不同時程APP/PS1 模型小鼠嗅球病理和突觸形態(tài)變化及美金剛干預作用
2022-07-04 07:55:00劉佳妮劉劍剛羅增剛
實驗動物與比較醫(yī)學 2022年3期
劉佳妮, 劉劍剛, 韋 云, 羅增剛, 李 浩, 王 怡, 李 琨
(1. 中國中醫(yī)科學院西苑醫(yī)院, 中國中醫(yī)科學院老年病研究所, 北京 100091; 2. 中國中醫(yī)科學院研究生院, 北京 100700; 3. 天津中醫(yī)藥大學,天津 301617; 4. 北京市中醫(yī)管理局,北京 100053; 5. 北京開放大學,北京 100081)
阿爾茨海默病(Alzheimer's disease,AD)患者在淀粉樣蛋白-β(amyloid beta,Aβ)斑塊沉積和學習記憶障礙開始之前,往往會出現(xiàn)嗅覺功能障礙,其典型特征是識別氣味的能力下降[1]。淀粉樣前體蛋白(amyloid precursor protein,APP)/早老素1(presenilin 1,PS1)等雙轉基因模型AD 小鼠也表現(xiàn)出嗅覺受損[2]。嗅覺功能障礙被認為是AD的前驅癥狀,已被提議作為認知障礙的預測因素[3]。因此,針對嗅覺功能障礙的干預可能有助于延緩AD患者認知障礙病程的進展。然而目前嗅覺減退的神經(jīng)機制在很大程度上是未知的。由于通常不會對嗅覺結構進行神經(jīng)病理學分析[4],所以其臨床特征與潛在病理學之間的關系尚不清楚,但有研究者提出AD 患者嗅覺障礙可能是由嗅球(olfactory bulb)、嗅覺上皮或嗅皮質受損引起[5]。
嗅球作為第一個處理嗅覺信息[6]、最早積累Aβ的大腦結構,其病理變化已被證實可以顯示AD的早期神經(jīng)退行性變化,便于早期發(fā)現(xiàn)、早期干預[7]。已知嗅覺功能障礙癥狀會隨著年齡的增長而逐漸加重[8],同時伴隨Aβ的沉積和擴散;嗅球也隨著年齡的增長會對Aβ的影響變得更加敏感[9],更容易促進Aβ的積累和釋放[10]。
鹽酸美金剛(memantine hydrochloride,MEM)是首個經(jīng)FDA批準應用于治療中、重度AD的臨床藥物,具有調節(jié)谷氨酸活性的作用,屬于N-甲基-D-天冬氨酸受體拮抗劑的代表藥物[11]。多項研究表明,美金剛可以改善APPswe/PS1ΔE9 (APP/PS1)轉基因小鼠模型的學習記憶能力[12-13]。然而,美金剛對AD 嗅覺系統(tǒng)的作用報告并不多見,尚不清楚美金剛是否能改善APP/PS1小鼠模型的嗅球病理變化。
為了初步評估不同時程APP/PS1 雙轉基因模型小鼠的空間記憶能力和嗅球病理變化,以及美金剛的治療作用,本研究參考相關文獻報告[14],選取6 月齡(成年期)和12月齡(中年期)的APP/PS1小鼠,比較美金剛對不同月齡APP/PS1 小鼠嗅球病理和突觸形態(tài)的干預作用,為美金剛用于治療AD嗅覺障礙提供初步的實驗依據(jù)。
1 材料與方法
1.1 實驗動物
SPF 級4 周齡雄性APP/PS1 雙轉基因小鼠40 只,4周齡雄性C57BL/6 小鼠20 只,體質量均為13~15 g,由北京華阜康生物科技股份有限公司提供[SCXK(京)2019-0008],動物質量合格證號為11401300070455。小鼠飼養(yǎng)于中國中醫(yī)科學院西苑醫(yī)院屏障環(huán)境實驗動物室[SYXK (京) 2018-0018],室溫控制在24~26 ℃,相對濕度為40%~70%。小鼠自由進食和飲水,每日光照和黑暗各12 h,待飼養(yǎng)到3月齡或9月齡后進入分組實驗。實驗過程中嚴格遵循實驗動物3R 原則,按照國家科技部《關于善待實驗動物的指導性意見》和北京市《實驗動物福利倫理審查技術規(guī)范》的要求執(zhí)行。本研究方案經(jīng)過中國中醫(yī)科學院西苑醫(yī)院醫(yī)學倫理委員會審查通過(倫理審批號為2018XLC012-2)。
