徽式臘肉刀板香中優勢微生物的分離鑒定
2022-06-28 05:17:07朱旱林,徐寶才,黃千里
肉類研究 2022年5期
朱旱林,徐寶才,黃千里





摘? 要:刀板香作為一種傳統的徽派腌臘肉制品,品質優良、風味獨特,但生產工藝相對落后、品質不穩定,因此,需要對刀板香中優勢微生物的分離鑒定及相應發酵劑研制的需求迫切。通過低溫選擇性培養和分子生物學鑒定,共分離得到5 株葡萄球菌(4 株馬胃葡萄球菌和1 株木糖葡萄球菌)、6 株嗜冷菌(1 株小束噬脯氨酸菌、1 株沙雷氏菌和4 株嗜冷桿菌),進一步測馬胃葡萄球菌(A3)、木糖葡萄球菌(A4)、小束噬脯氨酸菌(L1)和嗜冷桿菌(N17)的溫度適應、耐鹽和耐乙醇能力。結果表明:菌株A3和A4有較好的耐鹽能力,菌株L1和A4對溫度的適應能力較好,4 株菌對于乙醇的耐受性均較差。因此,篩選得到菌株A3和A4可作為良好的發酵劑,菌株L1和N17有作為低溫發酵劑的潛力。
關鍵詞:刀板香;優勢微生物;發酵劑;分離純化;鑒定;耐受性
Isolation and Identification of Dominant Microorganisms in “Daobanxiang” Cured Meat
ZHU Hanlin, XU Baocai, HUANG Qianli*
(School of Food and Biological Engineering, Heifei University of Technology, Heifei? ? ?230009, China)
Abstract: As a traditional Huizhou-style cured meat product, Daobanxiang (DBX) has excellent quality and unique flavor. As its production technology is relatively outdated and its quality is unstable, there is an urgent need to isolate and identify the dominant microorganisms in DBX and develop starter cultures for its production. Through low temperature selective culture and molecular biological identification, four strains of Staphylococcus equinovarus, one strain of S. xylose, and six strains of psychrophilic bacteria, including one strain of Prolinoborus fasciculus, one strain of Serratia and four strains of Psychrobacter were obtained. Furthermore, the salt tolerance, temperature acclimation and ethanol tolerance of S. equinovarus A3, S. xylose A4, Prolinoborus fasciculus L1 and Psychrobacter N17 were determined. It turned out that S. equinovarus A3 and S. xylose A4 had better salt tolerance, P. fasciculu L1 and S. xylose A4 had better adaptability to temperature, and all the four strains had weak tolerance to ethanol. Strains A3 and A4 can be used as good starter cultures, and strains L1 and N17 have the potential to be used in low-temperature fermentation.
