不同育肥海區(qū)香港牡蠣的營養(yǎng)評價及基因表達相關性分析
2022-07-08 13:54:46嚴雪瑜覃波忠黃偉德彭金霞蔣美君李選積鐘方杰韋嬪媛
食品工業(yè)科技 2022年13期
嚴雪瑜,覃波忠,黃偉德,彭金霞,朱 鵬,蔣美君,張 斌,李選積,鐘方杰,,韋嬪媛,
(1.廣西北部灣海洋生物多樣性養(yǎng)護重點實驗室,北部灣大學海洋學院,廣西欽州 535011;2.欽州市水產技術推廣站,廣西欽州 535000;3.廣西水產遺傳育種與健康養(yǎng)殖重點實驗室,廣西壯族自治區(qū)水產科學研究院,廣西南寧 530021)
香港牡蠣(Crassostrea hongkongensis),俗稱生蠔或大蠔,隸屬雙殼綱(Bivalvia)牡蠣科(Ostreidae),其肉質鮮美、營養(yǎng)豐富,干肉中含有蛋白質45%~57%、脂肪7%~11%、糖元20%~40%,還富含多種維生素、必需氨基酸、牛磺酸以及鈣、鋅、硒等微量礦物元素,具有較高的營養(yǎng)價值[1]。香港牡蠣屬于暖水性近岸海域灘涂生長品種,適宜鹽度范圍為5‰~25‰,主要分布于我國南方沿海地區(qū),是廣西貝類養(yǎng)殖的主導品種[2]。目前,廣西香港牡蠣成品蠔的養(yǎng)殖方式采取了每年秋、冬季節(jié)(10~11 月份)在近岸或灣外開放性海域投放大規(guī)格牡蠣苗,進行異地養(yǎng)成和育肥養(yǎng)殖的方法,從而快速獲得滿足上市規(guī)格的肥蠔[3]。相關研究表明,牡蠣主要營養(yǎng)成分含量會隨著生長時期的不同而呈現季節(jié)性變化[4],生長海域的水質、鹽度、溫度、餌料種類和營養(yǎng)組成等因素都會對香港牡蠣的生化組成和品質產生一定的影響[5?7]。目前,關于香港牡蠣營養(yǎng)品質的分析雖有報道,但針對不同育肥養(yǎng)殖海區(qū)中育肥期香港牡蠣的營養(yǎng)組成和相關代謝基因的表達差異以及兩者之間的相關性等進行系統(tǒng)比較分析尚未有深入研究。
廣西欽州是我國著名的“大蠔之鄉(xiāng)”,欽州灣水質優(yōu)良,餌料豐富,是適宜香港牡蠣繁育生長的天然牧場,也是香港牡蠣的主產區(qū)及苗種供應地。欽州灣的牡蠣育肥養(yǎng)殖海區(qū)主要分布在大風江和欽州港海域,均屬于海河口交界處,具有海水鹽度適中,餌料豐富等特點。因此,本文以育肥期的香港牡蠣為研究對象,對廣西欽州灣海域的大風江和欽州港兩個育肥養(yǎng)殖海區(qū)的香港牡蠣主要營養(yǎng)成分、礦物元素、氨基酸含量和營養(yǎng)代謝相關基因的表達情況進行比較和相關性分析,為了解不同育肥養(yǎng)殖海區(qū)香港牡蠣的營養(yǎng)結構組成和相關遺傳背景,以及全面評價育肥期香港牡蠣的營養(yǎng)價值及營養(yǎng)評價候選基因的篩選奠定理論基礎,從而進一步為香港牡蠣深加工及資源合理開發(fā)利用提供科學依據。
1 材料與方法
1.1 材料與儀器