1.2 主要試劑與儀器
MEM 片(H.Lundbeck A/S 丹麥靈北制藥有限公司生產(chǎn),進口藥品注冊證號為H20120268、H20130372)由中國中醫(yī)科學院西苑醫(yī)院藥劑科提供;丙酮(批號20170411,進口分裝)由國藥集團化學試劑有限公司提供;Spon 812 包埋劑(批號ZB-S0060)購自美國SPI 公司;戊巴比妥鈉(批號T860901,進口分裝)由佛山市化工實驗廠提供;甲醛溶液(批號C10561782)購自上海麥克林生化科技有限公司。ZS-001 型Morris水迷宮系統(tǒng)由北京眾實迪創(chuàng)公司生產(chǎn);H-7650型透射電鏡由日立(日本)公司生產(chǎn);BX43型光學顯微鏡由日本奧林巴斯公司生產(chǎn);RM2245型半自動輪轉式切片機、ASP200S型全自動脫水機、EG11型自動包埋機和EM UC6超薄切片機均由德國Leica公司生產(chǎn)。
1.3 動物模型和分組
APP/PS1雙轉基因小鼠常規(guī)飼養(yǎng)至3月齡時,根據(jù)數(shù)字表法隨機分為6 月齡模型組(6-APP/PS1 組)、12月齡模型組(12-APP/PS1 組),以及使用藥物干預APP/PS1 雙轉基因小鼠的6 月齡MEM 組(6-MEM 組)、12 月齡MEM 組(12-MEM 組)。另設空白對照組,即常規(guī)飼養(yǎng)至3 月齡時的C57BL/6 小鼠分為6 月齡C57 組(6-C57組)、12月齡C57組(12-C57組)。以上每組各10只。空白組和模型組均以等體積純凈水灌胃,MEM組每日以MEM 2.6 mg/kg水溶液灌胃給藥。兩組小鼠分別在3月齡和飼養(yǎng)至9月齡時開始灌胃給藥,藥液體積為0.1~0.2 mL/10 g,均灌胃3 個月,分別至6 月齡和12月齡時進行水迷宮行為學檢測。
1.4 水迷宮法檢測小鼠行為學變化
采用Morris 水迷宮評估小鼠的空間學習記憶能力[15]。Morris 水迷宮由一個直徑1 m、高0.6 m的圓形水池組成,水位為35 cm,水溫保持在23 ℃。添加脫脂奶粉使池水變得不透明。逃生平臺位于水池的一個象限內,位于水面以下1 cm處。小鼠依次放置在其他3個不同象限的水池中,每只小鼠有90 s的時間爬上平臺,并允許在平臺上停留5 s,若小鼠找不到平臺則引導其上到平臺。通過每日4次、連續(xù)5 d的訓練,小鼠獲得了關于逃生平臺位置的空間記憶。第5 天訓練后24 h,移除平臺,再讓小鼠在池中游90 s,記錄和分析小鼠在原平臺位置的穿越次數(shù)(即穿臺次數(shù))、在目標平臺象限的停留時間和路程(即目標象限時間、目標象限路程),以及總路程。
1.5 HE染色觀察小鼠嗅球病理形態(tài)
在Morris水迷宮測試后次日,小鼠腹腔注射2%戊巴比妥鈉(30 mg/kg 體質量)深度麻醉,低溫下快速取出嗅球。每組隨機取5 只小鼠的嗅球組織,用質量分數(shù)為4%的多聚甲醛溶液固定,常溫下保存。組織浸泡后,包埋浸蠟,切片脫蠟,二甲苯浸洗3 次,每次15 min;100%、90%、80%和70%梯度乙醇脫苯,每次3 min;自來水沖洗3 min,去離子水漂洗3 min;蘇木精染液染色10 min后水洗,鹽酸乙醇分色,2 s后水洗;1% 伊紅染液染色1 min,去離子水稍水洗;100%、90%、80%和70%梯度乙醇脫水,每次1 min;二甲苯透明3次,每次15 min;中性樹膠封固后,每張切片分別于40 倍和200 倍光學顯微鏡下觀察嗅球病理組織形態(tài),并進行拍照,采用Image Pro Premier圖像分析系統(tǒng)進行分析。
