植物對鎘吸收、轉運及耐性調控機制研究進展
2022-06-22 02:58:14李慧君明荔莉張文生
生態毒理學報 2022年2期
李慧君, 明荔莉, 張文生,*
1. 北京師范大學天然藥物教育部工程研究中心,北京 100088
2. 拱北海關技術中心,珠海 519000
3. 地表過程與資源生態國家重點實驗室珠海基地,珠海 519087
4. 北京師范大學中藥資源保護與利用北京市重點實驗室,北京 100875
鎘(cadmium, Cd)是自然界中普遍存在的一種重金屬元素,具有較高的生物毒性。 自然狀態下,土壤中Cd 含量處于較低水平。 然而,隨著工農業的發展,我國遭受Cd 污染的土地面積不斷增加,農田土壤Cd 污染尤為突出[1]。 采礦、冶煉、污水灌溉、污泥培肥和磷肥施用等人類活動以及大氣沉降是農田土壤Cd 污染的主要來源。 2014年,我國原環境保護部和原國土資源部公布的《全國土壤污染狀況調查公報》數據顯示,我國耕地土壤重金屬污染點位超標率達19.4%,其中Cd 的點位超標率為7.0%,污染最為嚴重[2]。 與其他重金屬相比,土壤中的Cd 遷移性較強、生物毒性較高,容易被作物吸收、富集,進入食物鏈,最終危害人體健康[3-4]。 因此,Cd 污染受到人們的廣泛關注。
Cd 是植物生長發育過程中的非必需金屬元素。研究表明,Cd 吸收會導致細胞損傷,干擾細胞正常生理代謝過程,影響植物生長發育[5]。 為抵御Cd 毒害,植物進化出了多種解毒機制,主要包括限制根系Cd 吸收、阻止Cd 進入細胞、螯合作用、液泡儲存和激活抗氧化防御系統等[6]。 在不斷認識植物對Cd毒性生理調控的同時,人們開始探索其背后的分子機制。 本文圍繞植物對Cd 吸收、轉運的吸收蛋白和排出蛋白,以及調控Cd 耐性基因表達的轉錄因子和microRNA 這4個方面總結相關研究進展,以期為今后相關研究提供參考。
1 植物對鎘的吸收、轉運(Uptake, translocation of cadmium in plants)
1.1 鎘吸收蛋白
重金屬吸收蛋白是Cd 進入根細胞的主要轉運蛋白,一般定位在細胞質膜上,負責將重金屬轉運至細胞質,包括鋅/鐵蛋白(ZIP)家族、天然抗性巨噬細胞蛋白(NRAMP)家族和黃色條紋蛋白(YSL)家族等。