實驗所用香港牡蠣樣品 分別采集于廣西欽州灣大風江牡蠣育肥養(yǎng)殖海區(qū)(簡稱大風江養(yǎng)殖區(qū),N 21°38′24.10″,E 108°52′27.91″)和欽州港牡蠣育肥養(yǎng)殖海區(qū)(簡稱欽州港養(yǎng)殖區(qū),N 21°44′22.31″,E 108°34′4.28″),規(guī)格大小為:殼高10~15 cm、殼長6~8 cm、殼寬4~6 cm、軟體組織重量30~40 g;石油醚、鹽酸、硼酸、氫氧化鈉、硫酸鉀、硝酸銀 分析純,天津大茂化學試劑廠;TransZol RNA提取試劑盒、Trans Script All-in-one First-Strand cDNA Synthesis Super Mix for qPCR 試劑盒、PerfectStartTM Green qPCR SuperMix 試劑盒 北京全式金生物公司。
GZX-GF101-3-BS 電熱恒溫鼓風干燥箱 上海躍進醫(yī)療器械有限公司;B180 馬弗爐 德國納博熱公司;消化爐 上海洪紀儀器設備有限公司;KDN-815 定氮儀、SZC101S1 脂肪測定儀 上海纖檢儀器有限公司;HITACHI 高速冷凍離心機 日本日立公司;Light Cycler 96 實時熒光PCR 儀 瑞士羅氏公司。
1.2 實驗方法
1.2.1 樣品的處理 分別于大風江養(yǎng)殖區(qū)和欽州港養(yǎng)殖區(qū)采集30 個育肥期(11~12 月份)的香港牡蠣樣品,所有樣品均活體開殼取出全部軟體組織,每組隨機選取6 個牡蠣個體采集新鮮的外套膜、閉殼肌和鰓等組織樣品,置于液氮中冷凍后于?80 ℃的條件下保存待測;其余牡蠣樣品均取其整體的軟體組織,進行組內樣品混合和勻漿后于?20 ℃的條件下保存待測。
1.2.2 指標的測定
1.2.2.1 常規(guī)營養(yǎng)成分測定 香港牡蠣組織中的常規(guī)營養(yǎng)成分含量參照食品安全國家標準進行測定,其中能量參照GB 28050-2011(修訂版)進行測定;蛋白質參照GB 5009.5-2016(第一法)進行測定;脂肪參照GB 5009.6-2016(第二法)進行測定;氨基酸組分和含量參照GB 5009.124-2016 進行測定;膽固醇參照GB 5009.128-2016(第二法)進行測定;維生素A 和維生素E 參照GB 5009.82-2016(第一法)進行測定。
1.2.2.2 礦物元素測定 香港牡蠣組織中的鈣(Ca)、鋅(Zn)、鈉(Na)、鎂(Mg)等礦物元素含量均參照食品安全國家標準GB 5009.82-2016(第二法)進行測定。
1.2.2.3 氨基酸測定與營養(yǎng)價值評定分析 香港牡蠣組織中的氨基酸組分和質量分析參照GB 5009.124-2016 進行測定和分析;根據1973 年FAO/WHO 提出的氨基酸計分模式和中國預防醫(yī)學科學院營養(yǎng)與食品衛(wèi)生研究所提出的雞蛋蛋白模式進行比較,并計算氨基酸評分(amino acid score,AAS)和化學評分(chemical score,CS)[8]。
AAS 和CS 的計算公式分別為:
AAS=被測蛋白質氨基酸含量(mg/g N)/FAO/WHO 評分標準模式中相應氨基酸含量(mg/g N);
CS=被測蛋白質氨基酸含量(mg/g N)/雞蛋蛋白質中相應氨基酸含量(mg/g N)。
1.2.3 香港牡蠣組織相關基因的表達分析
1.2.3.1 總RNA 提取和cDNA 合成 參照Trizol 法[9]利用TransZol RNA 提取試劑盒進行組織總RNA 的提取,并根據RNA 反轉錄試劑盒進行cDNA 第1 鏈的合成。
1.2.3.2 引物設計和實時熒光定量聚合酶鏈式反應(PCR)根據美國國家生物信息中心(NCBI)上公布的牡蠣基因序列和相關參考文獻[10?12],設計并合成熒光定量PCR 引物,如表1 所示。實時熒光定量PCR的反應體系(20 μL)為:上、下游引物各0.5 μL,cDNA模板2 μL,2×TransStart Tip Green qPCR SuperMix反應液10 μL,無核酸酶水7 μL;反應程序為:94 ℃預變性30 s,94 ℃ 5 s、58 ℃ 15 s、72 ℃ 10 s 擴增40 個循環(huán);以翻譯延伸因子1A(EF1A)為內參基因,采用2?ΔΔCt計算目的基因的相對表達量。

表1 實時熒光定量PCR 引物Table 1 The primer sequences for quantitative RT-PCR
1.2.4 香港牡蠣代謝基因表達和營養(yǎng)成分相關性分析 利用SPSS 軟件對香港牡蠣代謝基因的相對表達量和營養(yǎng)成分的數據進行斯皮爾曼(Spearman)相關性分析,P<0.05 表示差異顯著,P<0.01 表示差異極顯著。
1.3 數據處理
采用Excel、SPSS、GraphPad Prism 5 和Omic-Share Tools 等軟件或在線工具進行數據統(tǒng)計分析和圖片處理,數據結果以“平均值±標準誤(Mean±SEM)”表示,顯著性分析采用配對樣品T檢驗,P<0.05 表示差異顯著,P<0.01 表示差異極顯著。
2 結果與分析
2.1 不同育肥養(yǎng)殖海區(qū)香港牡蠣常規(guī)營養(yǎng)成分和礦物元素含量分析
研究顯示,牡蠣軟體組織中糖原具有呈味作用[13],是影響牡蠣風味特征的重要因子[14],而水分含量則會影響牡蠣肉的濕度和多汁性,脂肪含量的變化也會賦予牡蠣不同的堅實度和口感[15]。由表2 可知,欽州灣兩個養(yǎng)殖海區(qū)育肥期的香港牡蠣在能量、蛋白質、脂肪、氨基酸、膽固醇和維生素等營養(yǎng)成分含量均存在不同程度的差異;其中,欽州港養(yǎng)殖區(qū)香港牡蠣的能量、蛋白質、脂肪、氨基酸、膽固醇、維生素A 和維生素E 等成分的含量均高于大風江養(yǎng)殖區(qū),且膽固醇、維生素A 和維生素E 的含量組間差異顯著(P<0.05),這3 種成分的含量分別是大風江養(yǎng)殖區(qū)的1.9、1.7 和1.6 倍;表明欽州灣不同育肥海區(qū)養(yǎng)殖的香港牡蠣的常規(guī)營養(yǎng)成分含量存在一定的差異,這可能與不同海域的海水理化環(huán)境存在差異有關。因為牡蠣屬于變滲透壓動物,需要調整其生理代謝和能量分配來適應不同的鹽度和環(huán)境的變化,從而影響了自身的營養(yǎng)成分結構和含量[16]。本試驗結果與丁丹勇等[17]的研究相似,該研究認為欽州、湛江東海島、湛江官渡、陽西養(yǎng)殖區(qū)香港牡蠣的營養(yǎng)成分存在顯著差異,這可能與不同養(yǎng)殖區(qū)海水的鹽度差異有關。
