肝細胞癌的細胞分裂周期基因20表達與臨床病理特征關系及對預后的影響
2022-06-11 07:42:34宋應周孟松峰
安徽醫藥 2022年6期
關鍵詞:研究
宋應周,孟松峰
肝細胞癌在全球肝臟惡性腫瘤中發病率呈上升趨勢,已成為癌癥死亡的第二大誘因[1]。乙型肝炎病毒或丙型肝炎病毒慢性感染、肝硬化、酒精性肝病和黃曲霉毒素等造成了肝細胞癌的高發病率[2-4]。然而,促進肝癌發病機制仍不清楚。在肝細胞癌發生過程中,細胞周期檢查點的改變可能與癌癥形成密切相關[5]。細胞分裂周期基因20(CDC20)是細胞周期檢查點的調節劑,與細胞周期后期中復合物/環體相互作用的調節蛋白和有絲分裂相關[6]。CDC20可與APC結合并在有絲分裂和G1期激活細胞周期蛋白的泛素化活性,促進細胞增殖[7]。CDC20可通過調節細胞周期、促進細胞凋亡,抑制惡性轉化[8]。研究表明,多種癌癥中CDC20的高表達,與腫瘤的增殖、侵襲及不良預后有關[9-12]。然而,CDC20在肝細胞癌中的研究較少,該研究通過檢測肝細胞癌組織中CDC20的水平,分析CDC20的表達與病人臨床病理特征的關系及對預后的影響,為肝細胞癌的預后判斷提供參考。
1 資料與方法
1.1一般資料收集2016年5月至2017年12月在商丘市第三人民醫院行手術治療的肝細胞癌病人146例,男115例,女31例,年齡范圍為38~62歲,收集病人的臨床資料。收集手術切除的肝細胞癌組織及對應的癌旁組織,放置于?80℃超低溫冰箱中。術后采用電話的方式定期隨訪,最短隨訪時間5個月,最長隨訪時間36個月,隨訪截止日期為2020年12月1日,146例的臨床資料及隨訪資料完整。該研究經商丘市第三人民醫院倫理委員會批準(20160412),病人或其近親屬同意并簽署知情同意書。
1.2研究方法
1.2.1實時熒光定量逆轉錄聚合酶鏈反應(qRTPCR) qRT-PCR檢測肝細胞癌組織中CDC20mRNA的表達。組織中的總RNA根據Trizol試劑盒進行提取。提取的RNA逆轉錄為互補DNA(cD‐NA),進行實時熒光定量PCR。CDC20正向引物序列5′-TCGAAGTCACTCGGTAGCGT-3′,反向引物序列5′-GATCCTGCATACTGCTTGCA-3′;U6正向引物序 列5′-GGCCTCATCCTCATCATGGCAGUAG-3′,反向引物序列5′-AGCCAGATGCCTAGCGACGTGC‐GC-3′。