1.6 超薄切片染色觀察小鼠嗅球超微形態(tài)
另5 只小鼠嗅球組織用2.5%戊二醛磷酸緩沖液固定,保存于4 ℃冰箱。使用振動切片機將嗅球冠狀切成厚度約0.2 mm的切片,然后用4%戊二醛固定2 h以上,再將嗅球組織切片暴露于1%鋨酸固定液2 h,用PBS 漂洗3 次(5 min/次),50%、70%、90%和100%梯度丙酮脫水(每次15 min),然后梯度浸透(丙酮和包埋劑比例依次為2∶1、1∶1、0∶1,各2 h),烤箱烘干(36~60 ℃,12~48 h)形成包埋塊。用超薄切片機行70 nm 切片,附于銅網(wǎng)上,超薄切片分別用醋酸鈾溶液和檸檬酸染液染色30 min,然后用透射電子顯微鏡觀察嗅球中神經(jīng)和突觸的超微結構改變。
1.7 統(tǒng)計學方法
采用SPSS 23.0軟件進行統(tǒng)計學分析。數(shù)據(jù)進行正態(tài)分布檢驗,符合正態(tài)分布的計量資料以±s表示。組間比較時采用單因素方差分析,方差齊時采用Tukey檢驗,方差不齊時采用Games-Howell 檢驗。不符合正態(tài)分布的計量資料以中位數(shù)(四分位數(shù)間距)表示,組間比較時使用非參數(shù)檢驗的Kruskal-WallisH檢驗。以P<0.05為差異有統(tǒng)計學意義。
2 結果
2.1 不同時程APP/PS1模型小鼠的空間記憶能力變化
連續(xù)5 d 訓練小鼠記住逃生平臺的位置后,對第6天小鼠空間探索實驗的穿臺次數(shù)、目標象限時間、目標象限路程和總路程進行分析。與6-C57 組比較,6-APP/PS1組的穿臺次數(shù)減少(P<0.05),目標象限時間和路程縮短(P<0.05)。與12-C57 組比較,12-APP/PS1組小鼠穿臺次數(shù)明顯減少(P<0.05),目標象限時間和路程顯著縮短(P<0.05)。與6-C57組比較,12-C57組小鼠穿臺次數(shù)減少(P<0.05),目標象限時間和路程縮短(P<0.05)。與6-APP/PS1 比較,經(jīng)過MEM(2.6 mg/kg)干預后,6-MEM 組穿臺次數(shù)顯著增加(P<0.05),目標象限時間和路程明顯延長(P<0.05)。和12-APP/PS1比較,12-MEM組穿臺次數(shù)增加(P<0.05),目標象限時間和路程明顯延長(P<0.05)。總路程上,6-C57 組和6-APP/PS1 組無明顯差異(P>0.05),而12-APP/PS1 組小鼠比12-C57 組顯著縮短(P<0.05),其余各組無顯著差異。實驗結果表明,美金剛治療改善了APP/PS1 小鼠的空間記憶能力。結果見圖1A。
在空間探索實驗中,C57 組小鼠通常采用趨向平臺式的搜索方式,而APP/PS1 組則多采用隨機式或邊緣式的搜索策略,提示APP/PS1 組小鼠空間記憶能力出現(xiàn)損傷;MEM 干預后的APP/PS1 小鼠顯示傾向使用趨向平臺式的搜索方式,說明MEM能提高APP/PS1小鼠的空間記憶能力。結果見圖1B。
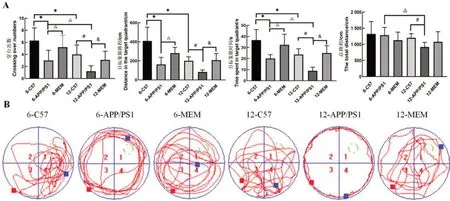
