紅豆杉葉際共生菌參與紫杉醇合成的宏基因組學分析
2022-07-04 06:56:40董華恒溫欣于淼陳營
農業(yè)與技術 2022年12期
董華恒 溫欣 于淼 陳營
(煙臺大學生命科學學院,山東 煙臺 264005)
紅豆杉是紅豆杉目紅豆杉科紅豆杉屬的一種喬木,又稱紫杉,也稱赤柏松,是世界上公認的瀕臨滅絕的天然珍稀抗癌植物。紅豆杉的藥用價值主要體現在其提取物——次生代謝衍生物——紫杉醇。紫杉醇是從紅豆杉的樹皮中分離提純的天然次生代謝產物,在臨床上已經廣泛用于乳腺癌、卵巢癌等的治療。紫杉醇作為一個具有抗癌活性的二萜生物堿類化合物,其廣泛而顯著的生物活性、全新獨特的作用機制等使其受到了植物學家、化學家、藥理學家、分子生物學家的極大青睞,使其成為20世紀下半葉舉世矚目的抗癌明星和研究重點[1,2]。但是從紅豆杉樹皮中分離提純的紫杉醇數量少,遠遠滿足不了臨床的需要[3,4],也有學者采用化學合成的方法,但其合成路線復雜、反應條件難以控制,合成率低[5],微生物發(fā)酵法生產雖具有生產周期短、操作簡單的優(yōu)勢,但是目前已知的微生物生產紫杉醇產量很低,不能夠滿足產業(yè)化生產要求,因此獲得能夠適用于發(fā)酵生產的工業(yè)菌株是實現發(fā)酵生產紫杉醇的前提。
宏基因組學是一種直接對微生物群體中包含的全部基因組信息進行研究的手段。其規(guī)避了對樣品中的微生物進行分離培養(yǎng),提供了一種對不可分離培養(yǎng)的微生物進行研究的途徑,更真實地反映樣本中微生物組成、互作情況,同時在分子水平對其代謝通路、基因功能進行研究[6,7]。本實驗基于宏基因組技術分析紅豆杉葉發(fā)酵液的微生物菌群,分析其主要代謝通路和合成紫杉醇的相關酶和微生物,以期為后續(xù)實驗提供優(yōu)勢菌種,從而縮短提取時間,提高紫杉醇的提取率。
1 材料與方法
1.1 材料與試劑
1.1.1 原料
采摘加拿大曼地亞紅豆杉針葉和枝條,置于冰箱內冷藏備用。
1.1.2 培養(yǎng)基
富集培養(yǎng)基:酵母膏0.5g,(NH4)2SO43g,NaCl 5g,KH2PO40.4g,MgSO40.3g,FeSO40.01g,蒸餾水1000mL。