總反應體系為20μL,反應條件:95℃預變性5 min,95℃變性30 s,60℃退火30 s,72℃延伸30 s,40個循環。以管家基因U6 mRNA水平作為內參照,采用2?ΔΔCt法計算CDC20 mRNA的相對表達水平。
1.2.2蛋白質印跡法 蛋白質印跡法檢測肝細胞癌組織中CDC20的表達。取組織中的總蛋白,根據BCA蛋白定量試劑盒取30μg的蛋白,行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE),將電泳凝膠上的蛋白濕轉至聚偏二氟乙烯(PVDF)膜上,PVDF膜放置含有1%BSA的盒子中,孵育2 h、之后經過一抗孵育(1∶1 000稀釋,4℃過夜)、TBST洗滌3次、二抗孵育(1∶4 000稀釋,室溫1 h)、TBST洗滌3次后,在PVDF膜上滴加適量化學發光顯色液,在凝膠成像系統中拍照,根據灰度值估測CDC20蛋白表達水平。
1.2.3免疫組織化學實驗 免疫組織化學檢測肝細胞癌組織中CDC20的表達。切制冷凍切片,經過脫蠟和梯度乙醇水化處理后,利用PV6000法染色,加入3%過氧化氫滅活內源性過氧化物酶。置于修復液中,加熱行抗原進行修復,然后放置室溫自然冷卻。經封閉后,滴加一抗(1∶200),置于4℃冰箱孵育過夜。利用磷酸緩沖鹽溶液(PBS)沖洗,滴加相應種屬二抗(1∶400),室溫下孵育60 min,并用PBS進行3遍沖洗。滴加新鮮配制的顯色液,利用蘇木素染料復染,中性樹膠封固,最后置于顯微鏡下觀察。染色強度評分:強陽性、中等陽性、弱陽性和陰性評分分別為3分、2分、1分和0分,陽性細胞數的評分:>75%~100%、>50%~75%、>25%~50%、5%~25%和<5%評分分別為4分、3分、2分、1分和0分,染色強度評分和陽性細胞數的評分的乘積即為總分,0~7分為低水平,8~12分為高水平。
1.3統計學方法應用SPSS 21.0軟件行統計分析。計量資料以±s表示,組間比較采用配對t檢驗;計數資料以例(%)表示,組間比較采用χ2檢驗,多組間的兩兩比較檢驗水準α=0.016;采用Kaplan-Meier法繪制疾病無進展生存率和總生存率曲線,log-rank檢驗比較不同組間的生存率。以P<0.05為差異有統計學意義。