圖1 Morris水迷宮實驗評估各組小鼠的行為學變化Figure 1 Morris water maze test to evaluate the behavioral changes of mice in each group
2.2 不同時程APP/PS1 模型小鼠的嗅球病理變化
HE 染色后光學顯微鏡下觀察結果(圖2)顯示,6-C57組小鼠的嗅球在低倍鏡(×40)下呈卵圓形,結構完整清晰,內層可見大量神經(jīng)元呈環(huán)狀、層狀排布,規(guī)則緊密,細胞核呈圓形,自外向內依次可觀察到嗅神經(jīng)層、小球層(glomerular layer)、外叢狀層、僧帽細胞層、內叢狀層和顆粒細胞層;高倍鏡(×200)下可見小球層緊致,邊界清楚,呈球形排列,每個突觸小球周圍的球周細胞(periglomerular cells)分布密集。外叢狀層細胞深部可見僧帽細胞(mitral cells)胞體較大,胞質豐富。
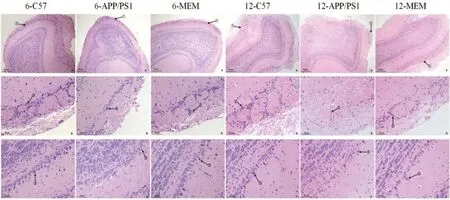
圖2 HE染色后光學顯微鏡觀察各組小鼠的嗅球病理變化Figure 2 Pathological changes of the olfactory bulb in mice in each group observed by optical microscopy after hematoxylin-eosin staining
與6-C57 組相比,6-APP/PS1 組低倍鏡(×40)下可見嗅球層狀結構完整清晰,突觸小球無明顯變化;高倍鏡(×200)下可見部分突觸小球周圍的球周細胞分布稀疏且數(shù)量稍減少,僧帽細胞萎縮且數(shù)量明顯減少。
與6-APP/PS1 組相比,6-MEM 組低倍鏡(×40)下可見嗅球形態(tài)較好,各細胞層結構較完整,小球層稍松散但均勻分布,呈球形結構,數(shù)量較多;高倍鏡(×200)下可見球周細胞數(shù)量稍增多,僧帽細胞數(shù)量則明顯增多。
與6-C57 組相比,12-C57 組低倍鏡(×40)下可見嗅球形態(tài)出現(xiàn)輕度改變,各細胞層結構尚完整,小球層部分欠均勻分布,邊界尚清楚;高倍鏡(×200)下可見球周細胞稍減少,僧帽細胞密度下降。
與12-C57 組相比,12-APP/PS1 組的部分嗅球層結構缺失,小球層松散且數(shù)量變少;高倍鏡(×200)下突觸小球數(shù)量減少伴有結構不清,周圍的球周細胞明顯減少,僧帽細胞數(shù)量大幅減少。
與12-APP/PS1 組相比,12-MEM 組低倍鏡(×40)下可見嗅球形態(tài)輕度變形,結構尚完整,小球層結構部分欠均勻分布,邊界尚清楚;高倍鏡(×200)下可見球周細胞和僧帽細胞數(shù)量稍增多。
統(tǒng)計分析各組小鼠嗅球中球周細胞和僧帽細胞的數(shù)量(圖3),結果發(fā)現(xiàn)APP/PS1 小鼠6 月齡時嗅球病理形態(tài)與C57BL/6 小鼠無顯著差異,僧帽細胞數(shù)量開始出現(xiàn)減少(P<0.05);12 月齡可觀察到嗅球的球周細胞和僧帽細胞數(shù)量均顯著減少(P<0.05)。與同齡的APP/PS1 小鼠相比,MEM 干預后,6 月齡時嗅球的僧帽細胞數(shù)量增加(P<0.05),而12 月齡時球周細胞及僧帽細胞數(shù)量無明顯增加(P>0.05)。結果說明,MEM有助于緩解嗅球病理學損傷,且在早期干預效果更加明顯。
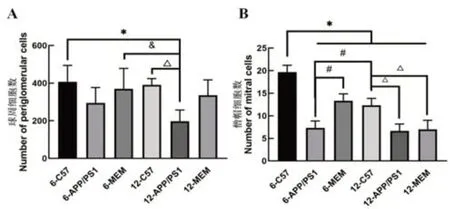