1.2 實驗方法
1.2.1 樣品制備
取適量實驗室留存的紅豆杉枝葉樣品清洗晾干后取其葉片絞碎,樣品置于300mL錐形瓶中。配制1000mL富集培養(yǎng)基,121℃滅菌20min。向錐形瓶中加入10g針葉碎片,35℃、140rpm震蕩培養(yǎng),直至培養(yǎng)液渾濁并且針葉碎片出現明顯降解,為原代共生菌富集液。取原代富集液按5%接種含10%紅豆杉葉的富集培養(yǎng)基,35℃、140rpm震蕩培養(yǎng)4~5d,為一代菌群富集液。同樣操作至三代菌群富集液。每代菌群富集液為3個平行。
在超凈臺中分別吸取上述最終菌群富集液的3個平行樣品4mL,置于滅過菌的離心管中,8000rpm離心5min。取出后倒掉上清液,留底部沉淀,命名為h1、h2、h3,冷凍保存后送于公司進行微生物宏基因組分析。
1.2.2 宏基因組測序
本實驗樣本采用Illumina Novaseq平臺對測序樣本進行雙端測序。采用Illumina測序平臺測序獲得的原始數據(Raw Data)存在一定比例低質量數據,為了保證后續(xù)分析的結果準確可靠,需要對原始的測序數據進行預處理,包括質控和去宿主序列,獲取用于后續(xù)分析的有效序列(clean data)。3組數據經過過濾后剩余的序列數占Raw reads 的百分比相近。測序質量基本合格,可以用于后續(xù)分析。
1.2.3 物種組成分析
為了研究樣品物種組成及多樣性信息,用Kraken2對所有樣品的全部的有效序列進行注釋分類。繼續(xù)用Bracken對Kraken2得到的分類結果進行分類后貝葉斯重新估算豐度來估算宏基因組樣本的物種水平或屬級豐度。
1.2.4 功能數據庫注釋
使用HUMAnN2軟件,將質控和去宿主之后的序列與蛋白質數據庫進行比對,過濾掉比對失敗的reads,統(tǒng)計UniRef90各個蛋白的相對豐度,根據UniRef90 的ID和各個功能數據庫ID的對應關系,統(tǒng)計各個功能數據庫對應功能相對豐度。
2 結果與討論
2.1 紅豆杉葉際共生菌群的多樣性分析