2 結果
2.1qRT-PCR檢測肝細胞癌組織中CDC20mRNA的表達CDC20 mRNA在肝細胞癌組織表達水平(2.33±0.67)高于對應的癌旁組織(0.97±0.29)(t=5.95,P<0.001)。
2.2蛋白質印跡法檢測肝細胞癌組織中CDC20的表達CDC20蛋白在肝細胞癌組織中相對表達水平(0.91±0.05)高于對應的癌旁組織(0.14±0.05)(t=34.30,P<0.001)。
2.3免疫組織化學檢測肝細胞癌組織中CDC20的表達CDC20在肝細胞癌陽性表達率(77.4%,113/146)高于對應癌旁組織(8.2%,12/146),差異有統計學意義(χ2=142.69,P<0.001)。見圖1(僅展示3例病人的結果)。
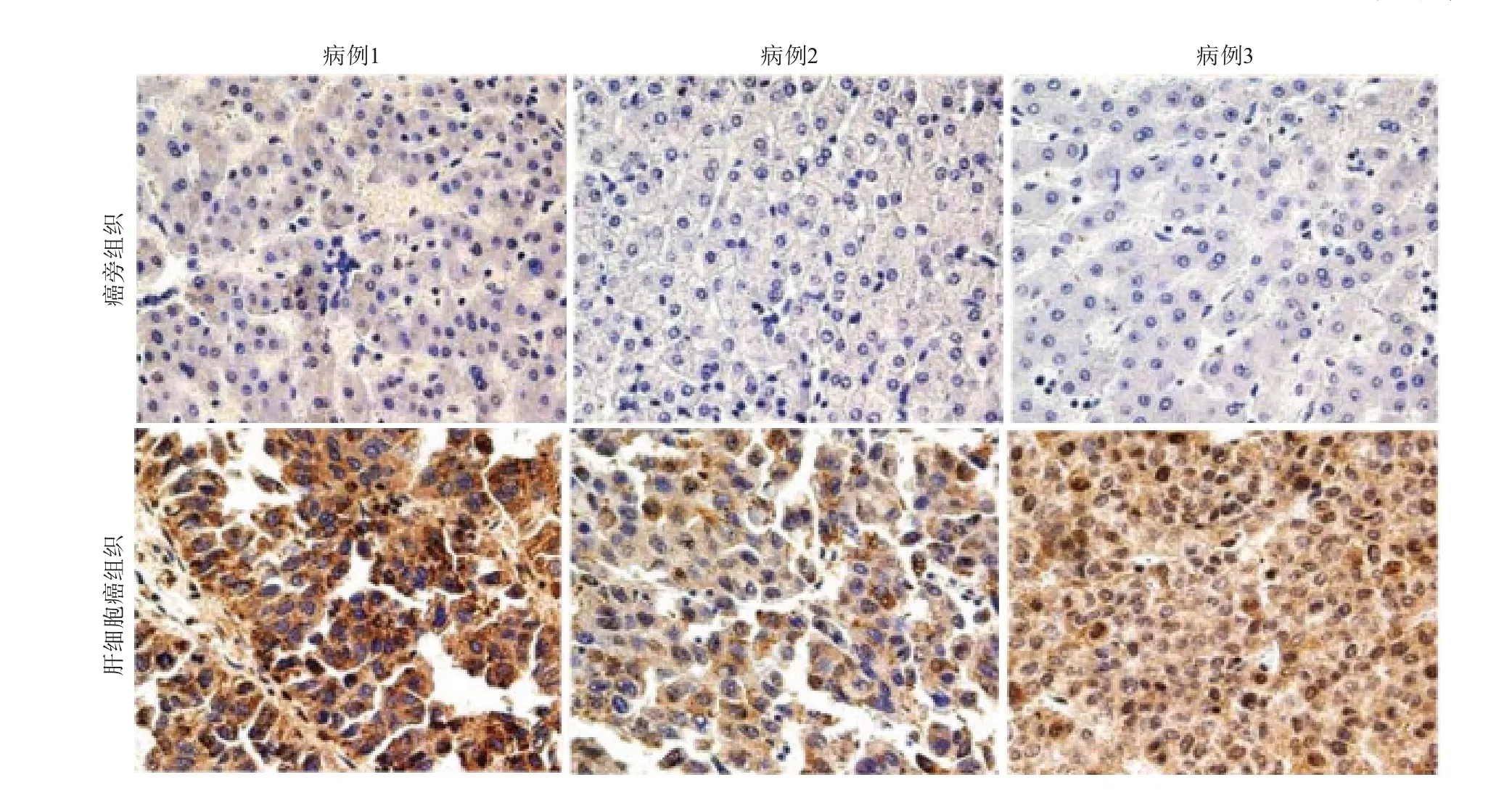
圖1 細胞分裂周期基因 20(CDC20)在3例肝細胞癌組織中的表達情況(免疫組織化學×200)
2.4CDC20表達與病人臨床病理特征的關系CDC20的表達與病人的腫瘤長徑、被膜侵犯、脈管侵犯、腫瘤多發、分化程度和TNM分期有關(P<0.05),見表1。
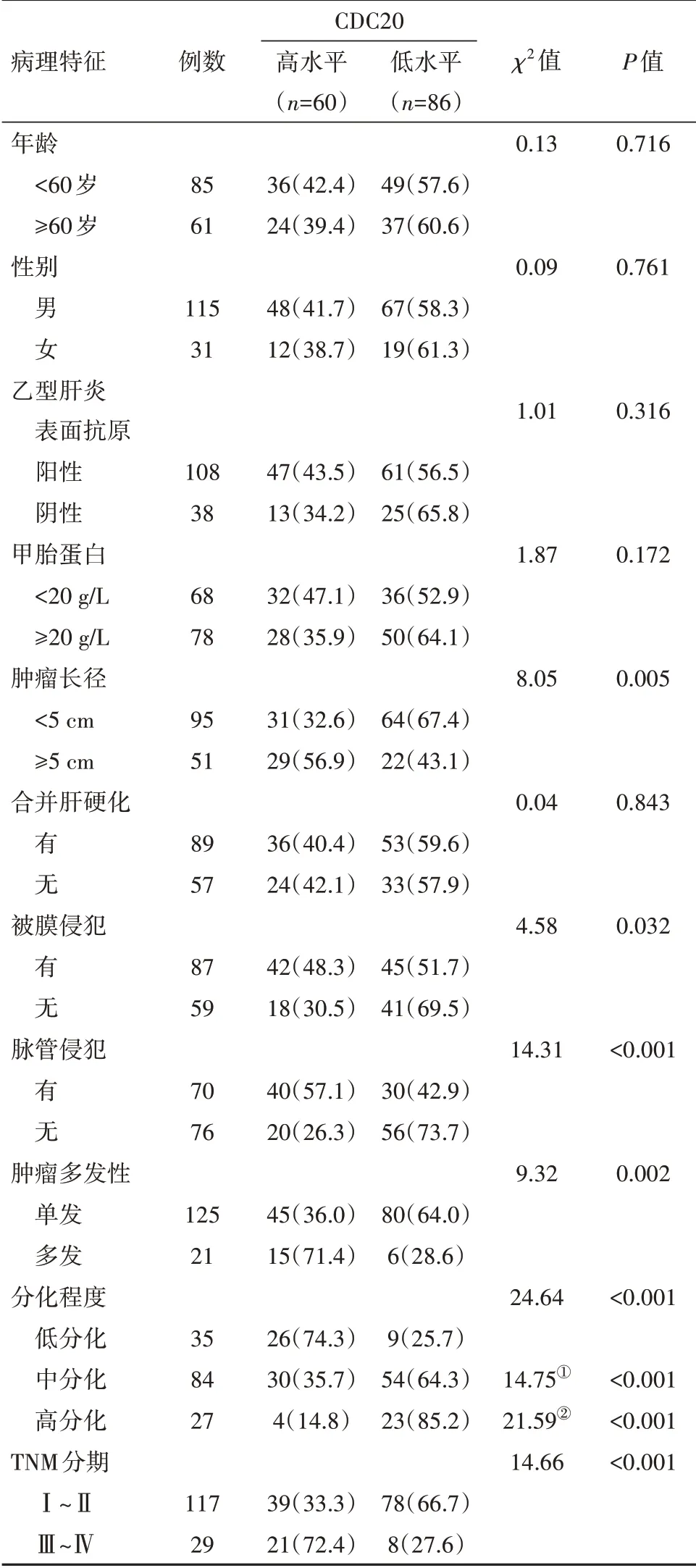
表1 細胞分裂周期基因20(CDC20)表達水平與肝細胞癌病人臨床病理特征的關系/例(%)
2.5CDC20表達對病人術后生存率的影響CDC20高水平組總生存率為(61.7%)和疾病無進展生存率(50.0%)均低于CDC20低水平組(87.2%、69.8%)(log-rankχ2=14.19,P<0.001;log-rankχ2=9.61,P=0.005)。
3 討論
肝細胞癌是常見的惡性腫瘤,在過去的20年中,觀察到與肝細胞癌相關的年死亡率顯著增加[13-14]。此外,根據美國國立癌癥研究所“監測、流行病學和結果數據庫”SEER注冊機構項目研究,到2030年肝細胞癌的發病率將繼續上升[15]。準確估計預后,在肝細胞癌治療中起著至關重要的作用。尋找預測肝細胞癌預后的生物標志物具有重要的臨床價值。許多CDC20底物都參與有絲分裂過程,包括細胞周期蛋白B1、細胞周期蛋白A、分離酶抑制蛋白(Securin)、雙能蛋白(Geminin)和周期素依賴激酶抑制劑p21(P21)[16]。最近,越來越多的研究顯示CDC20在人類癌癥中起致癌作用,在多種人類腫瘤中觀察到CDC20的過度表達,與生存率降低有關[17]。