ZIP 家族是鋅轉運蛋白(ZRT)和鐵轉運蛋白(IRT)家族的總稱,優先參與鋅(Zn)和鐵(Fe)的吸收、轉運,同時也參與根系對Cd 的吸收過程。 擬南芥中的AtIRT1是從植物中克隆出的第一個ZIP家族基因,它是缺Fe 情況下根部誘導的Fe2+高親和吸收蛋白[7]。 目前,已在擬南芥、水稻、苜蓿和大豆等植物中鑒定出了100 多個ZIP家族基因,該家族成員具有廣泛的底物特異性,參與Cd2+、Zn2+和Cu2+等多種金屬離子的吸收轉運[8]。 Zheng 等[9]發現 Cd 脅迫下,AtIRT1和OsZIP1基因表達上調,證明了AtIRT1和OsZIP1分別參與擬南芥和水稻根系對Cd2+的吸收。 在缺鐵的情況下,水稻中OsIRT1和OsIRT2會促進根部吸收Cd,且OsIRT1較OsIRT2對Cd 表現出更高的敏感性[10]。 Zn/Cd 超積累植物東南景天能通過位于細胞質膜的 SaZIP4 蛋白超吸收 Zn 和Cd[11]。 最近,Tan 等[12]發現,OsZIP5和OsZIP9協同作用,參與水稻對Zn 和Cd 的吸收。 研究表明,Fe和Zn 均與Cd 的吸收存在一定競爭關系,即在Cd脅迫下,植物將減少對Zn/Fe 的吸收[13]。 大多數ZIP蛋白的表達是由Zn、Fe 或Mn 元素缺乏誘導的,調控ZIP 家族蛋白的內源表達對維持細胞金屬穩態至關重要,主要是由于該家族基因參與了植物根系對金屬的吸收及金屬在植物不同組織部位的運輸和分配[14]。
NRAMP 家族是一個膜整合蛋白家族,最初被認為只能轉運Fe2+,之后的研究發現其對Cd2+、Zn2+和Mn2+等離子轉運也具有重要的調控作用[8]。 如At-NRAMP3 和AtNRAMP4 位于擬南芥液泡膜上,參與Fe2+、Mn2+和 Cd2+從液泡向細胞質的轉運[15-17]。 水稻中OsNRAMP1和OsNRAMP5是同源質膜基因,主要在根部表達,具有向根細胞轉運Cd 和Mn 的能力[18-19]。 其中,OsNRAMP5 是水稻中主要的 Cd 吸收蛋白,幾乎承擔了90%的Cd 吸收轉運[20]。 Tang等[21]通過敲除OsNRAMP5得到的突變體水稻較普通野生型水稻根和芽中Cd 含量明顯降低。 Chang等[22]發現OsNRAMP5過表達可提高水稻根系中Cd的積累,但是莖和籽粒中Cd 積累顯著降低,研究者認為主要是因為Cd 在根系的徑向運輸受到干擾,從而影響了其在木質部的裝載。 此外,Zhang 等[23]在東南景天中發現了一種新的參與Cd 和Zn 超積累的質膜轉運蛋白——SaNRAMP1,煙草過表達SaNRAMP1能顯著增加體內 Cd、Zn 和 Mn 的積累。 李亞敏等[24]根據模式生物Nramp基因的保守序列,克隆測序得到了玉米Nramp1基因,Cd 脅迫實驗發現玉米葉片和根系中ZmNramp1基因的表達量與Cd 含量呈現顯著正相關關系,證實了該基因與玉米中重金屬Cd 的吸收轉運具有密切聯系。