本實驗首先是以紅豆杉葉為唯一碳源進行富集培養(yǎng),使富集到的菌群均為紅豆杉葉際共生菌,以排除其他雜菌的干擾,確保這些檢測到的菌種與紫杉醇的合成代謝相關。
由每個樣本在各分類水平注釋的相對程度可知,本實驗樣品的所有序列都能夠確定到種水平上。由圖1可以看出,3個樣本的絕對優(yōu)勢菌門為Ascomycota(子囊菌門)存在較高的相對豐度。在屬水平上,該樣本的優(yōu)勢菌屬有Wickerhamomyces(威克罕默氏屬)、Clavispora(棒孢酵母屬)、Meyerozyma(季也蒙念珠菌)、Aspergillus(曲霉屬)。在種水平上,Wickerhamomycesanomalus(威克罕默氏菌)、Clavisporalusitaniae(葡萄牙棒孢酵母)、Meyerozymaguilliermondii(季也蒙念珠菌)、Aspergillusnidulans(構巢曲霉)為樣本的優(yōu)勢菌種,且威克罕默氏屬的威克罕默氏菌為絕對優(yōu)勢菌種,存在較高的相對豐度。上述檢測結果與Flores-Bustamante ZR等[8]的研究結果一致,即紫杉醇的生產者主要屬于子囊菌門,故這些菌種可能與紫杉醇的合成有關。
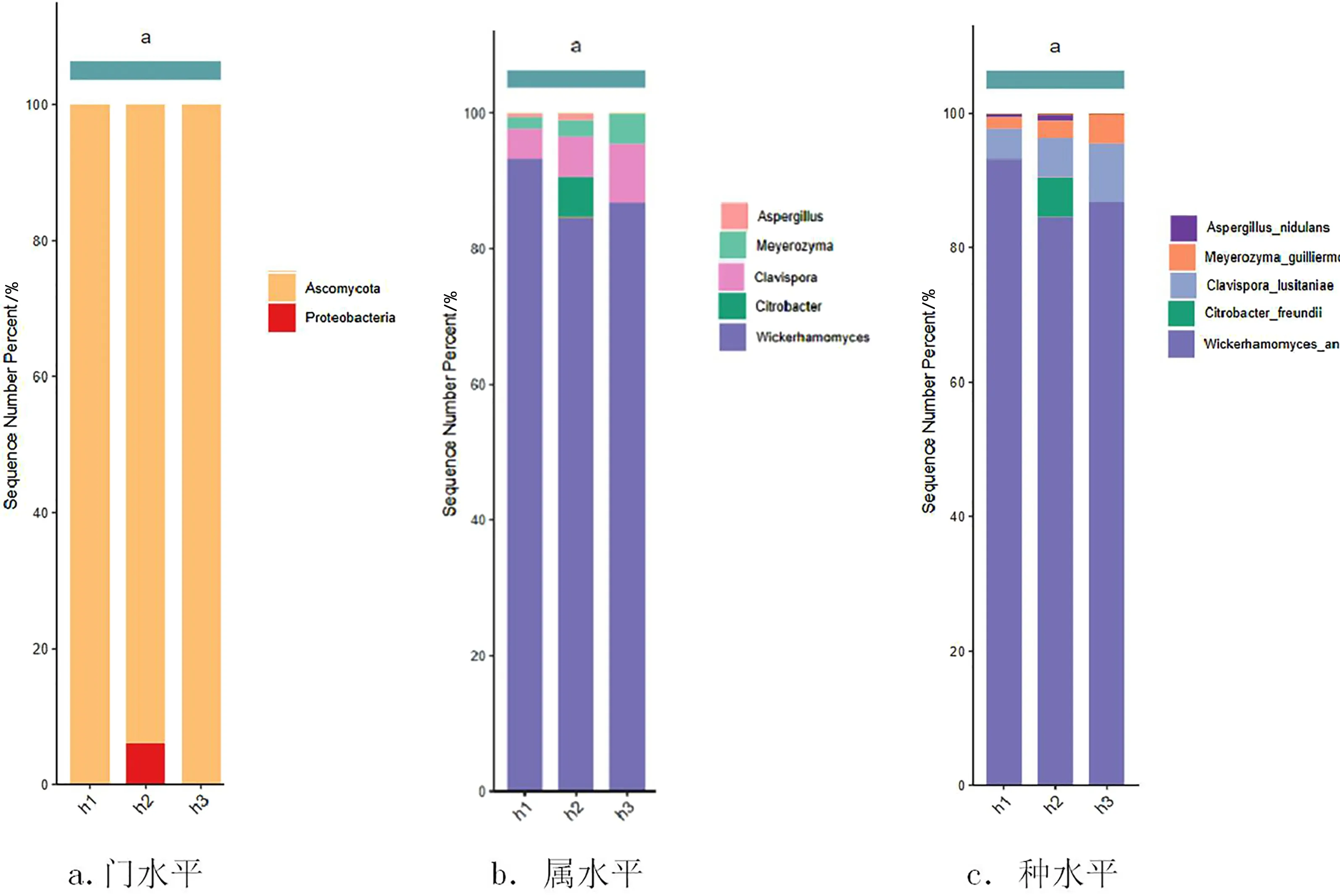
圖1 各個樣品在門、屬、種水平上的相對分布情況柱形圖注:相對豐度前20的物種。
2.2 紅豆杉共生菌主要代謝通路及紫杉醇合成的分析
對樣本的基因進行KEGG注釋,統(tǒng)計其可能參與或涉及的代謝途徑。由圖2可知,樣本的功能基因在Level1層級有6大類的代謝通路,分別為細胞過程(Cellular Processes)、環(huán)境信息處理(Environmental Information Processing)、遺傳信息處理(Genetic Information Processing)、人類疾病(Human Diseases)、新陳代謝(Metabolism)、生物體系統(tǒng)(Organismal Systems),其主要位于新陳代謝的代謝途徑中。