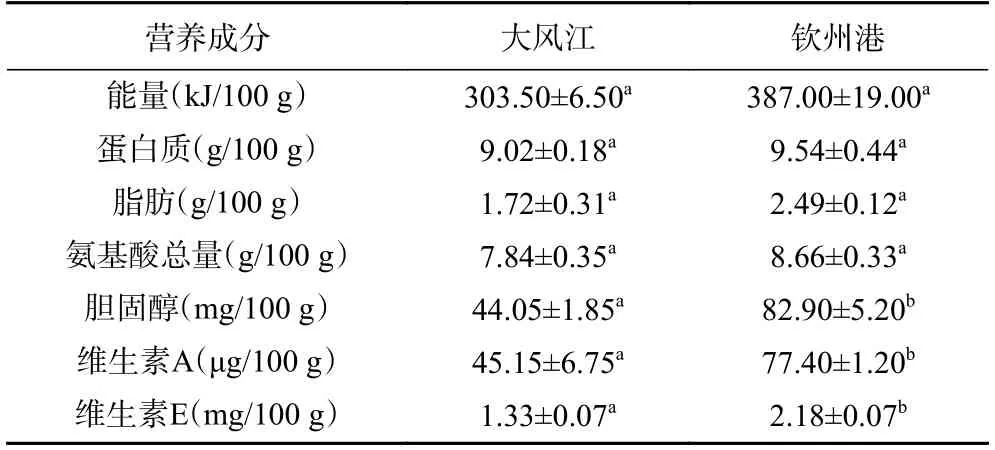
表2 欽州灣不同養(yǎng)殖區(qū)香港牡蠣常規(guī)營養(yǎng)成分含量Table 2 The main nutritional composition contents of C.hongkongensis from different culture areas
研究表明,牡蠣中含有豐富的鈣、鐵、鋅、硒、鉀、鈉、鎂等人體生理功能重要的礦物質元素[18?20];其中,牡蠣組織中的微量元素鋅和硒是海產品含量之首[5],鋅對人體的免疫功能起著調節(jié)作用,具有“生命之花”的美稱[21];牡蠣還因其富含鈣和鋅等礦物質元素而被稱為“海中牛奶”[22]。由表3 可知,大風江養(yǎng)殖區(qū)香港牡蠣的鈣、鈉和鎂等礦物元素的含量均顯著高于欽州港養(yǎng)殖區(qū)(P<0.05),分別是欽州港養(yǎng)殖區(qū)的2.4、2.2 和1.6 倍;而欽州港養(yǎng)殖區(qū)香港牡蠣的鋅元素含量則顯著高于大風江養(yǎng)殖區(qū)(P<0.05),為大風江養(yǎng)殖區(qū)的1.7 倍。這可能是由于牡蠣對不同元素的累積與其所在海域的礦物元素含量存在著一定的差異有關,且部分元素間的累積存在相互競爭或抑制作用[23]。林海生等[14]等研究也認為牡蠣中金屬元素的含量具有一定的地域差異,其原因與牡蠣品種、生長周期及其生長海域環(huán)境中的礦物元素種類有關。

表3 欽州灣不同養(yǎng)殖區(qū)香港牡蠣主要礦物元素含量Table 3 The main mineral element contents of C.hongkongensis from different culture areas
2.2 不同育肥養(yǎng)殖海區(qū)香港牡蠣氨基酸組成及營養(yǎng)價值分析
不同育肥養(yǎng)殖海區(qū)香港牡蠣氨基酸組成及含量如表4 所示,除了色氨酸被酸水解外,欽州灣兩個養(yǎng)殖區(qū)香港牡蠣組織中檢測出了17 種常見氨基酸,包括7 種必需氨基酸、2 種半必需氨基酸和8 種非必需氨基酸,氨基酸總量為7.84~8.66 g/100 g,其中谷氨酸的含量最高,胱氨酸的含量最低。在兩個養(yǎng)殖區(qū)香港牡蠣的氨基酸組成比較中,欽州港養(yǎng)殖區(qū)香港牡蠣的苯丙氨酸、組氨酸、谷氨酸和丙氨酸等氨基酸含量顯著高于大風江養(yǎng)殖區(qū)(P<0.05),其它氨基酸含量差異不顯著。氨基酸是牡蠣的一種重要滲透壓效應物,牡蠣機體可通過氨基酸的代謝來調節(jié)體內的滲透壓,因此不同海區(qū)的牡蠣也會因其生長環(huán)境的不同而在氨基酸的組成和含量方面存在差異[24]。