YSL 蛋白是寡肽轉運蛋白(OPT)家族的一員,主要參與植物中金屬與植物載體(PS)或煙堿胺(NA)螯合物的吸收轉運。 玉米ZmYS1 是第一個被鑒定出的YSL 家族蛋白,作為載體參與Fe(Ⅲ)-PS 的跨膜吸收和運輸[25],同時在不同植物體內、不同組織部位介導細胞對 Cd2+、Fe2+和Zn2+的吸收轉運。 Feng等[26]在超富集植物龍葵中克隆得到了SnYSL3基因,發現 Cd 脅迫會刺激SnYSL3表達上調,將SnYSL3在酵母中表達可以促進 Cd2+/Fe2+/Cu2+/Zn2+-NA 復合物的轉運,首次報道了 SnYSL3 參與Cd-NA 復合物的轉運。 Wang 等[27]將從印度芥菜中分離得到的BjYSL7在煙草中過表達后發現,莖中Cd 含量顯著高于野生型植株,這說明BjYSL7可能介導了Cd 向地上部分的運輸。 紫花苜蓿MsYSL1定位于質膜,過量的 Mn、Cd、Pb 或缺 Fe、Zn、Cu 會刺激其在根中表達,表明MsYSL1 可能參與包括Cd在內的多種重金屬-NA 復合物的轉運[28]。
1.2 鎘排出蛋白
排出蛋白的功能是將重金屬排出細胞質,或運載至液泡,是植物根細胞中Cd 進入木質部或區隔在液泡中的主要轉運蛋白。 重金屬ATP 酶(HMA)和Ca2+/H+反向轉運蛋白(CAX)是植物體內重要的Cd 排出蛋白。
HMA 是一類能夠利用ATP 水解產生的能量驅動離子進行跨膜運輸的轉運蛋白,在Cd 向地上部運輸和液泡區室化中發揮重要作用。HMA家族基因在擬南芥和水稻中的研究相對成熟。 Wong 和Cobbett[29]研究發現,敲除AtHMA2和AtHMA4基因后,Cd 在擬南芥根莖間的轉移幾乎完全消失,證明了定位于質膜的HMA2 和HMA4 是負責擬南芥根莖間Cd 轉運的主要因素。 Liedschulte 等[30]在煙草中也有類似的發現:敲除NtHMA4基因會導致根部Cd 向莖的轉移減少90%以上。 水稻中OsHMA2基因主要在根部維管束薄壁細胞中表達,與擬南芥的AtHMA2和AtHMA4功能類似,參與 Cd 在木質部的裝載[31]。 而HMA3 位于液泡膜上,在植物的不同組織部位廣泛分布,具有區隔化Cd 和Zn 的功能。Ueno 等[32]首次報道了水稻OsHMA3基因位于根部細胞液泡膜上,可選擇性地將Cd 區隔在液泡中,限制Cd 向地上部的轉運。 而Shao 等[33]則在控制Os-HMA2表達的前提下發現,過表達OsHMA3會促進水稻各組織部位Cd 向液泡轉運,從而降低水稻籽粒中的Cd 含量。 最近,吳海濤等[34]從野生重金屬耐性植物伴礦景天中克隆到一個HMA家族的基因——SpHMA2,該基因高表達可提高模式植物擬南芥的Cd 耐性及地上部分積累,說明SpHMA2是一個對Cd 正響應的基因,且在Cd 脅迫時,可能主要負責Cd 從地下往地上部的轉運。