Keywords: Daobanxiang; dominant microorganism; starter cultures; isolation and purification; identification; tolerance
DOI:10.7506/rlyj1001-8123-20220322-021
中圖分類號:TS201.3? ? ? ? ? ? ? ? ? ? ? ? ? 文獻標志碼:A? ? ? ? ? ? ? ? ? ? ? ? ? ? 文章編號:
引文格式:
朱旱林, 徐寶才, 黃千里. 徽式臘肉“刀板香”中優勢微生物的分離鑒定[J]. 肉類研究, 2022, 36(5):? . DOI:10.7506/rlyj1001-8123-20220322-021.? ? http://www.rlyj.net.cn
ZHU Hanlin, XU Baocai, HUANG Qianli. Isolation and identification of dominant microorganisms in “Daobanxiang” cured meat [J]. Meat Research, 2022, 36(5):? . DOI:10.7506/rlyj1001-8123-20220322-021.? ? http://www.rlyj.net.cn
腌臘制品在我國有著上千年的歷史,深受人們的喜愛。生肉經過腌制和成熟后,肉色臘紅,耐貯存,有著獨特的腌臘風味[1-2]。傳統腌臘肉制品開放式的制作模式、漫長的制作周期以及適宜微生物生長的條件,是滿足微生物繁殖、代謝的絕佳天然培養基[3-4]。加之千百年的工藝傳承、特定的區域環境,一定程度上是對相關微生物的定向馴化與選擇,或許也正是因此才成就了我國多種地區性腌臘肉制品。
刀板香是安徽黃山地區特有的一種腌臘肉制品,由當地土豬肉制成,肥而不膩,具有極強的地域特色,這與黃山地區獨特的氣候環境有著密不可分的關系,但難以實現異地生產。黃山屬亞熱帶季風濕潤氣候,獨特的水土環境和氣候條件使得當地形成了獨特穩定、成分復雜且難以復制的微生物群落環境,這也可能是刀板香獨特風味形成的主要因素。刀板香原料采用徽州地區土雜豬厚度均勻的五花肉,在適當的環境條件下(溫度、濕度),經預處理、腌制、晾曬(輕發酵)等核心工藝加工而成,具有典型特征品質,是體現濃郁地方特色的中式傳統肉制品。微生物可通過自身分泌的豐富酶系參與肉制品發酵中的一系列過程,如脂肪氧化[5-6]、蛋白質降解[7-8]、碳水化合物水解[9]等,生成醛、酸、酮、酯和短肽等物質,而這些物質存在的量及相互間的比例,對區域腌臘肉制品獨特風味和色澤的形成影響顯著。
究竟哪些微生物在腌臘肉制品風味形成過程中起重要作用?黃艾祥[10]研究云南干腌火腿品質特征形成規律及微生物對其品質特征形成的作用,結果發現,微生物將火腿中蛋白質、脂肪降解生成一系列的游離氨基酸和脂肪酸。此外,Lizaso等[11]研究西班牙干腌香腸成熟過程中脂質和蛋白質的變化,發現高亞硝酸細菌和微球菌引起蛋白質的水解,導致香腸中游離氨基酸含量增加,對最終的感官品質形成具有重要作用。Zhao Dandan等[12]研究不同發酵階段干腌魚風味與微生物之間的關系,發現葡萄球菌是干腌魚發酵過程的優勢菌屬,參與發酵過程風味的形成。Prado等[13]研究西班牙傳統發酵香腸中微生物的作用,發現主要微生物為植物乳桿菌,該菌有效抑制了腐敗微生物的生長并促進了風味的形成。可見,不同的腌臘肉制品中有不同的優勢微生物。從中篩選出產生最初產品獨特風味、優良品質的菌種并研發發酵劑成為新的趨勢。