圖3 美金剛對球周細胞(A)和僧帽細胞(B)的影響Figure 3 Effects of memantine on periglomerular cells(A)and mitral cells(B)
2.3 不同時程APP/PS1 模型小鼠的嗅球神經(jīng)超微結構改變
超薄切片染色后透射電子顯微鏡觀察結果(圖4A)顯示,6-C57組小鼠嗅球組織神經(jīng)細胞形態(tài)正常,呈橢圓形,結構清晰;細胞膜完整;核仁明顯,大小基本正常;染色質分布較均勻,偶有染色質凝聚。6-APP/PS1 組小鼠嗅球神經(jīng)元形態(tài)尚規(guī)則,少量胞體及細胞器腫脹;細胞核形態(tài)有改變,出現(xiàn)分葉;核膜凹陷形成核裂,染色質濃縮邊集,部分核仁消失。6-MEM 組小鼠嗅球神經(jīng)元形態(tài)得到改善,形態(tài)較為規(guī)則,細胞膜偶有不連續(xù);細胞核較為完整,可見部分細胞存在核仁;染色質部分邊集,分布尚均勻。
12-C57 組小鼠嗅球組織神經(jīng)細胞形態(tài)基本正常,部分結構不清晰,細胞膜尚連續(xù),染色質出現(xiàn)邊集趨勢,細胞內脂褐素增多。12-APP/PS1組小鼠神經(jīng)細胞失去正常形態(tài),出現(xiàn)廣泛水腫、變性和壞死,形態(tài)不規(guī)則;細胞核形態(tài)明顯改變,染色質高度凝聚,細胞核固縮、溶解或核仁消失。12-MEM 組小鼠嗅球組織神經(jīng)細胞異常形態(tài)較多,胞體水腫,部分細胞核形態(tài)改變,染色質邊集。
隨著月齡增加,APP/PS1 小鼠的嗅球神經(jīng)元變性程度逐漸加重,可見嗅球神經(jīng)元形態(tài)不規(guī)則,尤其染色質會出現(xiàn)明顯的凝聚邊集現(xiàn)象。與APP/PS1 小鼠相比,MEM可使細胞形態(tài)更規(guī)則完整,染色質邊集情況一定程度減輕,且6-MEM 比12-MEM 改善更加明顯。結果說明,MEM 早期干預有助于改善APP/PS1 模型小鼠嗅球神經(jīng)元超微結構的損傷。
2.4 不同時程APP/PS1 模型小鼠的嗅球突觸超微結構改變
超薄切片染色后透射電子顯微鏡觀察結果(圖4B)顯示,6-C57 組小鼠嗅球組織突觸結構正常,可見大量對稱性突觸(抑制性突觸)和非對稱性突觸(興奮性突觸),突觸界面完整清晰,突觸膜后致密物(postsynaptic density,PSD)較厚,突觸前囊內有大量神經(jīng)遞質小泡,突觸小泡膜結構清晰正常。與6-C57組相比,6-APP/PS1 組突觸結構出現(xiàn)腫脹,突觸界面損傷,PSD 厚度減小,對稱性突觸數(shù)量減少,突觸小泡膜結構損傷。與6-APP/PS1 組相比,6-MEM 組突觸形態(tài)有一定改善,突觸界面較清晰,神經(jīng)遞質小泡較多,對稱性突觸數(shù)量增多。
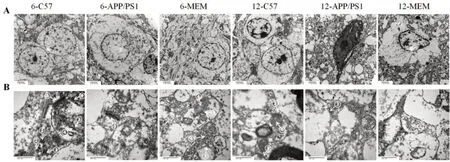
圖4 超薄切片染色后透射電子顯微鏡觀察各組小鼠嗅球神經(jīng)(A,×7 000)和突觸(B,×60 000)細胞超微結構Figure 4 Ultrastructure of the olfactory bulb nerve cells(A,×7 000)and synapses(B,×60 000)in each group observed by transmission electron microscope after ultrathin section staining