CAX 是一類定位在細胞質膜、液泡膜、葉綠體類囊體膜及線粒體膜上的轉運蛋白,能夠與HMA蛋白家族協同作用參與細胞質中二價陽離子向液泡的轉移積累[35]。 植物液泡膜的CAX 主要是Ca2+轉運蛋白,介導Ca2+從細胞質進入液泡,同時也具有轉運Cd2+、Zn2+和Mn2+等離子的能力。 對參與 Cd轉運的CAX基因研究較多的是擬南芥。 擬南芥中包含6個CAX家族成員,即CAX1 ~CAX6,均具有轉運Cd 的能力,其中CAX2和CAX4編碼蛋白對Cd 的轉運能力最強[36-37]。 Korenkov 等[38-39]將At-CAX2和AtCAX4在煙草中表達發現,煙草根部細胞質中Cd、Zn 和Mn 向液泡轉運增加,并且CAX2和CAX4對Cd 吸收最敏感。 這可能是由于Cd2+具有與Ca2+相似的離子半徑,使得這2個離子競爭相同的膜轉運蛋白[40]。 最近,廖瓊等[41]發現甘藍型油菜CAX家族共包含17個成員,并且CAX基因主要在地下部表達,其中BnaC4.CAX1-2和BnaC3.CAX2-2是該家族的核心基因,并受Cd2+脅迫的顯著誘導。 陳少鵬等[42]克隆了越橘中VcCAX2基因,發現該基因在越橘根系中表達量隨著Cd 脅迫程度的提高呈現增加的趨勢。
此外,ATP 結合盒轉運蛋白(ABC)在植物細胞Cd 的外排過程中也發揮著重要轉運作用。 MRP(ABCC)和PDR(ABCG)是目前研究較為深入的ABC轉運蛋白亞族。 如 Brunetti 等[43]通過Cd 特征熒光物定位實驗證明AtABCC3存在時,Cd 在擬南芥液泡中的分布遠多于細胞質,并且推測AtABCC3 將Cd 以植物螯合肽(PCs)-Cd 復合體的形式轉運到液泡區隔。 Kim 等[44]發現了定位于擬南芥根部表皮細胞質膜的AtPDR8 具有將Cd 排出細胞外的功能。
總之,由于Cd 是植物的非必需元素,因此植物體內沒有Cd 的特異性轉運蛋白,Cd 在植物體內的跨膜轉運主要利用特異性較低的必需元素Fe2+、Zn2+和Mn2+等二價陽離子的轉運蛋白。 Cd 在植物根系吸收、轉運的主要轉運蛋白如圖1 所示。
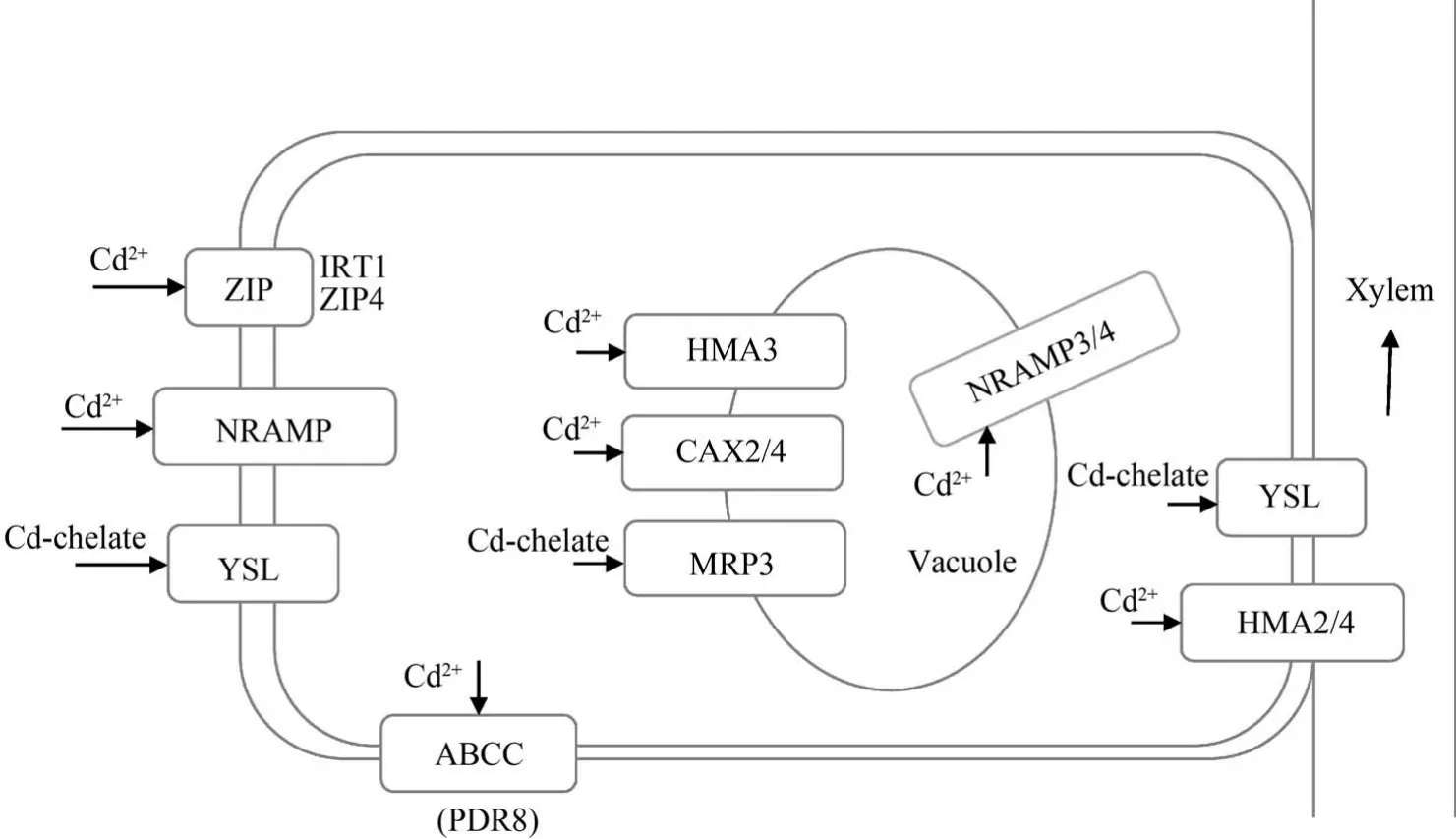
圖1 鎘在植物體內吸收、轉運的主要轉運蛋白示意圖[45]Fig.1 Schematic diagram of the main transporters involved in Cd uptake and translocation in plants[45]