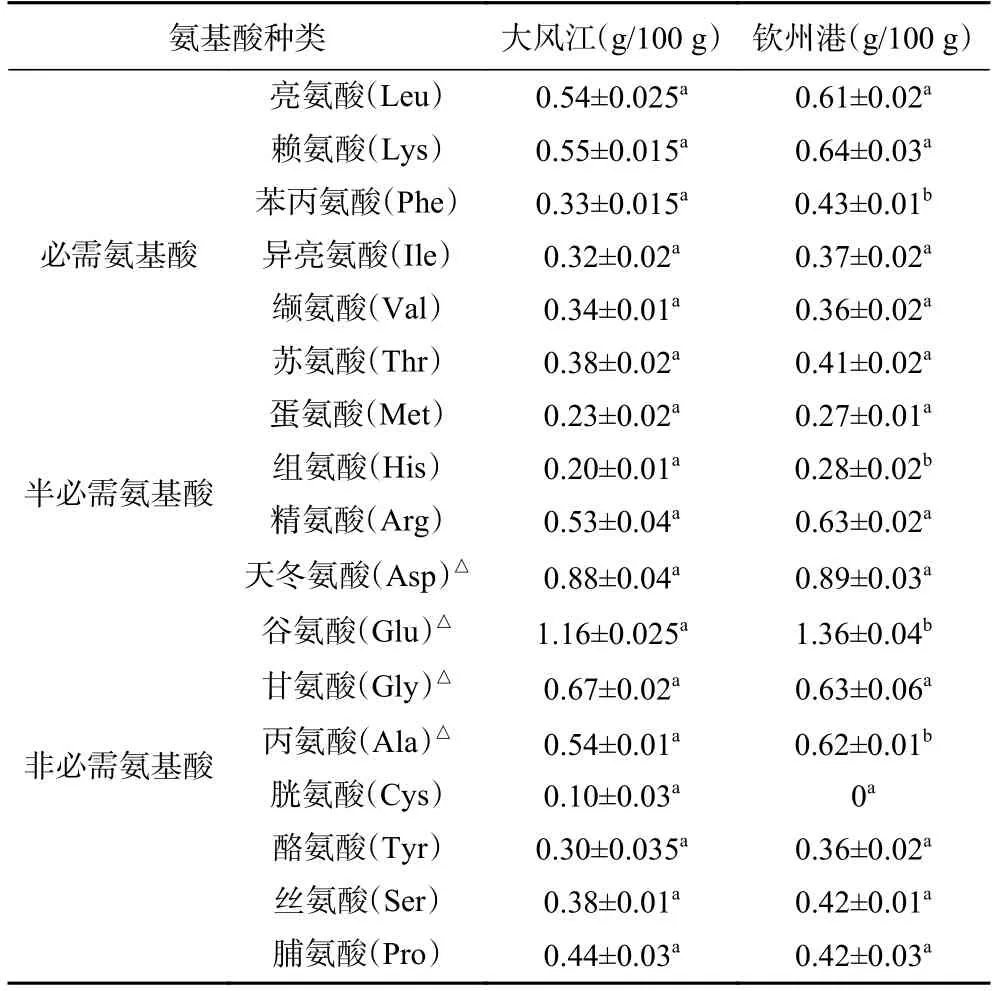
表4 欽州灣不同養(yǎng)殖區(qū)香港牡蠣氨基酸組成及含量Table 4 Amino acid composition and contents of C.hongkongensis from different culture areas
牡蠣蛋白質的營養(yǎng)價值主要取決于其氨基酸種類的齊全程度和必需氨基酸含量的高低[20]。由表5可知,欽州港養(yǎng)殖區(qū)香港牡蠣的必需氨基酸含量為3.08 g/100 g,占氨基酸總量的35.51%,與WHO/FAO模式推薦的模式(35.38%)接近[25];而大風江養(yǎng)殖區(qū)香港牡蠣的必需氨基酸含量和比例分別為2.67 g/100 g 和34.06%,稍低于欽州港養(yǎng)殖區(qū)。欽州港養(yǎng)殖區(qū)香港牡蠣半必需氨基酸為0.91 g/100 g(10.51%),同樣高于大風江養(yǎng)殖區(qū)(0.73 g/100 g,9.31%)。
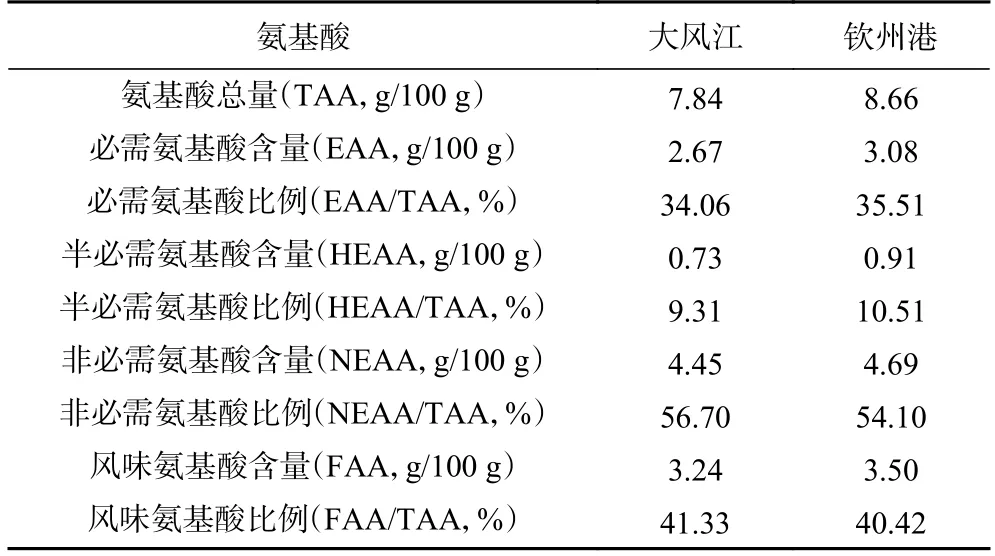
表5 欽州灣不同養(yǎng)殖區(qū)香港牡蠣氨基酸組成評價Table 5 Evaluation of amino acid composition in C.hongkongensis from different culture areas
為進一步評價香港牡蠣蛋白質營養(yǎng)價值的高低,采用FAO/WHO 評分模式和雞蛋蛋白的必需氨基酸含量作標準計算,欽州灣不同養(yǎng)殖區(qū)香港牡蠣的氨基酸評分(AAS)和化學評分(CS)結果詳見表6。根據必需氨基酸組成評價,大風江養(yǎng)殖區(qū)香港牡蠣纈氨酸的AAS 和CS 評分最低,為第一限制氨基酸;欽州港養(yǎng)殖區(qū)香港牡蠣的第一限制氨基酸則為纈氨酸(AAS)和蛋氨酸+胱氨酸(CS)。根據AAS 評分,欽州港養(yǎng)殖區(qū)香港牡蠣的異亮氨酸、亮氨酸、賴氨酸、苯丙氨酸+酪氨酸、蘇氨酸的AAS 評分均高于大風江養(yǎng)殖區(qū),其中AAS 評分最高的是苯丙氨酸+酪氨酸(1.35);根據CS 評分,欽州港養(yǎng)殖區(qū)香港牡蠣的異亮氨酸、亮氨酸、賴氨酸、苯丙氨酸+酪氨酸、蘇氨酸的CS 評分也高于大風江養(yǎng)殖區(qū),其中CS 評分最高的是賴氨酸(0.94)。因此,欽州港養(yǎng)殖區(qū)香港牡蠣的氨基酸平衡效果相對更好,含量豐富且均勻,蛋白質質量好,均屬于優(yōu)質的蛋白質,具有較高的營養(yǎng)價值。

表6 欽州灣不同養(yǎng)殖區(qū)香港牡蠣必需氨基酸評分Table 6 Scores of essential amino acids of C.hongkongensis from different culture areas