目前,較多研究直接將已知的發酵劑應用到肉制品中,據此分析不同發酵劑對特定肉制品品質的影響,如Ravyts[14]、Stahnke[15]、Hu Yingying[16]等將發酵劑直接應用到干腌香腸中。從傳統的肉制品中分離,并通過一系列篩選得到具有良好發酵性能的菌株,有助于新發酵劑的開發。不同發酵肉制品具有不同的特征,其中關鍵發酵微生物的篩選標準并不一致,因此需要對傳統肉制品建立符合其特征的篩選體系。刀板香作為一種傳統的徽派腌臘肉制品,仍然依賴作坊式生產,工藝相對落后、品質不穩定,已經不適于現代社會的需求;其中優勢微生物的分離鑒定及相應發酵劑研制的需求迫切。本研究組前期[17]采用高通量測序技術對不同加工階段刀板香中的微生物進行測序,發現刀板香的主要優勢菌屬為葡萄球菌和嗜冷桿菌,這與刀板香生產環境為低溫環境相吻合。鑒于此,本研究在低溫環境下對刀板香的優勢菌屬進行選擇性分離培養,采用形態學觀察、生理生化鑒定和分子生物學的方法對分離純化的微生物進行菌株鑒定,以獲得在刀板香風味形成過程中發揮重要作用的菌株,進而建立刀板香發酵劑的篩選體系,對于優良微生物種質資源的挖掘以及地方傳統特色腌臘肉制品的規模化、標準化生產均有重要意義。
1? ?材料與方法
1.1? ?材料與試劑
刀板香冬季成品取自安徽黃山全江生態農業科技有限公司。
MSA培養基、LB培養基、營養瓊脂、革蘭氏染色液、產H2S培養基、硝酸鹽還原培養基、氨基酸脫羧酶培養基、生化鑒定管? ?廣東環凱微生物科技有限公司。
細菌基因組DNA快速抽提試劑盒、Taq酶、引物、Marker、瓊脂糖? ?生工生物工程(上海)股份有限公司。
1.2? ?儀器與設備
ME204E-02型電子分析天平? ? 瑞士梅特勒-托利多儀器有限公司;Synergy H1酶標儀? ?美國BioTek儀器公司;BSD-150型振蕩培養箱? ?上海博迅醫療生物儀器股份有限公司;AC2-4S1型超凈工作臺? ?蘇州安泰空氣技術有限公司。
1.3? ?方法
1.3.1? ?樣品制備
原料在低溫下使用粗鹽腌制,輔以少量的白酒和香辛料,腌制時間為30 d。腌制結束后經過冷水洗鹽,然后轉移到室外晾曬30 d左右,晾曬后進行真空包裝即得到成品。
1.3.2? ?菌株的分離和純化
依據GB 4789.2—2016《食品安全國家標準 食品微生物學檢驗 菌落總數測定》[18]并進行部分修改。在無菌條件下,稱取10 g刀板香樣品于無菌均質袋中,再加入90 mL生理鹽水,利用拍打式均質機將樣品拍打1~2 min,制成1︰10的樣品勻液,然后將其進行10 倍梯度稀釋為質量濃度分別為10-3、10-4、10-5 g/mL,分別涂布于MRS培養基,放置于10 ℃培養48 h,涂布于LB培養基,放置于4 ℃培養7 d左右,挑取生長狀況良好的單菌落,劃線分離純化2~3 次。
1.3.3? ?形態學觀察
將分離純化得到的菌株分別在MRS培養基和LB培養基進行劃線,37 ℃培養24 h后觀察菌落生長情況,并通過革蘭氏染色和顯微鏡觀察其菌落狀態,選取革蘭氏陽性菌株。
1.3.4? ?菌株篩選實驗
產黏性實驗:直接挑取平板上的單菌落觀察,篩選出不產黏液的菌株。
發酵葡萄糖產氣實驗[19]:將平板上的菌株接種于葡萄糖發酵管(內置杜氏小管)中,放置于37 ℃培養18~24 h,若內置倒管內有氣泡產生,則菌株產氣。
產過氧化氫實驗[20]:用接種環挑取平板上的單菌落于無菌玻片上,滴加一滴體積分數3%的過氧化氫試劑,觀察是否有氣泡產生。
產H2S實驗[19]:用接種針挑取平板上的單菌落穿刺接種于H2S生化鑒定管中,于37 ℃培養18~24 h,若顯黑色則呈陽性,否則為陰性,篩選出不產H2S的菌株。
產氨實驗[19]:將待測菌株接種于產氨培養基中,以不接種作為空白對照,37 ℃培養24 h,在菌液中加入3~5 滴奈氏試劑,有黃色或棕紅色沉淀出現者為陽性,無沉淀為陰性,篩選出不產氨的菌株。
硝酸鹽還原酶實驗[21]:挑取平板上的單菌落接種于硝酸鹽還原酶的生化鑒定管中,培養18~24 h后,加入硝酸鹽還原試劑甲、乙液各2 滴,混勻,若顯深紅色則為陽性,篩選出硝酸鹽還原酶陽性的菌株。
氨基酸脫羧酶實驗[20]:將待測菌株接種于氨基酸脫羧酶生化鑒定管的實驗管和對照管中,并加入滅菌液體石蠟5~8 滴覆蓋培養基表面;培養18~24 h,若實驗管為黃綠色,對照管變黃,則呈陽性,篩選氨基酸脫羧酶陰性的菌株。
蛋白酶檢測實驗[22]:將待測菌株以點種法接種于酪蛋白檢測培養基上,37 ℃培養48 h后,在菌落周圍滴加10 g/100 mL三氯乙酸溶液,若菌落周圍有透明圈出現,則為陽性。
脂肪酶檢測實驗[22]:將待測菌株以點種法接種于脂肪酶檢測培養基上,37 ℃培養48 h后,若菌落顏色變紅,則為陽性。
1.3.5? ?分子生物學鑒定