2 植物鎘耐性調控機制(The tolerance mechanism of cadmium in plants)
2.1 轉錄因子調控
轉錄因子(transcriptional factors, TFs)可通過與特定基因的順式作用元件結合調控下游基因的表達,在植物響應逆境脅迫的過程中發揮重要作用[46]。 大量研究表明,WRKY、MYB 和bHLH 等轉錄因子家族均參與植物Cd 耐性的調控。
WRKY 蛋白是植物特有的一類轉錄因子,同時也是植物中最大的轉錄因子家族之一,主要通過與啟動子區域的W-box 元件結合調控下游靶基因的表達。 如大麻的40個WRKY 轉錄因子中有14個成員參與 Cd 脅迫響應[47]。 Liu 等[48]的研究表明,AtWRKY18、AtWRKY40 和 AtWRKY60 能與特定基因啟動子區的W-box 結合,從而負調控擬南芥的Cd 耐性。 AtWRKY12 對擬南芥 Cd 耐性也具有負調控作用。 Han 等[49]揭示了 AtWRKY12 作用機制主要是其能與GSH1啟動子結合,直接抑制GSH1基因的表達,間接干擾Cd 解毒物質PCs 的合成。而Sheng 等[50]則發現擬南芥的WRKY13 轉錄因子可以與PDR8啟動子直接結合,激活Cd 排出蛋白PDR8 的表達,減少細胞內Cd 積累,正調控擬南芥的 Cd 耐性。 Hong 等[51]用 Cd 處理玉米幼苗,發現ZmWRKY4、超氧化物歧化酶(SOD)和抗壞血酸過氧化物酶(APX)基因表達上調,并推測可能是由于ZmWRKY4可以激活ZmSOD4和ZmcAPX的表達,從而促進抗氧化酶的合成,提高玉米的Cd 耐性。 此外,在Cd 超積累植物中也發現了參與調控Cd 耐性的WRKY 轉錄因子。 如王影等[52]從超積累型東南景天中獲得了一條新的東南景天WRKY 轉錄因子家族基因——SaWRKY7,并發現其在早期響應Cd 脅迫誘導表達,推測其可能在東南景天Cd 積累或Cd 耐受過程中發揮重要作用。
MYB 轉錄因子是一類結構中含有一段高度保守的 DNA 結構域(MYB)的轉錄因子,分為1RMYB、R2R3-MYB、R1R2R3-MYB 和 4R-MYB 這 4類,其中R2R3-MYB 數量最多,多數R2R3-MYB 轉錄因子可以正調控植物對逆境的響應。 如丁杰等[53]從大豆根系中獲得了16個與Cd 脅迫應答相關的R2R3-MYB基因,進一步分析發現,大豆根系中的MYB基因主要通過調控重金屬Cd 的吸收、轉運、解毒過程來緩解Cd 的毒害作用。 Agarwal 等[54]研究發現,擬南芥中R2R3-MYB 亞家族的AtMYB4基因過表達可顯著提高植株的Cd 耐性,并認為主要是由于MYB4 能特異性激活PCS1和MT1C基因的表達,促進PCs 和金屬硫蛋白(MTs)的合成,從而降低Cd 毒性。 