(Caprifoliaceae)、茜草科(Rubiaceae)、榆科(Ulmaceae)、小檗科(Berberidaceae)和松科(Pinaceae)等,所有單種科數占總科數的60.00%。
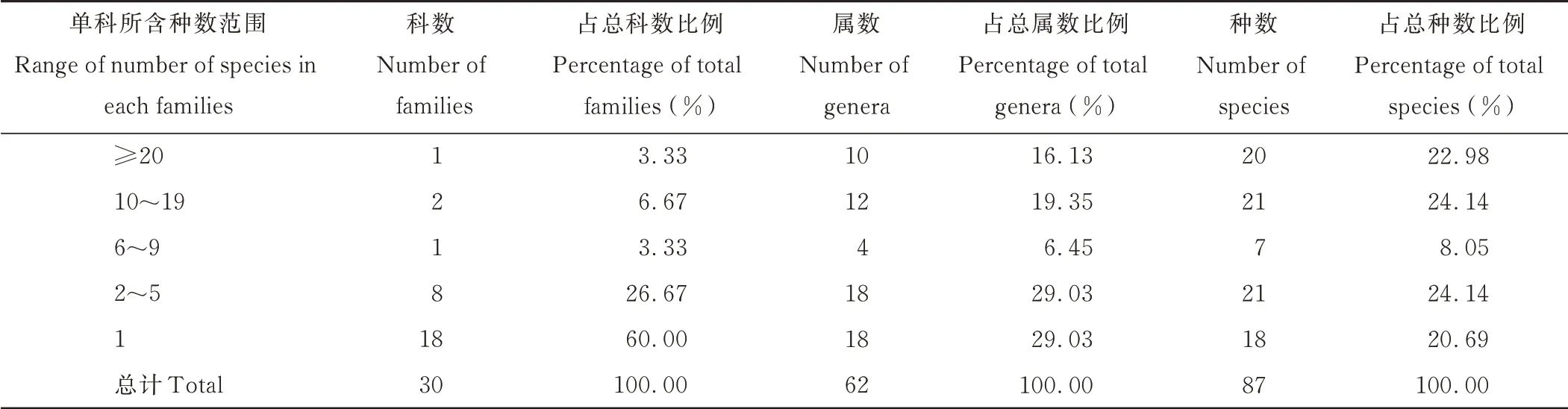
表2 賀蘭山巖羊主要活動區植物群落物種數量Table 2 Species number of plant community in main active areas of P.nayaur in Helan Mountains
在賀蘭山巖羊主要活動區植物群落的物種中,包括一年生草本、多年生草本、半灌木、灌木、喬木和木質藤本6 種生活型的植物。其中,草本63 種,一年生草本4 種,多年生草本59 種;木本23 種,灌木12 種,半灌木7 種,喬木4 種,藤本1 種,為木質藤本(圖1)。
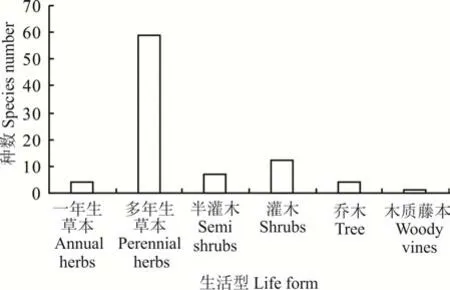
圖1 賀蘭山巖羊主要活動區植物生活型Fig. 1 Plant life forms of the main active areas of P. nayaur in Helan Mountains
2.2 賀蘭山巖羊主要活動區不同海拔的植物群落組成與特征
不同海拔植物群落組成各有差異。海拔在1100~1500 m 的研究區科數最少,與其他海拔研究區的科數具有顯著差異(P<0.05),而其他海拔研究區之間不存在顯著差異(P>0.05);在屬水平上,2500 m以上海拔的研究區屬數最多,和其他海拔研究區之間存在顯著差異(P<0.05),最少的是1100~1500 m 海拔處的研究區,與其他海拔研究區的屬數具有顯著差異(P<0.05);在種水平上,海拔在2500 m 以上的研究區植物種數最多,和其他海拔研究區之間存在顯著差異(P<0.05),最少的是1100~1500 m 海拔處的研究區,與其他海拔的種數具有顯著差異(P<0.05)(表3)。
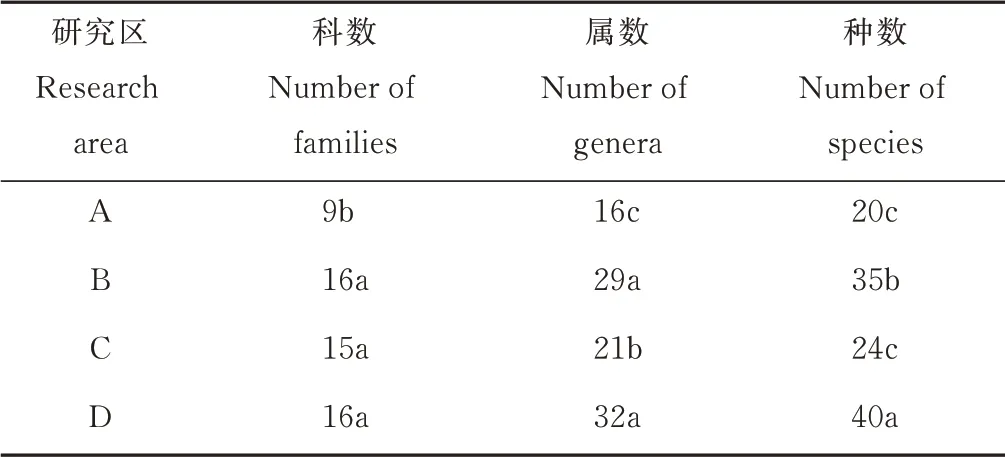
表3 賀蘭山巖羊主要活動區不同海拔植物所含科、屬、種的數量Table 3 Number of families,genera,and species of plants at different altitudes in main active areas of P. nayaur in Helan Mountains