挑取平板上的單菌落接種于營養瓊脂培養基中,37 ℃培養過夜,取1.5 mL菌液12 000 r/min離心2 min后,棄去上清,然后按照細菌基因組DNA快速抽提試劑盒說明書操作,所得的基因組利用0.02 g/mL瓊脂糖凝膠電泳查看提取結果。
16S rDNA擴增序列所用引物,正向引物為27F:5′-AGAGCCTGGCTCAGTTTGAT-3′;反向引物為1492R:5′-GGTTACCTTGTTACGACTT-3′。
聚合酶鏈反應(polymerase chain reaction,PCR)體系(50 μL):上、下游引物各1 μL、模板DNA1 μL、2×Taq Master Mix 25 μL、dd H2O 21μL[23]。
將電泳檢測后的PCR產物送至生工生物工程(上海)股份有限公司進行測序,獲得16S rRNA基因序列。測序結果在美國國家生物技術信息中心(National Center of Biotechnology Information,NCBI)的比對搜索工具(basic local alignment search tool,BLAST)分析比對。
1.3.6? ?菌株生長曲線和耐受特性測定
菌株生長曲線測定:將篩選得到的菌株接種于營養肉湯培養基中,37 ℃培養,每隔2 h測1 次600 nm波長處光密度(OD600 nm)。
菌株對溫度適應能力:將篩選得到的菌株接種于營養肉湯培養基中,分別在4、10、15、25、37、45 ℃培養24 h后,測其OD600 nm。
菌株耐鹽能力測定:將篩選得到的菌株接種于鹽(NaCl)質量濃度分別為0、2、4、6、8 g/100 mL的營養肉湯培養基中,培養24 h后,測其OD600 nm。
菌株耐乙醇能力測定:將篩選得到的菌株接種于乙醇體積分數分別為0%、3%、6%、9%、12%的營養肉湯培養基中,培養24 h后,測其OD600 nm。
1.4? ?數據處理
所有數據測定均重復3 次,使用Excel 2019軟件(Microsoft,USA)進行數據統計和分析,使用Origin 2020軟件(OriginLab,USA)進行繪圖。使用MEGA 7軟件(version 7.0,Mega Limited,Auckland,New Zealand)對分離、純化優勢菌株的測序結果構建進化樹。
2? ?結果與分析
2.1? ?細菌的菌落形態和鏡檢結果
通過對刀板香中葡萄球菌和嗜冷菌的選擇性分離培養,MSA培養基得到35 株菌株,LB培養基得到40 株菌株。MSA培養基形成圓形凸起、邊緣整齊、表面光滑、濕潤、不透明的菌落,LB培養基培養7 d后形成淺黃色、圓形、邊緣整齊、不透明的菌落。通過革蘭氏染色和鏡檢,共篩選到27 株革蘭氏陽性葡萄球菌和32 株革蘭氏陰性嗜冷菌。
2.2? ?菌株篩選結果
利用發酵劑的主要目的是縮短加工周期,改善肉制品的質地、風味和色澤,延長食品的貨架期[24]。因此對于發酵劑的篩選主要有以下標準:不產黏液、發酵葡萄糖不產氣、過氧化氫酶陽性、不產H2S、不產氨、具有較高的亞硝酸鹽還原酶活性和不具有氨基酸脫羧酶活性[25]。
對于革蘭氏染色和鏡檢后篩選到的菌株進一步篩選,由表1可知,27 株葡萄球菌和32 株嗜冷菌不產黏液,20 株葡萄球菌和26 株嗜冷菌不產氣,18 株葡萄球菌和20 株嗜冷菌不產H2S,15 株葡萄球菌和19 株嗜冷菌不產氨,13 株葡萄球菌和14 株嗜冷菌具有亞硝酸鹽還原酶活性,8 株葡萄球菌和6 株嗜冷菌不具有氨基酸脫羧酶活性。
將這些初步篩選到的菌株作進一步脂肪酶和蛋白酶活性檢測實驗,發現共有5 株葡萄球菌和6 株嗜冷菌具有較強的蛋白酶和脂肪酶活性。
2.3? ?分子生物學鑒定結果
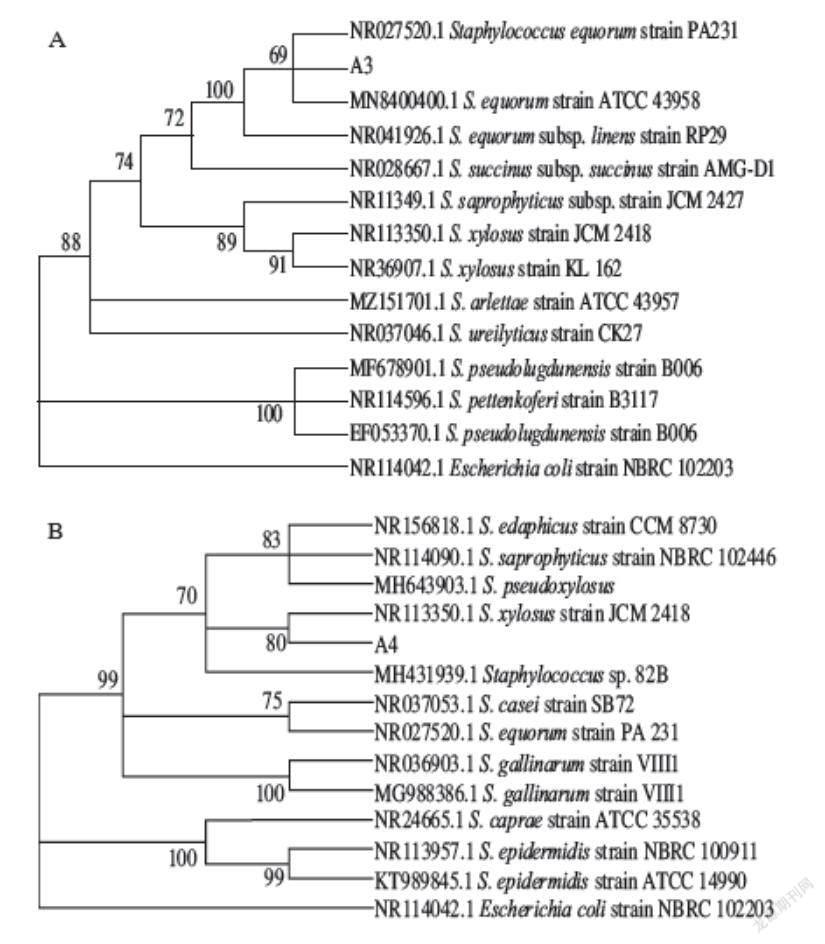