除了直接作用于Cd 解毒物質合成過程的基因,MYB 轉錄因子還可以通過脫落酸(ABA)信號途徑調控植物Cd 耐性。 如Zhang 等[55]發現擬南芥R2R3-MYB 轉錄因子MYB49 具有正調控Cd耐性作用,并認為MYB49 主要通過與ABI5互作,干擾MYB49 與下游基因bHLH38和bHLH101的直接結合,減少Cd 吸收蛋白IRT1 的合成,提高植株對 Cd 的耐受性。 此外,Hu 等[56]報道了水稻中1R-MYB 亞家族基因OsMYB45可被Cd 誘導表達,該基因突變會抑制OsCATA和OsCATA的表達,降低過氧化氫酶(CAT)的活性,這可能是MYB45 轉錄因子調控Cd 耐性的方式。
bHLH 轉錄因子家族成員均含有由50 ~60個氨基酸組成的典型bHLH 結構域,該結構域分為N端的堿性區域和C 端的螺旋-環-螺旋(HLH)區域2個不同的功能區[57]。 堿性區域通常包括10 ~18個氨基酸,具有DNA 的結合位點,能夠識別靶基因啟動子區的 E-box 和/或 G-box 元件;HLH 區域由 2個α-螺旋以及連接2個螺旋的環狀結構組成,是二聚體形成的結合域[58]。 Wu 等[59]研究發現 Cd 脅迫可以提高擬南芥中bHLH家族基因FIT、AtbHLH38和AtbHLH39的表達,并且過表達FIT/AtbHLH38或FIT/AtbHLH39可增強植株Cd 耐性。 進一步實驗證明FIT/AtbHLH38或FIT/AtbHLH39過表達一方面能夠啟動重金屬區室化基因的表達(如HMA3、MTP3、IREG2和IRT2),從而將 Cd 區隔在根部,抑制Cd 向地上部分轉運;另一方面還能啟動尼克酰胺合成酶基因(NAS1和NAS2)的表達,從而促進植物體內鐵螯合物尼克酰胺(NA)的合成,增加Fe 的吸收轉運,減少 Cd 的積累。 最近,Yao 等[60]在擬南芥中也有類似發現:bHLH104 能夠正調控4個重金屬解毒基因的表達,即IREG2、MTP3、HMA3和NAS4,從而提高擬南芥的 Cd 耐性。 此外,人們在大豆中也發現了參與其Cd 耐性調控的bHLH 轉錄因子。 如Xu 等[61]發現 Cd 處理會誘導大豆根部bHLH 轉錄因子 GmORG3 表達上調, 過表達GmORG3可以提高植物的Cd 耐性,其作用機制可能是由于植物對Fe 的吸收轉運增強,從而限制了Cd 向地上部分的轉運。 劉曉慶等[62]則從大豆根系中篩選得到了響應Cd 脅迫的GmbHLH041基因。實驗發現,GmbHLH041基因在 Cd 脅迫24 h 后表達量顯著提高,說明該基因在大豆Cd 脅迫應答中可能發揮著重要作用。