在1100~1500 m 的研究區,主要植被類型為荒漠草原,共有9 科16 屬20 種植物,其中以禾本科為主,有7 種,其次為莧科、旋花科、豆科、菊科等,主要屬有針茅屬(Stipa)、旋花屬(Convolvulus)、隱子草屬(Cleistogenes)和堿豬毛菜屬(Salsola)等;海拔在1500~2000 m 的山地疏林草原,共有16 科29 屬35 種,其中禾本科最多,有8 種,其次為薔薇科(7 種)、菊科和豆科(各4 種),主要屬有針茅屬、早熟禾屬(Poa)、繡線菊屬(Spiraea)、桃屬(Amygdalus)等;海 拔 在2000~2500 m 的 山 地 針 葉林,共有15 科21 屬24 種植物,主要植物為禾本科(5種)、薔薇科(3 種)和菊科(3 種),主要屬為蒲公英屬(Taraxacum)、披 堿 草 屬(Elymus)和 鵝 觀 草 屬(Roegneria);海拔在2500 m 以上的高山?亞高山灌叢草甸中共有16 科32 屬40 種植物,主要科為禾本科(7 種)、菊科和薔薇科(各6 種)、石竹科(4 種)、豆科(3種)等,主要屬有委陵菜屬(Potentilla)、披堿草屬、早熟禾屬等(圖2)。
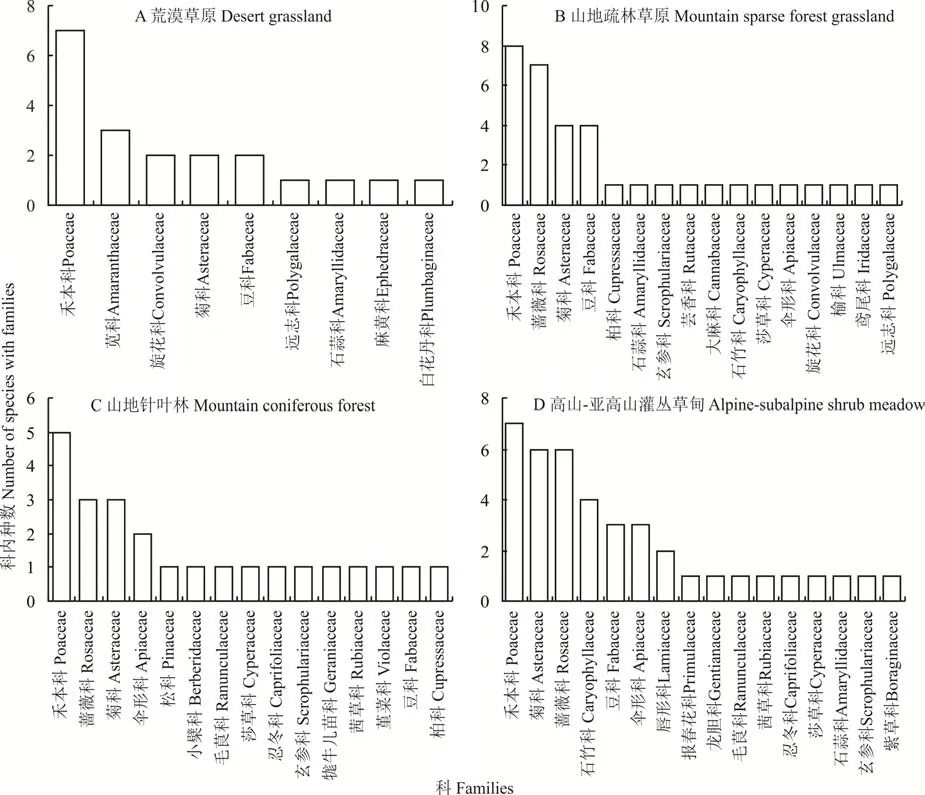
圖2 賀蘭山巖羊主要活動區不同海拔植物群落各科所含種數Fig. 2 Species number of each family in plant communities at different altitudes in main active areas of P. nayaur in Helan Mountains
2.3 不同海拔梯度植被優勢種及其重要值
賀蘭山巖羊主要活動區植被類型沿海拔由低到高依次為荒漠草原、山地疏林草原、山地針葉林和高山?亞高山灌叢草甸,巖羊主要活動區植物物種重要值排序結果顯示,賀蘭山巖羊主要活動區植物優勢種以禾本科草本植物和薔薇科灌木植物為主(表4)。
在草本層,低海拔荒漠草原的優勢種為短花針茅(Stipa breviflora)(重要值為0.332,下文括號中數值同為重要值)、無芒隱子草(Cleistogenes songorica)(0.200)等,中低海拔處的山地疏林草原優勢種為針茅(Stipa capillata)(0.327),中高海拔處的山地針葉林的優勢種為苔草(Carexspp.)(0.428)、真穗披堿草(Elymus gmelinii)(0.183)、小紅菊(Chrysanthemum chanetii)(0.126)等,高海拔的高山?亞高山灌叢草甸的優勢種為苔草(0.226)等(表4)。