牡蠣的鮮美口感還在一定程度上取決于谷氨酸、天冬氨酸、甘氨酸和丙氨酸等4 種風味氨基酸的組成和含量,其中谷氨酸和天冬氨酸是呈鮮味的特征氨基酸,而甘氨酸和丙氨酸則是呈甘味的特征氨基酸[17,26]。欽州灣兩個養(yǎng)殖區(qū)香港牡蠣的風味氨基酸的總含量為3.24~3.50 g/100 g,欽州港養(yǎng)殖區(qū)香港牡蠣的天冬氨酸、谷氨酸和丙氨酸的含量以及風味氨基酸總量(3.50 g/100 g)均高于大風江養(yǎng)殖區(qū)(3.24 g/100 g),但風味氨基酸占氨基酸總量的比例(40.42%)卻低于大風江養(yǎng)殖區(qū)(41.33%),表明大風江養(yǎng)殖區(qū)香港牡蠣的鮮美程度相對較高。黃艷球等[27]對比了廣西欽州、廣東陽江、廣東湛江和廣東汕頭四個養(yǎng)殖區(qū)的香港牡蠣的氨基酸組成和含量,也發(fā)現欽州大蠔在氨基酸總量、必需氨基酸、非必需氨基酸含量以及在風味氨基酸天冬氨酸、谷氨酸、甘氨酸、丙氨酸含量方面都具有明顯的優(yōu)勢。
2.3 不同育肥養(yǎng)殖海區(qū)香港牡蠣營養(yǎng)代謝相關基因組織表達分析
由圖1 可知,δ-1 吡咯啉-5-羧酸合成酶(delta-1-pyrroline-5-carboxylate synthase,P5CS)、糖原磷酸酶(glycogen phosphorylase,GP)、雙特異性酪氨酸磷酸化調節(jié)激酶(dual-specificity tyrosine-phosphorylation-regulated kinase,DYRK)和膽固醇調節(jié)元件結合蛋白(sterol-regulatory element binding protein,SREBP)等四個營養(yǎng)代謝相關基因的相對表達量在香港牡蠣外套膜、閉殼肌和鰓等不同組織的組間表達趨勢完全一致,均表現為在同組不同組織內整體呈上調或下調趨勢;其中,欽州港養(yǎng)殖區(qū)香港牡蠣的P5CS、GP和SREBP的基因相對表達量高于大風江組,而DYRK基因的組織表達結果則低于大風江養(yǎng)殖區(qū),但組間差異均不顯著(P>0.05)。
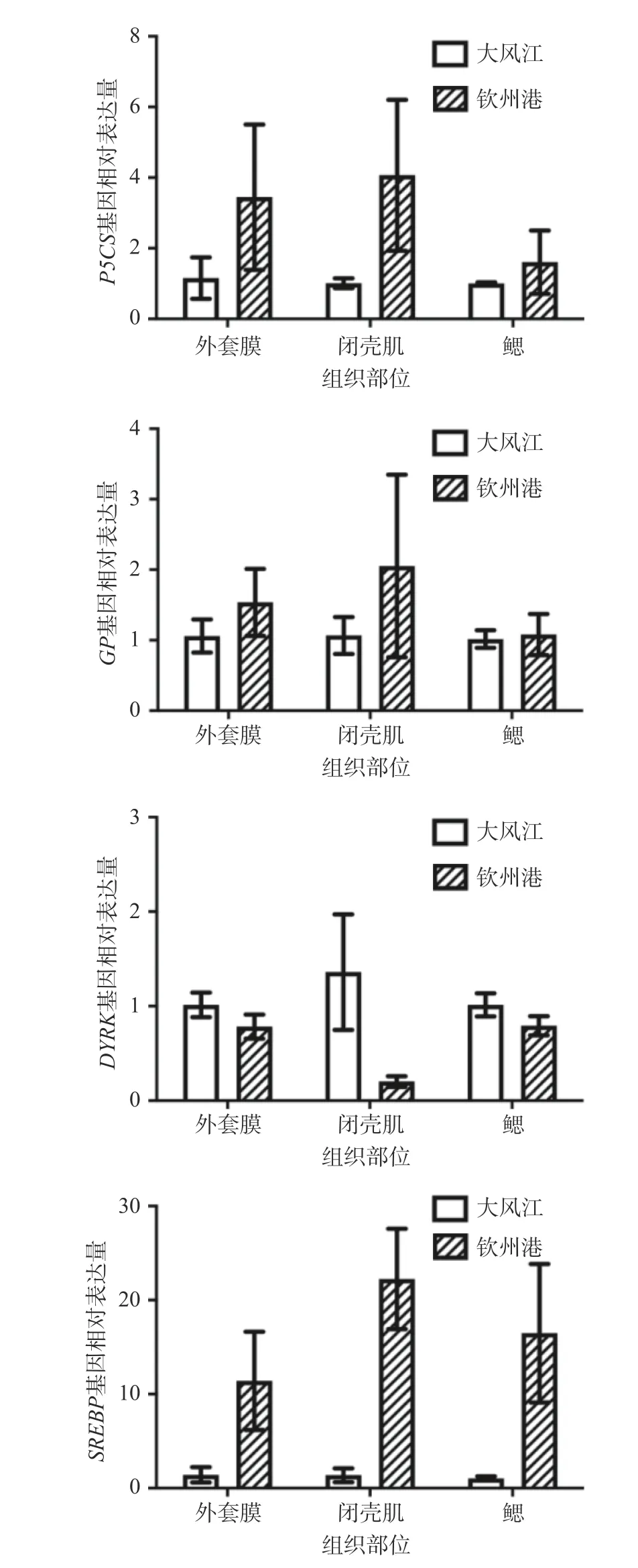
圖1 香港牡蠣營養(yǎng)代謝相關基因的組織表達分析Fig.1 Metabolism-related gene expression of C.hongkongensis in different tissues
P5CS基因是氨基酸代謝過程的關鍵酶,可以催化谷氨酸還原,該步驟是合成脯氨酸、鳥氨酸和精氨酸的關鍵步驟[28]。欽州港養(yǎng)殖區(qū)香港牡蠣的P5CS基因在外套膜、閉殼肌和鰓等組織的相對表達量分別是大風江組的3.0、4.0 和1.6 倍,表明欽州港養(yǎng)殖區(qū)香港牡蠣在氨基酸代謝及合成脯氨酸、鳥氨酸和精氨酸等方面更有優(yōu)勢。GP基因與葡萄糖代謝相關,琥珀酸脫氫酶在三羧酸循環(huán)中可以催化琥珀酸氧化成延胡索酸[29],而GP通過釋放葡萄糖-1-磷酸催化糖原分解,是這個過程的限速步驟[30]。欽州港組GP基因在外套膜、閉殼肌和鰓等組織的相對表達量分別是大風江組的1.4、1.9 和1.0 倍,表明欽州港養(yǎng)殖區(qū)香港牡蠣具有更強的葡萄糖代謝能力。