2.2 microRNA 調控
microRNA(miRNA)是一類長度約20 ~24個核苷酸、內源性、非編碼RNA 分子,可通過與靶序列互補結合,降解靶mRNA、抑制翻譯、DNA 甲基化,在轉錄和轉錄后水平調控靶基因的表達,是植物生長發育及應答非生物脅迫過程中的重要調控因子[63]。 近年來,人們利用微陣列芯片技術和高通量測序技術在不同植物中檢測出大量Cd 脅迫應答相關的miRNA。 如Ding 等[64]采用微陣列芯片技術在水稻中識別出19個Cd 脅迫應答相關miRNA,其中 miR528 表達上調,其余 18個表達下調。Tang 等[65]則通過高通量測序技術從水稻中鑒定出163個Cd 脅迫差異表達miRNA,并且根部與地上部展示出不同的基因表達變化。 最近,Jian 等[66]采用高通量測序技術對甘藍型油菜中Cd 脅迫應答的miRNA 進行全基因組分析,共鑒定出39個差異表達miRNA,包括8個已知的和31個新的miRNA,并推測miRNA 可能通過調控轉錄因子表達、生物脅迫防御、離子穩態及次生代謝物的合成以響應Cd 脅迫。
目前,關于植物miRNA 對Cd 脅迫的響應機制研究也在逐步開展,已有許多響應Cd 脅迫的miRNA 及其對應靶基因的研究報道(圖2)。 Cd 脅迫相關miRNA 的靶基因多為調節性基因,包括調控下游基因表達的轉錄因子和參與信號轉導的蛋白激酶等。 miR166 的靶基因為同源異形域-亮氨酸拉鏈(HD-ZIP)轉錄因子。 Ding 等[67]發現 Cd 脅迫下水稻根系中 miR166 表達受到顯著抑制,而過表達miR166 則可以降低Cd 向地上部的轉運,減少籽粒中Cd 積累。 進一步實驗發現,miR166 可能通過負調控其靶基因OsHB4(一種HD-ZIP 轉錄因子)的表達,進而調節OsHMA2和OsHMA3等Cd 耐性基因的表達,影響水稻中 Cd 的吸收、轉運和解毒。miR390 的靶基因則為富含亮氨酸的類受體蛋白激酶(RLK)[68]。 水稻過表達 miR390 可以顯著抑制miR390 靶基因OsSPK(一種RLK)的表達,但OsSPK與Cd 轉運蛋白之間的調控關系有待進一步驗證。此外,miRNA 對Cd 脅迫的響應還涉及抗氧化防御方面。 如miR398 靶基因是可清除活性氧(ROS)的CSD,Cd 脅迫下miR398 在小麥葉中表達下調會導致CSD 活性提高,增強植株的Cd 耐性[69]。
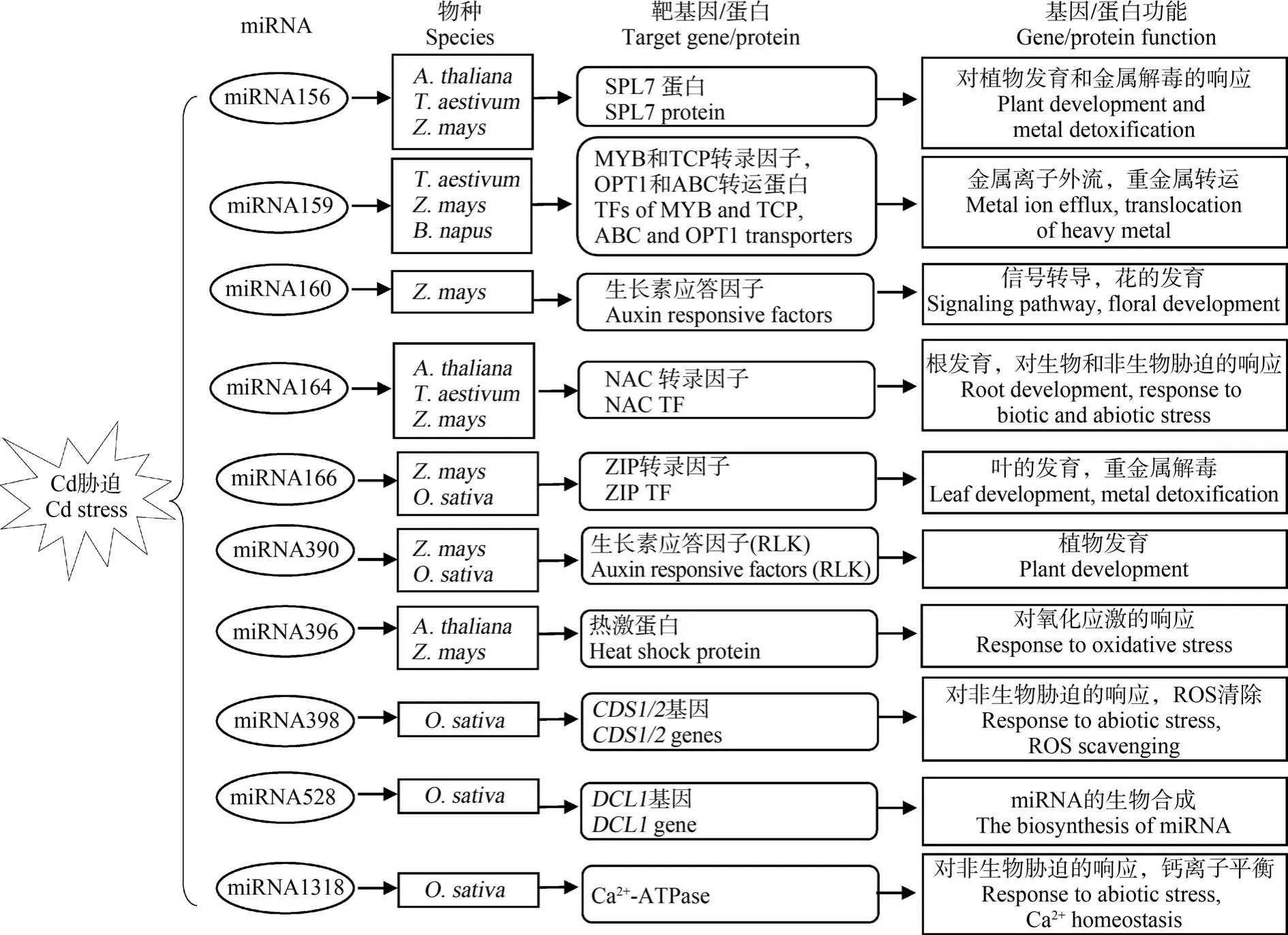