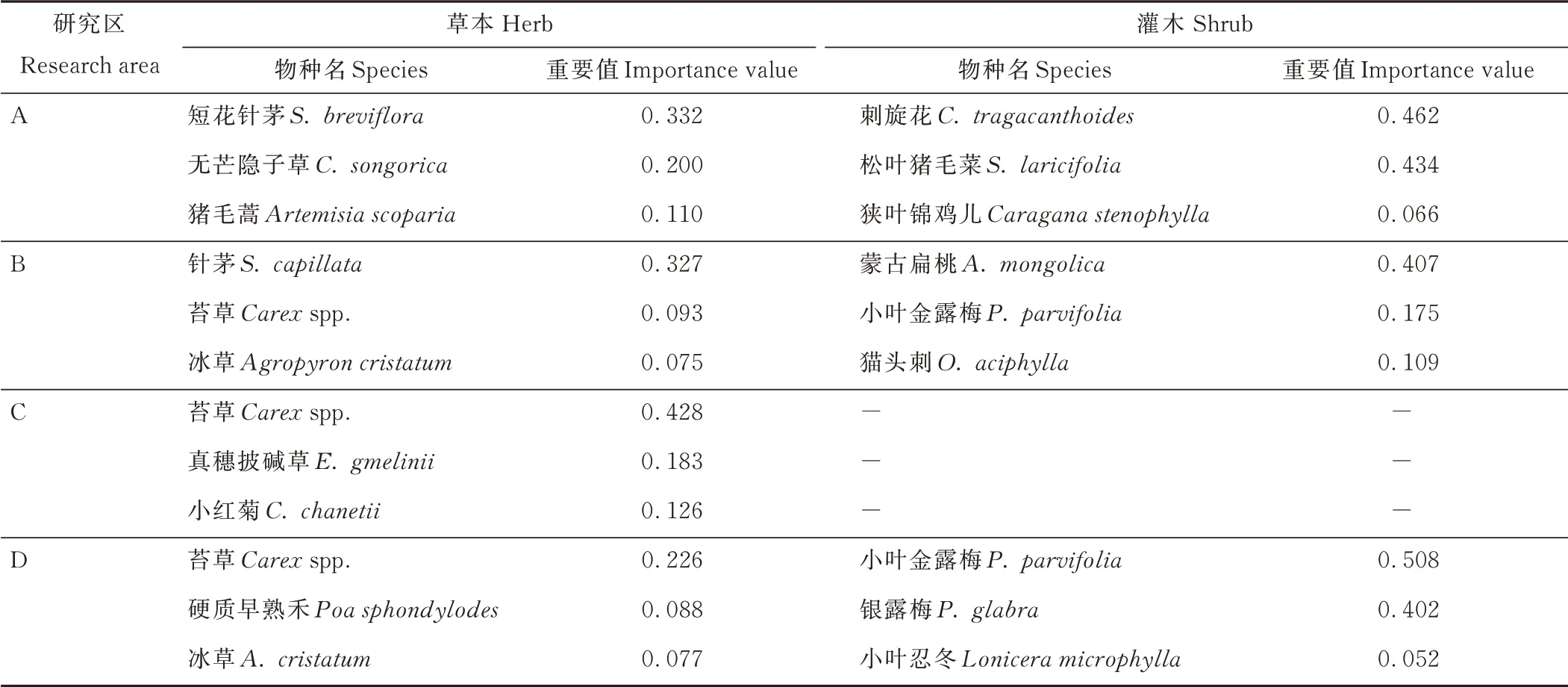
表4 賀蘭山巖羊主要活動區不同植物群落類型優勢種及其重要值Table 4 Dominant species and their importance values of different plant community types in main active areas of P. nayaur in Helan Mountains
在灌木層,低海拔荒漠草原的優勢種為刺旋花(Convolvulus tragacanthoides)(0.462)、松葉豬毛菜(Salsola laricifolia)(0.434)等,中低海拔山地疏林草原優勢種為蒙古扁桃(Amygdalus mongolica)(0.407)、小葉金露梅(Potentilla parvifolia)(0.175)和貓頭刺(Oxytropis aciphylla)(0.109)等,高海拔的高山?亞高山灌叢草甸的優勢種為小葉金露梅(0.508)、銀露梅(Potentilla glabra)(0.402)等,而中高海拔山地針葉林沒有灌木植物生長。
2.4 賀蘭山巖羊主要活動區不同海拔的植物物種多樣性變化
賀蘭山巖羊主要活動區植物群落多樣性方差分析和多重比較結果顯示,在群落水平上,草本層的物種豐富度、Shannon?Wiener 多樣性指數和Simpson 優勢度指數均大于灌木層,而均勻度指數小于灌木層。不同研究區之間的草本層植物的物種豐富度指數、Pielou 均勻度指數和Shannon?Wiener 多樣性指數都存在顯著差異(P<0.05),而Simpson 優勢度指數之間沒有顯著差異(P>0.05);不同研究區之間灌木層的豐富度指數、Shannon?Wiener 多樣性指數和Simpson 優勢度指數都存在顯著差異(P<0.05),而Pielou 均勻度指數沒有顯著差異(P>0.05)(表5)。
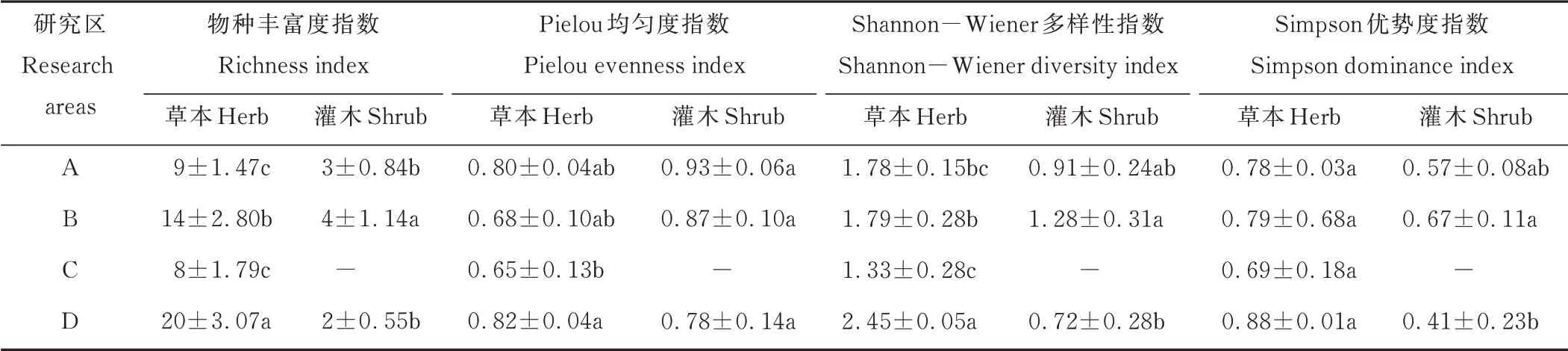
表5 賀蘭山巖羊主要活動區植物群落多樣性對海拔梯度的響應規律Table 5 Response of vegetation community diversity to altitudinal gradient in main active areas of P.nayaur in Helan Mountains