通過篩選得到5 株葡萄球菌和6 株嗜冷菌,將葡萄球菌分別命名為A3、A4、A7、A10、A11,嗜冷菌分別命名為L1、N1、L17、N3、N17、T7。將這些菌株的擴增序列測序結果在NCBI數據庫中進行比對,選取同源性≥99%的菌株序列,利用Mega 7.0軟件構建系統發育樹[26]。發現這幾株菌中有3 株菌與馬胃葡萄球菌(Staphylococcus equorum)序列相似度達到99%,2 株與木糖葡萄球菌(Staphylococcus xylosus)序列相似度達到99%,1 株與小束噬脯氨酸菌(Prolinoborus sp.)序列相似度達到99%,1 株與沙雷氏菌(Serratia sp.)序列相似度達到99%,4 株與嗜冷桿菌(Psychrobacter faecalis)序列相似度達到99%。由于發酵劑的選擇需要具有蛋白酶和脂肪酶活性,葡萄球菌具有較好的脂肪酶和蛋白酶活性,小束噬脯氨酸菌具有脂肪酶活性,而嗜冷桿菌則具有耐受低溫特性,因此選取1 株馬胃葡萄球菌A3、1 株木糖葡萄球菌A4、1 株小束噬脯氨酸菌L1、1 株嗜冷桿菌N17制作系統發育樹。
由圖1可知,菌株A3與Staphylococcus equorum strain PA 231和Staphylococcus equorum strain ATCC 43958在同一個分支上,且通過Bootstrap的驗證具有較高的置信度,因此可以將菌株A3初步確定為馬胃葡萄球菌。菌株A4與Staphylococcus xylosus strain JCM 2418在同一分支上,且通過Bootstrap的驗證具有較高的置信度,因此可以將菌株A4初步確定為木糖葡萄球菌。菌株L1與Prolinoborus fasciculus strain CIP 103579在同一分支上,且通過Bootstrap的驗證具有較高的置信度,因此可以將菌株L1初步確定為小束噬脯氨酸菌。菌株N17與Psychrobacter pulmonis strain CCUG 46240在同一分支上,且通過Bootstrap的驗證具有較高的置信度,因此可以將菌株N17初步確定為嗜冷桿菌。
大多數肉制品中分離的微生物主要為乳酸菌、葡萄球菌、酵母菌和霉菌等菌株,這些微生物可以分解蛋白質,是發酵肉制品風味物質的主要來源[27-28],也是篩選的發酵劑的主要來源。本實驗從刀板香分離得到的馬胃葡萄球菌和木糖葡萄球菌,都是肉制品中常見的發酵微生物。Irlinger等[29]從法國干酪中分離出馬胃葡萄球菌,發現它有助于發酵食品香氣的形成。Hu Meizhong等[30]從中國培根中篩選分離得到1 株木糖葡萄球菌(Staphylococcus xylosus P2),將其接種到肉制品中,發現是肉制品良好的發酵劑。Xiao Yaqing等[31]將木糖葡萄球菌接種到中國傳統干腌肉制品中,發現接種后肉制品的游離氨基酸和游離脂肪酸含量均增加。張詩意[32]、陳學云[33]等分別從威寧火腿和鹽干帶魚中分離得到木糖葡萄球菌和馬胃葡萄球菌。目前,對于小束噬脯氨酸菌和嗜冷桿菌的研究較少,小束噬脯氨酸菌可能參與肉制品脂肪氧化過程,嗜冷桿菌可以很好地適應低溫環境,還需要對這2 株菌株作進一步研究。
2.4? ?菌株生長情況和生長適應性測定結果
通過分子生物學共鑒定出11 株菌株,選取其中1 株馬胃葡萄球菌A3、1 株木糖葡萄球菌A4、1 株小束噬脯氨酸菌L1和1 株嗜冷桿菌N17,分別進行下一步的生長情況和耐受實驗測定。
2.4.1? ?菌株的生長情況
檢驗發酵劑中的功能菌能否在肉制品生產過程中生長良好是發酵劑的一項重要指標。由圖2可知:在0~2 h內,菌株處于遲緩期,OD600 nm變化不大;培養2 h后,菌株A3、A4和L1進入指數期,菌株N17在培養4 h后進入指數期,OD600 nm顯著上升,這說明菌株A3、A4和L1的調整期較短,能夠快速繁殖,此時菌株的生長旺盛,活力最強;菌株A3在培養24 h后進入穩定期,菌株A4在培養16 h后進入穩定期,菌株L1和N17在培養30 h后進入穩定期,菌株L1處于指數期時間最長,說明在發酵過程中可以長時間保持活力最強的狀態,有著很好的發酵能力。到達穩定期后,菌株A3、A4、L1和N17的OD600 nm分別為2.4、2.5、2.6和2.9,活菌數都可以達到109 CFU/mL。
2.4.2? ?菌株適應溫度的能力
溫度在發酵肉制品的生產過程起著至關重要的作用,一般情況下,發酵肉制品的腌制溫度為4 ℃,發酵溫度為20~25 ℃,因此對溫度的適應能力也是發酵劑的一項重要指標[34]。