12-C57組小鼠嗅球組織突觸數(shù)量略有減少,非對稱性突觸膜結構欠清晰,突觸小泡數(shù)量減少。與12-C57 組相比,12-APP/PS1 組突觸結構損害加重,形態(tài)不規(guī)則,突觸體廣泛腫脹、變性,鏡下呈顆粒樣,突觸界面和突觸小泡無法分辨。與12-APP/PS1 組相比,12-MEM 組突觸改善不明顯,仍有部分腫脹,突觸界面不清晰,突觸數(shù)量及突觸小泡數(shù)量無明顯增加。其中12-APP/PS1組以及12-MEM組由于突觸體腫脹和鏡下顆粒樣變化,無法對突觸、囊泡數(shù)量和PSD 厚度進行準確計數(shù)以及長度測量。
結果顯示不同月齡的C57BL/6、APP/PS1小鼠以及MEM 干預后APP/PS1 小鼠的嗅球突觸超微結構改變,表明在衰老過程中,APP/PS1 小鼠顯示出比與其年齡匹配的C57BL/6 小鼠相對更顯著的突觸結構受損,而MEM可以改善這種突觸損害,且早期干預的改善效果更明顯。
3 討論
AD是一種以進行性記憶缺陷和認知能力下降為特征的神經(jīng)退行性疾病。針對AD的APP/PS1模型動物的認知缺陷在水迷宮測試中已經(jīng)得到了證實[16],APP/PS1 小鼠空間記憶受損最早可在6 月齡時出現(xiàn)[17],12月齡時認知和行為缺陷明顯[18],且記憶損害會在該模型的后續(xù)生命發(fā)展中持續(xù)存在[19]。本研究發(fā)現(xiàn),與6-C57 組相比,12-C57 組小鼠的空間記憶能力有顯著下降,與以往文獻報告[20]相符。在6月齡時APP/PS1轉基因小鼠的空間記憶障礙開始出現(xiàn),在12月齡時小鼠空間記憶功能嚴重受損。既往研究表明,用美金剛治療可以改善轉基因AD小鼠模型的學習記憶缺陷以及突觸神經(jīng)傳遞[21],本研究給予APP/PS1小鼠美金剛灌胃治療,證明美金剛可以恢復不同月齡APP/PS1 小鼠的空間記憶能力。此外,本研究初步闡述了美金剛可能對AD嗅覺結構改善有作用。
嗅覺減退是AD 常見的臨床前驅癥狀[22],超過90%的AD 患者都會存在嗅覺功能障礙[23]。嗅覺功能障礙伴隨著嗅覺結構的病理改變和細胞損傷,其中嗅球對AD病理學高度敏感,特別容易出現(xiàn)功能障礙,也是最早積累Aβ的大腦結構之一。Moriguchi等[24]研究顯示,小鼠嗅球切除會引起認知缺陷,并降低海馬中的膽堿能活性,說明嗅球與認知障礙密切相關。僧帽細胞是嗅球中最主要的投射神經(jīng)元,在嗅覺信息的傳遞和加工中具有重要作用,而球周細胞會對僧帽細胞的電活動進行相關調控。本實驗結果表明,6 月齡APP/PS1 小鼠嗅球內開始出現(xiàn)僧帽細胞數(shù)量下降,12月齡APP/PS1小鼠部分嗅球的神經(jīng)細胞形態(tài)萎縮變形,尤其球周細胞和僧帽細胞數(shù)量顯著減少。既往實驗證實,APP/PS1 小鼠在6~10 月齡時出現(xiàn)嚴重的嗅覺缺陷[25];結合本研究結果推測,這可能與嗅球神經(jīng)細胞萎縮變形以及球周細胞和僧帽細胞數(shù)量減少有關,而MEM 的早期干預可以改善APP/PS1 小鼠嗅球病理形態(tài)的變化,增加僧帽細胞數(shù)量,保護神經(jīng)元結構的完整。