回歸分析結果顯示,草本層植物的物種豐富度隨著海拔升高而升高;Pielou 均勻度指數、Shannon?Wiener 多樣性指數、Simpson 優勢度指數均與海拔高度相關不顯著,呈現出先降低再升高的特征。灌木植物的物種豐富度隨海拔升高而降低;Pielou 均勻度指數與海拔高度之間呈顯著相關,隨海拔升高而降低;Shannon?Wiener 多樣性指數和Simpson 優勢度指數均與海拔呈顯著相關,隨海拔的增加先升高后降低(圖3)。
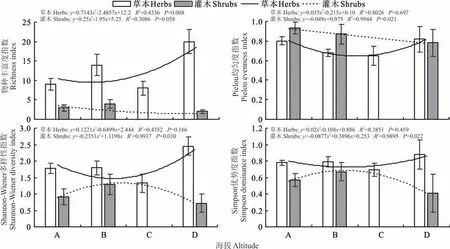
圖3 植物物種多樣性與海拔高度的關系Fig.3 Relationship between plant species diversity and altitude
草本層植物的物種豐富度、Pielou 均勻度指數、Shannon?Wiener 多樣性指數和Simpson 優勢度指數最高的均為海拔在2500 m 以上的高山?亞高山灌叢草甸,指數最低的均為海拔在2000~2500 m 的山地針葉林,不同海拔處的研究區草本植物物種多樣性指數由高到低依次是高海拔區、中低海拔區、低海拔區和中高海拔區。
灌木層植物的物種豐富度、Shannon?Wiener 多樣性指數和Simpson 優勢度指數最高的均為海拔位于1500~2000 m 的山地疏林草原,而Pielou 均勻度指數最高的為位于低海拔處的荒漠草原;各個指數最低的均為2500 m 海拔以上的高山?亞高山灌叢草甸。不同海拔處的研究區灌木植物物種多樣性指數由高到低依次是中低海拔區、低海拔區和高海拔區。
3 討論
3.1 賀蘭山巖羊主要活動區植物區系特征及其與巖羊等食草動物的關系
賀蘭山位于蒙古高原南緣、黃土高原西北處,又與青藏高原東北部鄰近,因此這里匯聚了蒙古高原、華北平原、青藏高原的植物成分,使得賀蘭山的植物地理成分復雜,而具有明顯的過渡性特征[23?24],又因其高度差較大、水熱分布差異,從而形成了明顯的垂直地帶性植被[25]。根據相關研究,賀蘭山西麓的植物物種以禾本科居多,菊科、豆科、薔薇科等次之[26]。而本研究在賀蘭山巖羊主要活動區植被調查共記錄到87 種植物,隸屬于30 科62 屬,禾本科包含的植物種類最多,其次是薔薇科、菊科和豆科等。巖羊主要活動區植物生活型組成有灌木、半灌木、多年生草本、一年生草本、木質藤本,這與胡天華[23]和段河等[26]在寧夏賀蘭山國家級自然保護區的研究結果相符。