主要功能基因二級代謝途徑有20類,其中富集差異表達基因最多的8個KEGG通路包括氨基酸代謝(Amino acid metabolism)、碳水化合物代謝(Carbohydrate metabolism)、輔因子和維生素的代謝(Metabolism of cofactors and vitamins)、信號轉導(Signal transduction)、翻譯(Translation)、折疊、分類和降解(Folding,sorting and degradation)、運輸和分解代謝(Transport and catabolism)、內分泌系統(tǒng)(Endocrine system)。
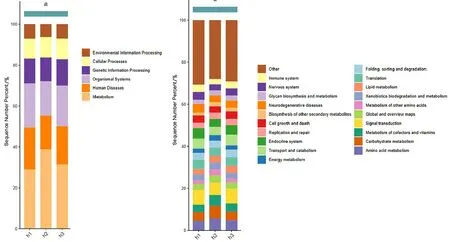
圖2 KEGG代謝通路的Level1和Level2層級豐度柱形圖
其中,氨基酸代謝和碳水化合物代謝最為活躍。在該樣本中檢測到了一些高豐度碳代謝基因,包括pgm(磷酸葡萄糖變位酶)、己糖激酶(HK)的基因、galM(醛糖-1-差向異構酶)等,共11個。糖酵解/糖異生途徑的特征基因明顯多于其他碳代謝途徑,這說明糖酵解和糖異生途徑在該樣本的碳循環(huán)過程中比較活躍,可以為生命活動提供能量[9]。除此之外,氨基酸代謝和碳水化合物代謝所產生的一些中間產物如乙酰輔酶A、丙酮酸、甘油醛-3-磷酸等與紫杉醇的合成相關。紫杉醇類化合物是通過異戊二烯前體合成的,萜類的異戊二烯結構單元可以通過位于細胞質中的甲羥戊酸途徑(MVA)或位于質體中的磷酸甲基赤蘚糖醇途徑(MEP)合成。MVA途徑起始于乙酰輔酶A,MEP途徑則起始于丙酮酸和甘油醛-3-磷酸,這2種途徑再經過共同的中間體IPP最終合成紫杉醇[5]。
通過查詢NCBI數據庫以及相關文獻[8,9]找到了合成紫杉醇的一些關鍵酶以及能分泌這些酶的微生物[10-12],如分泌去乙酰巴卡蛋白III-10β-O-乙酰轉移酶(Deacetyl Baccatin III-10β-O-acetyltransferase,DBAT)的菌種有Aspergillus candidus(念珠曲霉)、Fusarium solani(茄病鐮刀菌)等;分泌香葉酰香葉酰二磷酸合成酶(Geranylgeranyl