由圖3可知:菌株A3和A4在4~37 ℃溫度條件下,隨著溫度的升高生長能力不斷增強;菌株L1在4~25 ℃溫度條件下,隨著溫度的升高生長能力不斷增強;菌株N17在4~20 ℃溫度條件下,隨著溫度的升高生長能力不斷增強。菌株A3和A4在37 ℃培養,生長能力較其他溫度有顯著提高,且其他溫度下生長能力較低,說明這2 株菌株的溫度適應能力較差。菌株L1和N17的最適生長溫度分別為25 ℃和20 ℃,且在20~37 ℃菌株的生長能力較好,說明這2 株菌株的溫度適應能力較好,因此對于較低溫環境生產的發酵肉制品有著一定發酵作用。低溫環境可以抑制大多數腐敗菌的生長,可以有效延長產品的貨架期,這2 株菌對于開發低溫生產發酵肉制品有一定的潛力。4 株菌株在4、10、45 ℃下生長能力都較差,說明4 株菌株在較低溫和高溫狀態下生長能力都較差。但是肉制品的發酵溫度一般在25 ℃左右,所以4 株菌株都能適應肉制品的發酵溫度。
2.4.3? ?菌株的耐鹽能力
耐鹽能力是選擇發酵肉制品發酵劑的另一項重要指標,一般發酵肉制品的鹽含量均為6.0%~9.0%[35-36],這對于發酵肉制品的風味形成和安全性有重要的影響[37-38]。由圖4可知,菌株A3和A4在NaCl質量濃度為0~6 g/100 mL時,其生長能力較強,OD600 nm變化不明顯,在NaCl質量濃度達到8 g/100 mL時,菌株的生長能力受到一定程度的抑制,OD600 nm稍有下降,這說明菌株A3和A4有著較好的耐鹽能力,適合作為肉制品發酵劑。菌株L1和N17在NaCl質量濃度為0~4 g/100 mL時,其生長能力較強,而在NaCl質量濃度達到6 g/100 mL以上時,菌株的生長能力受到較大程度的抑制,且在NaCl質量濃度為8 g/100 mL時,菌株基本不生長,說明菌株L1和N17對鹽的耐受性較差,盡管發酵肉制品中鹽的質量分數一般為6.0%~9.0%,但由于目前發酵肉制品的生產都朝著低鹽的方向發展,加上發酵劑自身所帶有的抑菌作用,對于鹽的添加量可以適當降低,所以小束噬脯氨酸菌和嗜冷桿菌具有作為肉制品發酵劑的潛力。
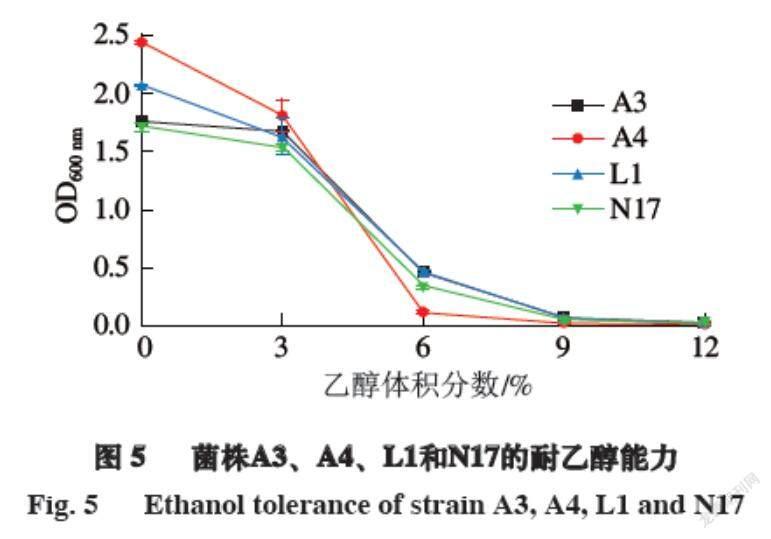
2.4.4? ?菌株的耐乙醇能力
在刀板香腌制的過程中都會添加少量的白酒用于殺菌去腥,因此菌株對于乙醇的耐受能力也是篩選發酵劑的重要指標。由圖5可知,在乙醇體積分數為0%~3%時,4 株菌株的生長能力較強,OD600 nm變化不明顯,而在乙醇體積分數達到6%時,菌株的生長能力有較大程度的下降,且在乙醇體積分數達到9%時,菌株的生長能力完全受到抑制,基本不生長。這說明4 株菌株對于乙醇的耐受程度較低,但由于發酵肉制品中添加的白酒含量較少,且在加工過程中有大部分揮發,所以耐乙醇能力對于菌株作為發酵劑影響較小。
3? ?結? 論
本研究對刀板香的優勢微生物進行分離純化,并通過生理生化實驗及分子生物學鑒定篩選得到4 株具有脂肪酶和蛋白酶活性、可作為發酵劑的菌株,包括2 株葡萄球菌A3和A4、1 株小束噬脯氨酸菌L1和1 株嗜冷桿菌N17。對4 株菌株作進一步的生物學特性和發酵特性分析,結果表明:菌株A3和A4分別在培養24、16 h進入穩定期,菌株L1和N17在培養30 h進入穩定期,菌株A3和A4生長較迅速,菌株L1在生長過程中可以長時間保持活力最強狀態,有較好的發酵潛力;菌株A3和A4對于溫度的適應性較差,而菌株L1和N17對于溫度的適應性較好,對于低溫發酵肉制品的開發有著一定的潛力;菌株A3和A4有著較好的食鹽耐受能力,而L1和N17的耐鹽能力則較差;4 株菌株對于乙醇的耐受能力均較差。菌株A3和A4可作為良好的發酵劑,L1和N17有作為低溫發酵劑的潛力。對刀板香的優勢微生物進行分離純化并對其發酵潛能進行研究,對于后續刀板香的工藝改善和標準化生產有很好的借鑒作用。
參考文獻:
[1] GUO Xin, HUANG Feng, ZHANG Hong, et al. Classification of traditional Chinese pork bacon based on physicochemical properties and chemometric techniques[J]. Meat Science, 2016, 117: 182-186. DOI:10.1016/j.meatsci.2016.02.008.