在賀蘭山,荒漠草原以短花針茅為優勢種,群落內植物多在10 種左右[25];山地疏林草原主要是灰榆、蒙古扁桃、短花針茅等小喬木和灌木稀疏分布[27];山地針葉林主要生長著油松(Pinus tabuliformis)和青海云杉(Picea crassifolia);海拔在2500 m 以上的高山?亞高山灌叢草甸植物種類繁多,草本植物主要有苔草、早熟禾及其他雜類草[26]。本研究關于巖羊主要活動區的植物區系特征印證了這些結果。
賀蘭山巖羊所采食的植物以禾本科植物為主[28],還包括薔薇科,豆科、菊科等植物,其中針茅是巖羊的大宗食物,早熟禾、灰榆、冰草等植物也是巖羊的主要采食植物[29]。本研究植物區系特征結果與巖羊主要采食的植物相符合,說明賀蘭山巖羊在賀蘭山能夠獲得生長發育所需的食物,同時,植物組成也可能是巖羊選擇其活動區的關鍵因素。
有研究表明,即使食草動物的數量很少,也會是保護和恢復目標的障礙,因為它們對一些植物物種有著持續壓力[30]。在賀蘭山,巖羊數量持續增長,且分布并不均勻[31],其生境已經呈破碎化狀態,部分生境內的植被因為過度啃食已經出現嚴重退化的現象,已經阻礙了植被的生長和恢復,對賀蘭山植被的保護、生物多樣性和群落結構的恢復及協調發展造成了巨大的威脅。
因此,植物群落組成及多樣性與賀蘭山巖羊等食草動物之間有著深刻的聯系。食草動物對生態系統功能具有至關重要的作用,一些關鍵的生態系統過程受到它們的影響,如養分的周轉和種子的傳播[32?33]。而且它們還調控著植物的多樣性和生產力[34],食草動物的捕食壓力下,植被幼苗死亡率高,且其高度的生長率甚至可以忽略不計,從而導致植物成活率極低,而食草動物數量的減少會促使大多數植被的幼苗增長[35]。食草動物對植物群落的多樣性和組成有著強烈的影響[36]。植物物種多樣性會隨著食草動物采食強度的增強而降低,而且在食草動物的采食行為中具有較強的選擇性,這種行為則可能會加劇植物物種多樣性的降低[37]。
同時,植物多樣性對食草動物具有積極的調節作用[38]。植物多樣性的增加通常會導致植物生物量和棲息地多樣性的增加,這兩者都有利于食草動物。植物多樣性的增加可以通過提供更廣的范圍和更豐富的食物來促進食草動物數量的增加。食草動物對植物多樣性的響應則反映了棲息地和營養效應之間的平衡狀態[39]。
3.2 賀蘭山不同海拔的巖羊主要活動區植物物種多樣性的影響因素