diphosphate synthase,GGPPS)的菌種有Clavisporalusitaniae(葡萄牙棒孢酵母)、Aspergillus luchuensis(蘆竹曲霉)、Fusarium solani(茄病鐮刀菌)等;Fusarium solani (茄病鐮刀菌)也是能分泌紫杉二烯合酶(Taxadiene synthase,TS)的菌種。
上述菌種均屬于本實驗的優(yōu)勢菌門子囊菌門,其中包括排名第2的優(yōu)勢菌種Clavispora lusitaniae(葡萄牙棒孢酵母),以及排名第4的優(yōu)勢菌屬Aspergillus(曲霉屬)與該實驗結果相對應。這些優(yōu)勢菌種的存在為后續(xù)實驗菌種的篩選提供了理論基礎。
2.3 紅豆杉共生菌主要碳水化合物活性酶與紫杉醇活性的關系
碳水化合物活性酶數據庫(CAZy)共包括糖苷水解酶、糖基轉移酶、多糖裂合酶、碳水化合物酯酶、碳水化合物結合模塊、輔助模塊酶類6大類蛋白質家族。由CAZy功能注釋結果可知,GT48、GH3、GT62、GH85、GH32、GH5等6類碳水化合物活性酶的活性在樣本中較高,其中樣本中的特征碳水化合物活性酶為GH5(糖苷水解酶家族5)。
糖苷水解酶在寡糖合成、烷基糖苷和芳香基糖苷的合成、氨基酸和多肽的糖基化以及抗生素的糖基化方面發(fā)揮了重要作用。糖基轉移酶在生物體內催化活化的糖連接到不同的受體分子,如蛋白、核酸、寡糖、脂和小分子上[13]。糖基化是生物細胞中最重要的反應之一,這種天然的修飾存在于多種生物學活性不一樣的天然化合物中,包括抗生素、抗癌藥物、激素、生物堿以及黃酮等多種代謝產物[14]。該樣品中還檢測到了一些抗生素的合成代謝途徑,如圖3所示,新霉素和卡那霉素屬于氨基糖苷類抗生素,這類抗生素的生物合成途徑中,糖基化通常是修飾的最后一步,通過糖的位置、類型和數量的改變對抗生素的活性進行調節(jié);圖4中的新生霉素屬于糖苷類抗生素,其通常含有一個或多個糖基,研究表明,糖基可以維持、調節(jié)和改變糖苷類抗生素的生物活性,糖基化可以增加糖苷類抗生素的極性和水溶性,也可以增強某些抗生素的化學穩(wěn)定性[15,16]。糖苷水解酶的糖基化作用不僅可以改變抗生素的活性,還能改變紫杉醇的特性,在植物體內除了有游離狀態(tài)的紫杉醇還有糖基化的紫杉醇,紫杉醇的糖基化可以大大提高其水溶性,從而便于其在臨床水平上的應用。