CDC20在結直腸癌、乳腺癌、前列腺癌和胃癌中的表達均增加[9-12],在該研究中,肝細胞癌組織中CDC20 mRNA和蛋白質水平均高于癌旁組織,提示CDC20促進肝細胞癌的發生發展。目前多項研究顯示,CDC20高水平參與多種腫瘤的進展,例如Kim等[12]研究顯示,CDC20在腸化生、低度不典型增生、高度不典型增生和早期胃癌中的表達明顯高于正常黏膜,且在高度不典型增生中的表達水平最高,且CDC20過表達與胃癌病人的年齡、腸道組織學、較低的腫瘤-淋巴結-轉移階段以及較長的無復發生存期和癌癥特異性生存期相關。CDC20與前列腺癌的惡性進展相關,CDC20高水平病人更具侵略性的臨床病理特征和不良預后[11]。Li等[18]證明了膀胱尿路上皮癌病例中CDC20升高,而CDC20高水平與病人的無復發生存期較短和總體生存期較差相關。該研究中,CDC20在肝細胞癌中的表達水平與腫瘤長徑、被膜侵犯、脈管侵犯、腫瘤多發、分化程度和TNM分期有關,說明在腫瘤長徑、被膜侵犯、脈管侵犯、腫瘤多發分化程度低和TNM晚期的病人中CDC20的表達水平更高。提示CDC20可能與肝細胞癌的增殖與侵襲有關,CDC20高水平的肝細胞癌病人具有較強的增殖活性和更強的侵襲能力,與肝細胞癌的發展密切相關。在該研究中,CDC20高水平病人的術后3年總生存率和疾病無進展生存率均較低,提示CDC20有可能成為評價肝細胞癌預后的有效標志物。CDC20促進肝細胞癌的進展,其失活可能對治療肝細胞癌有一定療效。目前已發現幾種CDC20抑制劑,例如類毒素-抗毒素混合物酯酶、pro-TAME和2-[芐基-(2-硝基-苯磺酰基)-氨基]-N-羥基-3-甲基-N-丙基-丁酰胺[19-20]。對CDC20抑制劑的研究可能有助于闡明CDC20在人類癌癥中的作用機制,這也是后期該研究的方向之一。
綜上所述,CDC20在肝細胞癌中的表達水平增加,可能參與腫瘤的增殖和侵襲。CDC20高水平預示病人的預后不良,有望成為判斷肝細胞癌預后的腫瘤標志物。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