能夠對物種多樣性產生影響的因素多種多樣,如降水、溫度、濕度、陽光、土壤條件、海拔和人為干擾等。本研究結果顯示,海拔在2500 m 以上的高山?亞高山灌叢草甸中的草本層植物的物種豐富度和Pielou 均勻度指數都高于其他研究區,而2000~2500 m 海拔處的山地針葉林研究區的草本植物物種豐富度和Pielou 均勻度指數最低。這也與巖羊在高山草原活動較頻繁[40]相印證。造成這種結果最主要的因素是水分、溫度等氣候因素[41?42],而海拔高度能夠對水、熱、濕度等條件產生直接決定作用[43]。寧夏賀蘭山處于中溫帶干旱地區,這一氣候區域使得熱量和水分條件成了植物生長的限制因子,氣候格局從而呈現出氣溫隨海拔升高而降低,降水隨海拔升高而增多[44]。在中低海拔區,相對來說溫度高而降水量少,因此其蒸散量和干旱程度高于高海拔地區,從而這一區域的植物生長受到水分的限制作用,而在高海拔地區降水量相對增多,蒸散量相對減少,水分不再是限制植物生長的主導因素,加之人為干擾減少,因此物種豐富度逐漸增加[45]。高山?亞高山灌叢草甸研究區海拔位于2500 m 以上,降水量多,受到太陽直射時間較長,光照使得溫度對植物的限制相對降低,因此這一研究區的草本植物物種多樣性最高。山地針葉林研究區海拔處于2000~2500 m,降水量多,但這一研究區因高大喬木郁閉度較高,導致陽光不能直射至林下,空氣濕度大,氣溫低,這可能是導致這一研究區草本層植物物種豐富度最低的原因,而該研究區沒有灌木植物生長也與這一氣候條件限制有關,灌木的生長喜光且不耐澇,這一地區林下既沒有陽光直射,濕度又大,不利于灌木的生長。相反在中低海拔區,由于灰榆等小喬木生長稀疏,冠層較小,使太陽光能夠照射至灌木層,又因這一區域溫度較高導致蒸散量和干旱程度高,降水量相對較少,符合灌木的生長習性,因此在中低海拔區的灌木層植物的物種豐富度和Pielou 均勻度指數相對較高,這也與Jiang 等[4]的研究結果一致。但灌木也存在死亡率較高的現象[46],這是由于當種間和種內對于光照、水分、養分和空間等因素的競爭激化時,灌木的生長受到抑制所致。