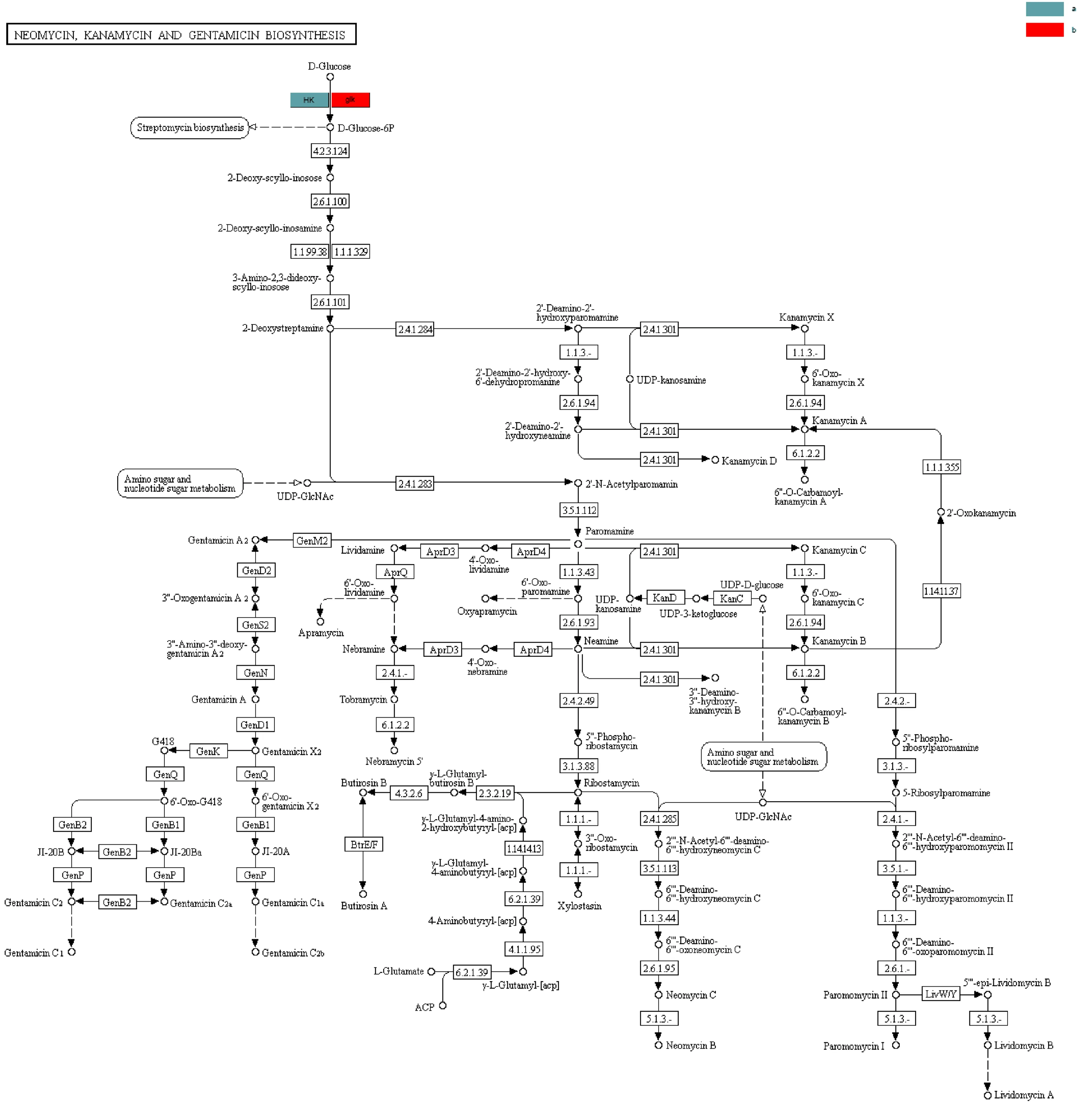
圖3 合成新霉素、卡那霉素和慶大霉素的相關途徑
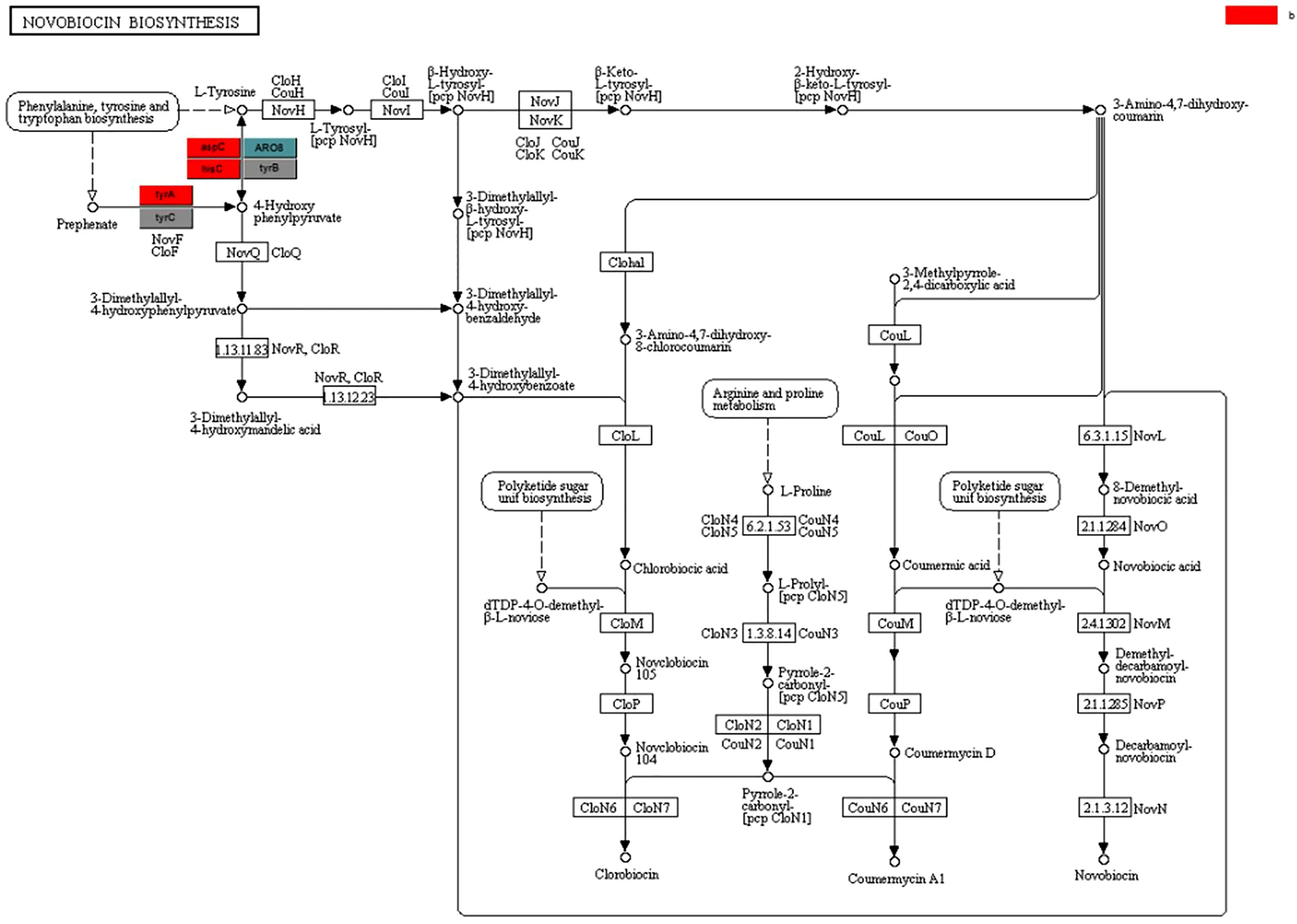
圖4 合成新生霉素的相關途徑
3 結論
紅豆杉是世界上公認的瀕臨滅絕的天然珍稀抗癌植物,其藥用價值主要體現在其提取物——次生代謝衍生物紫杉醇,故獲得合成紫杉醇的菌種,對縮短提取時間,提高紫杉醇的提取率具有重要意義。本實驗以紅豆杉葉為唯一碳源進行富集培養(yǎng),以確保富集到的菌群均為紅豆杉葉際共生菌,從而排除其他雜菌的干擾。后經宏基因組技術分析可得,本實驗所檢測到的菌種均處于紫杉醇的主要生產者——子囊菌門,且樣本中也存在合成紫杉醇的菌種Clavispora lusitaniae(葡萄牙棒孢酵母),故通過分析推測樣本中的菌種可以參與紫杉醇的合成代謝,并且其分泌的糖苷水解酶和糖苷轉移酶可以使紫杉醇發(fā)生糖基化修飾,從而提高其水溶性,便于其在臨床水平上的應用。
猜你喜歡
世界科學技術-中醫(yī)藥現代化(2022年2期)2022-05-25 13:17:14
昆明醫(yī)科大學學報(2021年12期)2021-12-30 07:00:10
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
實用口腔醫(yī)學雜志(2017年6期)2017-09-19 02:51:06
中外醫(yī)療(2016年15期)2016-12-01 04:25:50
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
哈爾濱醫(yī)藥(2015年2期)2015-12-01 03:57:41
中國衛(wèi)生(2014年12期)2014-11-12 13:12:52