DYRK基因是一種劑量依賴性的絲氨酸/蘇氨酸激酶,在細胞增殖和分化的調控中起著關鍵作用,該基因家族成員可磷酸化許多底物,包括細胞周期的關鍵調節(jié)因子[31],可以通過減少體脂來抑制肥胖等代謝病[32]。此外,李春燕[11]通過Quantitative trait locus(QTL)分析認為DYRK基因是可影響牡蠣生長和脂肪酸性狀的重要標記基因。SREBP基因是內質網膜上的轉錄因子,可以激活下游的脂肪合成相關酶基因,促進脂質的合成與沉積[33?37]。大風江養(yǎng)殖區(qū)香港牡蠣的DYRK基因在外套膜、閉殼肌和鰓等組織的相對表達量分別是欽州港養(yǎng)殖區(qū)的1.3、6.7 和1.3 倍,而欽州港養(yǎng)殖區(qū)香港牡蠣的SREBP基因在外套膜、閉殼肌和鰓等組織的相對表達量分別是大風江養(yǎng)殖區(qū)的8.2、16.4 和16.1 倍,因此推測大風江養(yǎng)殖區(qū)香港牡蠣的脂肪分解代謝能力更強,而欽州港養(yǎng)殖區(qū)的香港牡蠣可能在脂肪合成與沉積上更有優(yōu)勢。
2.4 香港牡蠣代謝基因表達和營養(yǎng)成分相關性分析
由圖2 可知,DYRK的基因表達量和香港牡蠣常規(guī)營養(yǎng)成分含量的相關性明顯與P5CS、GP和SREBP這3 個基因的結果相反,表現為DYRK基因的表達量與能量、蛋白質、脂肪、氨基酸、膽固醇、維生素A 和維生素E 等含量均呈負相關關系;牡蠣的遺傳圖譜與營養(yǎng)品質性狀QTL 定位分析發(fā)現DYRK基因與花生四烯酸的代謝相關[11],花生四烯酸是一種ω-6 多不飽和脂肪酸,具有酯化膽固醇的作用,因此DYRK基因可能在脂肪酸代謝過程中具有一定的調控作用。P5CS、GP和SREBP這3 個基因的表達量則表現為與香港牡蠣的常規(guī)營養(yǎng)成分含量整體呈正相關關系,這3 個基因的功能分別與氨基酸代謝、葡萄糖代謝及脂質合成有關,相關性分析結果與其基因功能基本相符,表明P5CS、GP和SREBP這3 個基因與香港牡蠣的常規(guī)營養(yǎng)成分的相關性具有一致性。此外,GP基因的功能與糖原分解和葡萄糖代謝密切相關,而相關性分析結果發(fā)現GP基因的表達量與能量、脂肪、膽固醇和維生素E 的含量極顯著正相關(P<0.01),表明GP基因與香港牡蠣的能量、脂肪、膽固醇和維生素E 這4 個營養(yǎng)成分指標的相關性較強,進一步說明GP基因是研究香港牡蠣營養(yǎng)代謝的關鍵候選基因。
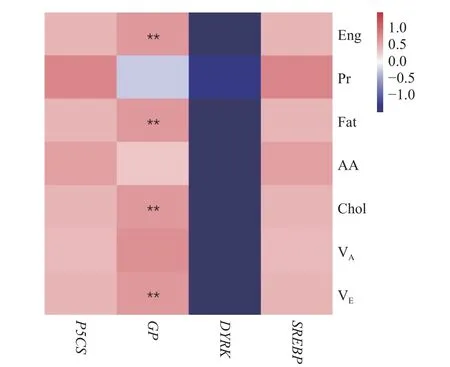
圖2 香港牡蠣基因表達與常規(guī)營養(yǎng)成分的相關性熱圖Fig.2 Heatmap of the correlation between gene expression and the main nutrient composition of C.hongkongensis