圖2 響應Cd 脅迫的miRNA 及其靶基因[63,70-71]注:A. thaliana 為擬南芥,T. aestivum 為小麥,Z. mays 為玉米,B. napus 為甘藍型油菜,O. sativa 為水稻;ROS 表示活性氧。Fig.2 miRNAs and their target genes in response to Cd stress[63,70-71]Note: A. thaliana is Arabidopsis thaliana; T. aestivum is Triticum aestivum; Z. mays is Zea mays; B. napus is Brassica napus;O. sativa is Oryza sativa; ROS stands for reactive oxygen species.
目前,關于植物對重金屬Cd 的耐性機制研究不斷深入。 通常,Cd 離子進入植物細胞后會影響其正常的代謝活動。 植物感受到Cd 離子的脅迫后,能夠通過信號傳導途徑激活體內抗氧化酶及相關金屬轉運蛋白的表達,從而避免或緩解Cd 離子對細胞的毒害作用。 在轉錄及轉錄后水平上,植物的鎘耐性涉及一系列復雜的調控網絡,在今后的研究中有待進一步探索和挖掘。
3 總結與展望(Summary and prospect)
Cd 污染是一個嚴重的環境問題。 土壤中的Cd進入植物細胞后不僅影響植物的生長發育與代謝過程,還會經過食物鏈傳遞危害人體健康。 因此,Cd在植物體內的吸收和轉運一直是研究的熱點。 目前,研究者對于土壤中的Cd 經植物根系吸收,并向地上部分轉運過程已經有了較好了解,尤其是Cd 轉運蛋白功能研究取得了一定進展。 同時,隨著在生理生化水平上對植物Cd 解毒機制認識的深入,其背后的分子調控機制也在逐漸得以揭示,已發現多個參與調控植物Cd 耐性基因表達的轉錄因子及miRNA。 但植物對Cd 的吸收、轉運是一個十分復雜的過程,關于植物Cd 耐性的具體分子調控機制仍有待進一步探索。 如解析Cd 轉運蛋白的晶體結構是理解這些轉運蛋白的分子作用機制的關鍵;探究Cd 脅迫下不同轉運蛋白之間的相互關系及協調作用有助于明確這些轉運蛋白的功能調控;挖掘響應Cd 脅迫轉錄因子和miRNA 的靶基因將成為闡明Cd 耐性基因表達調控分子機理的主要研究內容。 此外,植物遭受Cd 脅迫時,細胞內存在一系列信號傳遞,今后的研究還應關注這些信號因子對基因表達的影響,以及不同信號通路之間的對話,從而進一步明確植物響應Cd 脅迫的調控網絡。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