[2] 粟桂蓉, 彭鈺媛, 周璐璐, 等. 傳統土家臘肉加工過程中風味物質研究[J]. 食品科技, 2017, 42(3): 118-123. DOI:10.13684/j.cnki.spkj.2017.03.026.
[3] 周光宏, 趙改名, 彭增起. 我國傳統腌臘肉制品存在的問題及對策[J]. 肉類研究, 2003, 17(1): 3-7.
[4] ASHAOLU T J, KHALIFA I, MESAK M A, et al. A comprehensive review of the role of microorganisms on texture change, flavor and biogenic amines formation in fermented meat with their action mechanisms and safety[J]. Critical Review Food Science and Nutrition, 2021: 1-18. DOI:10.1080/10408398.2021.1929059.
[5] BRUNA J M, HIERRO E M, DE LA HOZ L, et al. Changes in selected biochemical and sensory parameters as affected by the superficial inoculation of Penicillium camemberti on dry fermented sausages[J]. International Journal of Food Microbiology, 2003, 85(2): 111-125. DOI:10.1016/s0168-1605(02)00505-6.
[6] 姬曉穎, 馬國源, 陳耀祥, 等. 腌制過程中亞硝酸鈉對羊肉脂肪酸組成以及脂質氧化的影響 [J]. 甘肅農業大學學報, 2020, 55(5): 195-202. DOI:10.13432/j.cnki.jgsau.2020.05.023.
[7] FLORES M, TOLDRá F. Microbial enzymatic activities for improved fermented meats[J]. Trends in Food Science and Technology, 2011, 22(2/3): 81-90. DOI:10.1016/j.tifs.2010.09.007.
[8] DE ALMEIDA M A, SALDANA E, DA SILVA PINTO J S, et al. A peptidomic approach of meat protein degradation in a low-sodium fermented sausage model using autochthonous starter cultures[J]. Food Research International, 2018, 109: 368-379. DOI:10.1016/j.foodres.2018.04.042.
[9] NIE Xiaohua, LIN Shengli, ZHANG Qilin. Proteolytic characterisation in grass carp sausage inoculated with Lactobacillus plantarum and Pediococcus pentosaceus[J]. Food Chemisty, 2014, 145: 840-844. DOI:10.1016/j.foodchem.2013.08.096.
[10] 黃艾祥. 云南干腌火腿品質特征形成與微生物作用研究 [D]. 重慶: 西南大學, 2006: 95-115.
[11] LIZASO G, CHASCO J, BERIAIN M J. Microbiological and biochemical changes during ripening of salchichón, a Spanish dry cured sausage[J]. Food Microbiology, 1999, 16: 219-228. DOI:10.1006/fmic.1998.0238.
[12] ZHAO Dandan, HU Jun, CHEN Wenxuan. Analysis of the relationship between microorganisms and flavour development in dry-cured grass carp by high-throughput sequencing, volatile flavour analysis and metabolomics[J]. Food Chemistry, 2022, 368: 130889. DOI:10.1016/j.foodchem.2021.130889.
[13] PRADO N, SAMPAYO M, GONZALEZ P, et al. Physicochemical, sensory and microbiological characterization of Asturian Chorizo, a traditional fermented sausage manufactured in Northern Spain[J]. Meat Science, 2019, 156: 118-124. DOI:10.1016/j.meatsci.2019.05.023.
[14] RAVYTS F, STEEN L, GOEMAERE O, et al. The application of staphylococci with flavour-generating potential is affected by acidification in fermented dry sausages[J]. Food Microbiology, 2010, 27(7): 945-954. DOI:10.1016/j.fm.2010.05.030.
[15] STAHNKE L H, HOLCK A, JENSEN A, et al. Maturity acceleration of Italian dried sausage by Staphylococcus carnosus-relationship between maturity and flavor compounds[J]. Journal of Food Science, 2020, 67(5): 1914-1921. DOI:10.1111/j.1365-2621.2020.08746.x.
[16] HU Yingying, CHEN Qian, WEN Rongxin, et al. Quality characteristics and flavor profile of Harbin dry sausages inoculated with lactic acid bacteria and Staphylococcus xylosus[J]. LWT-Food Science and Technology, 2019, 114: 108392. DOI:10.1016/j.lwt.2019.108392.
[17] 周瑩, 王兆明, 涂健, 等. 基于16S rRNA的徽派臘肉加工過程中微生物群落結構分析[J]. 肉類研究, 2021, 35(3): 1-7. DOI:10.7506/rlyj1001-8123-20201215-289.
[18] 中華人民共和國國家衛生和計劃生育委員會, 國家食品藥品監督管理總局. 食品安全國家標準 食品微生物學檢驗 菌落總數測定: GB 4789.2—2016[S]. 北京: 中國標準出版社, 2016.