香港牡蠣代謝基因表達和17 種氨基酸含量的相關性分析結果如圖3 所示,P5CS、GP和SREBP這3 個基因的表達量與蘇氨酸、絲氨酸、谷氨酸、丙氨酸、蛋氨酸、異亮氨酸、亮氨酸、酪氨酸、苯丙氨酸、賴氨酸、組氨酸和精氨酸等多種氨基酸的含量整體呈正相關關系;P5CS基因是氨基酸代謝過程的關鍵酶,對脯氨酸、鳥氨酸和精氨酸的合成具有重要的作用,GP和SREBP的基因功能則分別與葡萄糖代謝和脂肪合成有關,而苯丙氨酸、異亮氨酸、蘇氨酸、酪氨酸、色氨酸等生糖氨基酸或生酮氨基酸又可通過不同的途徑合成糖類和脂類,因此P5CS、GP和SREBP這3 個基因可能參與了多種氨基酸的代謝調控過程[38]。由圖3 可以看出,DYRK基因的表達量則與脯氨酸、甘氨酸和胱氨酸呈正相關關系,與天冬氨酸、蘇氨酸、絲氨酸、谷氨酸、丙氨酸、纈氨酸、蛋氨酸、異亮氨酸、亮氨酸、酪氨酸、苯丙氨酸、賴氨酸、組氨酸和精氨酸等14 種氨基酸的含量呈明顯的負相關關系,但DYRK基因參與氨基酸代謝過程的調控機制還需要進一步深入研究。綜上所述,相關性分析結果表明P5CS、GP和SREBP等基因可能與香港牡蠣的營養(yǎng)成分及多種氨基酸合成和代謝具有一定的正向協同作用。

圖3 香港牡蠣基因表達與氨基酸含量的相關性熱圖Fig.3 Heatmap of the correlation between gene expression and the amino acid contents of C.hongkongensis
3 結論
本研究測定了廣西欽州灣兩個育肥養(yǎng)殖海區(qū)香港牡蠣的營養(yǎng)成分和代謝相關基因的表達情況,并首次分析了育肥期香港牡蠣營養(yǎng)成分和基因表達的相關性。結果顯示,大風江和欽州港兩個育肥養(yǎng)殖海區(qū)的香港牡蠣的組織營養(yǎng)成分存在一定的差異:與大風江養(yǎng)殖區(qū)相比,欽州港養(yǎng)殖區(qū)香港牡蠣的必需氨基酸的含量和比例以及必需氨基酸評分更高,表明其氨基酸平衡效果好,含量豐富且均勻,屬于優(yōu)質的蛋白質,具有較高的營養(yǎng)價值;而大風江養(yǎng)殖區(qū)香港牡蠣在鈣、鈉和鎂等礦物元素的含量上更有優(yōu)勢,且其風味氨基酸的比例高于欽州港養(yǎng)殖區(qū)牡蠣,表明大風江養(yǎng)殖區(qū)香港牡蠣鮮味程度更佳。此外,基因表達及相關性分析結果表明,欽州港養(yǎng)殖區(qū)的香港牡蠣可能在涉及氨基酸、葡萄糖等營養(yǎng)物質代謝和脂肪合成等方面的基因表達量更高,而大風江養(yǎng)殖區(qū)香港牡蠣的DYRK基因表達量更高,提示其在脂肪分解代謝及脯氨酸、甘氨酸和胱氨酸合成等方面的基因表達上更有優(yōu)勢。本研究為全面認識不同海區(qū)育肥期香港牡蠣的營養(yǎng)特征及營養(yǎng)評價候選基因的篩選提供了理論依據,對香港牡蠣深加工及資源合理開發(fā)利用具有一定的參考價值。