嗅球具有多種類型的神經(jīng)遞質,其中谷氨酸和γ-氨基丁酸等是最常見最重要的遞質,這些神經(jīng)遞質的改變被認為是記憶障礙的主要原因之一[26]。僧帽細胞以谷氨酸作為神經(jīng)遞質[27],球周細胞以γ-氨基丁酸和多巴胺作為神經(jīng)遞質[28],其中谷氨酸是中樞神經(jīng)系統(tǒng)中一種重要的興奮性神經(jīng)遞質,通過特異性受體發(fā)揮作用,例如N-甲基-D-天冬氨酸受體。谷氨酸興奮性毒性會引發(fā)神經(jīng)元持續(xù)性去極化,造成神經(jīng)功能異常,是AD進展的重要原因之一[29]。而MEM屬于非競爭性N-甲基-D-天冬氨酸受體拮抗劑,可以阻斷谷氨酸濃度病理性升高導致的神經(jīng)元損傷[30],還可通過N-甲基-D-天冬氨酸受體減少不溶性Aβ含量,然后恢復AD動物模型中的谷氨酸穩(wěn)態(tài)[31]。筆者推測MEM干預后嗅球內病理形態(tài)改善、僧帽細胞數(shù)量增加可能與其減輕了嗅球中Aβ的積累、減輕谷氨酸誘導的僧帽細胞損傷有關。
突觸損傷是AD 病理生理學中的早期和關鍵事件,突觸變化的可能后果就是學習和記憶功能受損[32]。與Aβ斑塊、神經(jīng)原纖維纏結或神經(jīng)元丟失相比,突觸損傷與AD患者中觀察到的認知缺陷更相關[33]。目前AD突觸研究大量集中在海馬[34]、皮質等領域,極少涉及嗅球。有研究表明,6~7 月齡之后的APP/PS1 轉基因AD模型小鼠嗅球的樹突棘密度顯著降低,突觸小泡破損,PSD 厚度降低[35]。本研究發(fā)現(xiàn),6 月齡APP/PS1小鼠突觸結構出現(xiàn)腫脹,12 月齡小鼠的突觸結構損傷更為嚴重,突觸界面無法分辨,與以往文獻報告結果相同。MEM 干預3 個月后可緩解APP/PS1 小鼠的突觸損傷,早期(3月齡)較晚期(9月齡)時進行干預的改善效果更明顯。異常的神經(jīng)網(wǎng)絡興奮性是AD的一個普遍特征。Palop等[36]研究表明,中等水平的Aβ可以增強突觸前活動及突觸傳遞,而高水平的Aβ會降低谷氨酸能突觸傳遞并最終導致突觸丟失。另外,Wesson等[37]認為嗅球的早期神經(jīng)元過度興奮可能增強Aβ的突觸后釋放。因此,異常的神經(jīng)網(wǎng)絡興奮性與Aβ的產(chǎn)生、釋放之間形成了惡性循環(huán),這種惡性循環(huán)可能加速AD 的發(fā)病。筆者推測美金剛在Aβ 低濃度階段的早期(3月齡)干預降低了嗅球神經(jīng)元的過度興奮及其對Aβ的積累和傳遞,從而改善了6-APP/PS1組小鼠的突觸結構。
認知障礙主要是由與年齡相關的神經(jīng)病變累積驅動[38],同時自然老化過程也是影響嗅覺功能的主要因素之一[39]。有研究認為,嗅覺功能障礙已被確定為老年人5 年死亡率的最佳預測指標[40]。本研究觀察到,與6-C57 相比,12-C57 的僧帽細胞密度下降,突觸小泡數(shù)量減少,說明隨月齡增加,C57BL/6 小鼠不僅會出現(xiàn)空間記憶能力的下降,嗅球也會出現(xiàn)超微結構的早期改變,提示老化是影響認知功能和嗅覺病理的重要因素。研究顯示,APP/PS1轉基因小鼠嗅球中Aβ負荷呈年齡依賴性增加[41],而嗅覺系統(tǒng)中Aβ 沉積物會促進嚙齒類動物的AD 病理學改變[42]。本實驗發(fā)現(xiàn),與12-C57 相比,12-APP/PS1 小鼠的空間記憶能力有顯著下降,球周細胞、僧帽細胞數(shù)量大幅減少,突觸結構損害加重,突觸體廣泛腫脹、變性,說明APP/PS1 小鼠在衰老過程中比正常同齡小鼠更容易出現(xiàn)認知功能障礙和嗅球病理改變,而嗅覺系統(tǒng)的顯著病理改變可能是加重空間學習記憶障礙的重要原因。