已有研究表明,草本植物的Shannon?Wiener 多樣性指數隨海拔升高而增加[47]。本研究的結果與上述規律基本一致,除了山地針葉林研究區,其他研究區的Shannon?Wiener 多樣性指數沿海拔梯度的變化印證了這一結果。其中海拔2500 m 以上的高山?亞高山灌叢草甸研究區與其他研究區之間的多樣性指數存在顯著差異,產生這種差異的原因主要是不同生境之間的差異使得各研究區生長的物種產生差別,進而使植物群落在結構和功能方面都產生了差異[48?49]。本研究通過對各研究區之間的Shannon?Wiener 多樣性指數進行比較,發現山地針葉林研究區的Shannon?Wiener 多樣性指數最低,Jiang 等[4]研究發現賀蘭山以青海云杉(Picea crassifolia)為主的山地針葉林Shannon?Wiener 多樣性指數最低,Liu 等[50]發現巖羊對山地針葉林植物的食用量低,而在本研究中的Shannon?Wiener 多樣性指數最低的同樣是以青海云杉為主的山地針葉林,這也說明了本研究與前人的結果一致。
Simpson 優勢度指數和植物群落中的物種數量有關,優勢度越大,植物群落不同種類的植物數量分布越不均勻。在本研究中,草本層植物的Simpson 優勢度指數在0.69~0.88 之間,灌木層的優勢度指數在0.34~0.67 之間,灌木層和草本層之間優勢度的差異,可能是由于在巖羊主要活動區內灌木植物之間的種內和種間競爭激烈,從而導致灌木植物物種數量較低,進而使得灌木層優勢度指數低于草本層。灌木在海拔2500 m 以上的高山?亞高山灌叢草甸研究區的優勢度指數最小,可能是由于該研究區海拔高,人類活動少,但風力大,風蝕嚴重,不利于灌木的生長發育[16]。
物種多樣性受到多因素的調節,而植物也能根據環境等因素有形態適應、物候適應、生理生態適應的多樣性[10]。賀蘭山的植物種類豐富、區系地理復雜、沿海拔垂直帶變化多樣共同造成了其物種多樣性。
4 結論
根據植被調查結果,構成巖羊主要活動區的草本植物主要為禾本科植物,灌木以薔薇科植物為主。海拔梯度對賀蘭山巖羊主要活動區的植物物種多樣性變化有一定影響,草本層植物物種多樣性在高海拔地區達到峰值,而灌木植物由于其自身喜光但不耐澇的特性,在中海拔地區物種多樣性最大。整體來看,草本層植物物種多樣性隨著海拔的升高在巖羊主要活動區呈現出先降低后升高的趨勢,而灌木層植物物種豐富度、Pielou 均勻度指數呈現出沿海拔上升而降低的特征,Shannon?Wiener 多樣性指數和Simpson 優勢度指數呈現出先增加后降低的特征。因此,建議在不同海拔對賀蘭山巖羊采取不同的管護措施,以達到防止植被退化和促進巖羊種群健康發展的效果。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