[19] 東秀珠, 蔡妙英. 常見細菌系統鑒定手冊[M]. 北京: 科學出版社, 2001.
[20] SANTOS M H S. Amino acid decarboxylase capability of microorganisms isolated in Spanish fermented meat products[J]. International Journal of Food Microbiology, 1998, 39(3): 227-230. DOI:10.1016/S0168-1605(97)00129-3.
[21] 王燚. 抗氧化肉品發酵劑的篩選及其在羊肉香腸中的應用 [D]. 雅安: 四川農業大學, 2008: 38-40.
[22] OZTURKOGLU-BUDAK S, WIEBENGA A, BRON P A, et al. Protease and lipase activities of fungal and bacterial strains derived from an artisanal raw ewe’s milk cheese[J]. International Journal of Food Microbiology, 2016, 237: 17-27. DOI:10.1016/j.ijfoodmicro.2016.08.007.
[23] 牛雪峰. 發酵火腿中產香葡萄球菌的篩選及在發酵香腸中的應用[D]. 哈爾濱: 哈爾濱商業大學, 2020: 20-22.
[24] COLLINS-THOMPSON K L D A D L. Nitrite tolerance and nitrite reduction in lactic acid bacteria associated with cured meat products[J]. International Journal of Food Microbiology, 1984, 1: 163-170. DOI:10.1016/0168-1605(84)90007-2.
[25] 趙婧含, 鞠銘, 孫德鵬, 等. 火腿微生物發酵劑研究進展[J]. 農業與技術, 2021, 41(20): 16-21. DOI:10.19754/j.nyyjs.20211030004.
[26] 張坤煜, 趙舒欣, 王紅丁, 等. 神農架豆豉樣品中微生物的分離鑒定[J]. 綠色科技, 2020(20): 167-170. DOI:10.16663/j.cnki.lskj.2020.20.055.
[27] 劉柳, 劉學文, 張鑫燚. 發酵肉傳統制作工藝參數優化研究[J]. 食品工業科技, 2008(1): 202-204. DOI:10.13386/j.issn1002-0306.2008.01.054.
[28] 王正莉, 王衛, 陳林, 等. 傳統腌臘肉制品中微生物多樣性研究進展[J]. 食品研究與開發, 2021, 42(8): 202-206.
[29] IRLINGER F, LOUX V, BENTO P, et al. Genome sequence of Staphylococcus equorum subsp. equorum Mu2, isolated from a French smear-ripened cheese[J]. Journal of Bacteriology, 2012, 194(18): 5141-5142. DOI:10.1128/JB.01038-12.
[30] HU Meizhong, YU Jiansheng, YU Jianping, et al. Isolation and screening of Staphylococcus xylosus P2 from Chinese bacon: a novel starter culture in fermented meat products[J]. International Journal of Food Engineering, 2018, 15(2): 1-7. DOI:10.1515/ijfe-2018-0021.
[31] XIAO Yaqing, LIU Yingnan, CHEN Conggui, et al. Effect of Lactobacillus plantarum and Staphylococcus xylosus on flavour development and bacterial communities in Chinese dry fermented sausages[J]. Food Research International, 2020, 135: 109247. DOI:10.1016/j.foodres.2020.109247.
[32] 張詩意, 唐楠, 黃攀, 等. 威寧火腿中微生物的分離鑒定及其耐受特性[J]. 肉類研究, 2019, 33(8): 12-17. DOI:10.7506/rlyj1001-8123-20190625-147.
[33] 陳學云, 侯魯娜, 丁玉庭, 等. 鹽干帶魚中葡萄球菌的分離鑒定及其特性研究[J]. 肉類工業, 2010(5): 35-38.
[34] 馬德功, 王成忠, 崔文文. 發酵香腸乳酸菌發酵劑篩選標準[J]. 肉類研究, 2007, 21(12): 31-33.
[35] 馬德功. 發酵香腸中乳酸菌的分離,篩選及其應用[D]. 濟南: 山東輕工業學院, 2008: 5-11.
[36] DOS SANTOS B A, CAMPAGNOL P C B, FAGUNDES M B, et al. Adding blends of NaCl, KCl, and CaCl2 to low-sodium dry fermented sausages: effects on lipid oxidation on curing process and shelf life[J]. Journal of Food Quality, 2017, 2017: 7085798. DOI:10.1155/2017/7085798.
[37] OSHIBANJO D O, OLUSOLA O O, ADESHOLA A T, et al. Sensory acceptability and microbial load of partial, combined and total replacement of sodium chloride in beef sausage[J]. Annual Conference of the Nigerian Society for Animal Productio, 2018(1): 906-909.
[38]扈瑩瑩, 溫榮欣, 陳佳新, 等. 低鹽對發酵肉制品品質形成影響及減鹽手段研究進展[J]. 食品工業科技, 2019, 40(16): 324-328. DOI:10.13386/j.issn1002-0306.2019.16.054.