美金剛可以減少嗅球神經(jīng)元的損傷,增加僧帽細胞數(shù)量,改善突觸結構。根據(jù)既往研究推測,美金剛可能通過阻斷谷氨酸濃度的升高、減少AD早期嗅球神經(jīng)元過度活躍,減少腦內Aβ的產(chǎn)生和傳播,從而改善空間記憶能力。未來本研究組將重點研究AD模型小鼠空間記憶與嗅球內部神經(jīng)震蕩、神經(jīng)元活動的相關性,以此探討美金剛在AD 嗅覺障礙臨床應用中的潛在機制。此外,AD嗅覺缺陷是由于中樞還是外周嗅覺神經(jīng)系統(tǒng)的功能障礙所引起,目前這一問題仍不明確。大腦皮層和邊緣系統(tǒng)都有可能是AD嗅覺障礙的原因。美金剛對嗅覺投射的大腦皮層(顳葉)、邊緣系統(tǒng)(杏仁核、海馬和下丘腦)等其他嗅覺系統(tǒng)及認知功能的影響,也需要更多的基礎實驗和臨床試驗來明確。
綜上所述,本實驗初步觀察了不同時程APP/PS1雙轉基因小鼠嗅球病理和超微結構的變化,發(fā)現(xiàn)美金剛早期干預可能是改善AD 嗅球病理改變的有效方法,但這一結果還需要進一步的研究加以證實。
【醫(yī)學倫理聲明Medical Ethics Statement】
本研究涉及的所有動物實驗均已通過中國中醫(yī)科學院西苑醫(yī)院醫(yī)學倫理委員會審查批準(No.2018XLC012-2)。所有實驗過程均遵照實驗動物相關法律法規(guī)條例要求進行。
All experimental animal protocols in this study were reviewed and approved by Medical Ethics Committee of Xiyuan Hospital, China Academy of Chinese Medical Sciences (Approval Letter No. 2018XLC012-2), and all experimental protocols were carried out following the guidelines such asAnimal Management Regulations(01/03/2017),Laboratory Animal: Guideline for Ethical Review of Animal Welfare(GB/T 35892—2018), ARRIVE 2.0, IGP 2012 and IAVE 2010.
[作者貢獻Author Contribution]
劉佳妮負責具體的動物實驗操作、數(shù)據(jù)統(tǒng)計與分析、文章撰寫及修改;劉劍剛和韋云負責研究思路設計、實驗課題申報、實驗方案制定、部分實驗操作、倫理委員會審批程序并提供文件信息,以及修改文章;羅增剛、李浩和王怡提供技術援助并負責論文審核及把關;李琨收集資料并輔助數(shù)據(jù)分析與核對。
[利益聲明Declaration of Interest]
所有作者均聲明本文不存在利益沖突。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學苑創(chuàng)造·A版(2020年10期)2020-11-06 05:21:26
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
作文周刊·小學一年級版(2016年27期)2017-06-03 23:21:17
絲綢之路(2016年9期)2016-05-14 14:36:33
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
海外文摘(2016年4期)2016-04-15 22:28:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
