過(guò)量銅在兩個(gè)柑橘品種幼苗中的分布特征及根細(xì)胞壁響應(yīng)機(jī)制
2022-07-18 08:59:36李欣鈺林妹蘭胡萍萍陳立松黃增榮
植物營(yíng)養(yǎng)與肥料學(xué)報(bào) 2022年6期
李欣鈺,林妹蘭,盧 飛,胡萍萍,陳立松,黃增榮
(福建農(nóng)林大學(xué)資源與環(huán)境學(xué)院,福建福州 350002)
銅是高等植物生長(zhǎng)發(fā)育必需的微量營(yíng)養(yǎng)元素之一,參與了光合電子傳遞,抗氧化保護(hù)以及細(xì)胞壁修飾等生化反應(yīng)[1]。在柑橘生產(chǎn)中,含銅菌劑常用于葉片噴施防治柑橘瘡痂病和黑斑病等[2]。連年噴施造成銅在柑橘園土壤中逐年累積[3–4]。土壤中的銅主要以正二價(jià)(Cu2+)形態(tài)存在。相比其他二價(jià)金屬元素,土壤有機(jī)質(zhì)對(duì)Cu2+的結(jié)合能力最強(qiáng)[5]。土壤有機(jī)質(zhì)對(duì)銅的強(qiáng)烈吸附減少了銅向柑橘樹(shù)體的遷移。但據(jù)本課題組對(duì)福建平和300多個(gè)琯溪蜜柚果園土壤和葉片的調(diào)查,仍有70.3%的葉片銅含量超過(guò)適宜濃度[6]。銅超標(biāo)蜜柚葉片占比高,一方面是由于在柑橘生產(chǎn)中過(guò)多施用含銅殺菌劑(如波爾多液、藍(lán)礬等)以及含銅微量元素肥料,導(dǎo)致噴落的銅逐年富集在果園土壤表層[7–8];另一方面是由于果農(nóng)在蜜柚園養(yǎng)分管理中偏施化肥,少施有機(jī)肥,造成果園土壤逐年酸化(300多個(gè)琯溪蜜柚果園土壤pH平均值為4.34,且90.0%的土壤樣品pH小于5.0[6]),促進(jìn)土壤中吸附性銅的解吸,提高土壤溶液中銅的活性。活化的銅通過(guò)根向柑橘體內(nèi)遷移,增大柑橘銅中毒風(fēng)險(xiǎn)[9]。因此,研究酸性條件下柑橘苗對(duì)過(guò)量銅的響應(yīng)機(jī)制對(duì)于優(yōu)化果園銅養(yǎng)分管理、防控柑橘園銅毒害、提高柑橘產(chǎn)量和品質(zhì)具有重要意義。
銅的遷移與再分布是植物應(yīng)答銅毒害最重要的機(jī)制之一[10]。在組織水平上,大量銅富集在多數(shù)植物的根中。銅在根中的固定直接減少了其向地上部的遷移。在細(xì)胞水平上,植物可通過(guò)“拒銅”或“區(qū)隔化”抵抗銅毒害。例如,植物可通過(guò)根系分泌的有機(jī)酸[11–12]或通過(guò)質(zhì)子泵的外排作用[13]將銅拒之根表。對(duì)于進(jìn)入細(xì)胞質(zhì)的銅,可通過(guò)銅轉(zhuǎn)運(yùn)蛋白將其轉(zhuǎn)運(yùn)到液泡中,以降低對(duì)同化器官的破壞[14]。在亞細(xì)胞水平上,有關(guān)銅的主要分布部位目前仍存在爭(zhēng)議。如Mwamba 等[15]研究表明,過(guò)量銅主要結(jié)合在油菜根的可溶性成分。Wan等[16]研究表明,過(guò)量銅主要富集在蘋(píng)果根細(xì)胞壁上。由于植物細(xì)胞可溶性成分復(fù)雜,近年來(lái)有關(guān)植物對(duì)銅區(qū)隔與固定的研究主要關(guān)注植物根細(xì)胞壁的多糖組分。植物根細(xì)胞壁主要由纖維素、半纖維素和果膠等多糖組分構(gòu)成[17–18]。各多糖組分富含帶有負(fù)電荷的羥基和羧基等,可通過(guò)配位吸附固定銅。目前,有關(guān)過(guò)量重金屬在細(xì)胞壁上的主要結(jié)合位點(diǎn)有不同的報(bào)道。如Ren等[19]研究表明,蓖麻根細(xì)胞壁的半纖維素組分是銅的主要結(jié)合位點(diǎn)。Yang等[20]研究表明,甘蔗根細(xì)胞壁果膠組分是錳的主要累積部位。
根細(xì)胞壁對(duì)銅的固定與植物耐銅性相關(guān)。研究表明,過(guò)量銅主要固定于蘋(píng)果須根上,在須根亞細(xì)胞水平銅分布特征表現(xiàn)為細(xì)胞壁>質(zhì)體>可溶性部分>線粒體>細(xì)胞核,且耐銅品種須根細(xì)胞壁銅累積量顯著高于銅敏感品種[21]。進(jìn)一步研究表明,根細(xì)胞壁對(duì)銅的固定與其微結(jié)構(gòu)變化有關(guān)[22–23]。例如,經(jīng)200 μmol/L銅處理30天的毛竹根銅濃度可達(dá)810 mg/kg,而轉(zhuǎn)移至地上部的銅濃度僅為91.0 mg/kg,且過(guò)量銅處理的細(xì)胞壁可見(jiàn)明顯扭曲變形與增厚[24]。根細(xì)胞壁增厚現(xiàn)象在受銅毒害影響的牛至[25]和蓖麻幼苗中皆有報(bào)道[26–27]。FTIR分析顯示,大部分銅結(jié)合到細(xì)胞壁纖維素、半纖維素、果膠和蛋白質(zhì)的羧基、羥基和氨基甲基等官能團(tuán)上[28–29]。Krzes?owska[23]的研究結(jié)果顯示,含羧基的多糖數(shù)量決定了細(xì)胞壁結(jié)合金屬離子的能力。張曉斌等[30]的研究表明,耐重金屬植物根細(xì)胞壁相應(yīng)基團(tuán)表現(xiàn)出不同程度的位移或吸光度的差異。目前,關(guān)于過(guò)量銅在柑橘樹(shù)體中的遷移與微分布以及過(guò)量銅誘導(dǎo)的根細(xì)胞壁微結(jié)構(gòu)變化的報(bào)道較少。
本研究以具有耐銅差異的雪柑和沙田柚實(shí)生苗[31]為試驗(yàn)材料,設(shè)置連續(xù)18周含銅濃度分別為0.5 μmol/L (對(duì)照)和 300 μmol/L (銅過(guò)量)的銅處理,研究過(guò)量銅在雪柑和沙田柚實(shí)生苗中的分布特征以及柑橘根細(xì)胞壁微觀結(jié)構(gòu)變化,旨在進(jìn)一步解析柑橘過(guò)量銅應(yīng)答機(jī)制,為優(yōu)化柑橘銅養(yǎng)分管理,實(shí)現(xiàn)高產(chǎn)優(yōu)質(zhì)栽培提供理論和實(shí)踐依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料與處理
挑選飽滿(mǎn)的沙田柚和雪柑種子,播于含有干凈河沙的黑色穴盤(pán)中。出芽5周后,將長(zhǎng)勢(shì)一致的兩個(gè)柑橘品種實(shí)生苗移栽到含有干凈河沙的陶瓷花盆中,沙田柚和雪柑各24盆,每盆2株,在福建農(nóng)林大學(xué)金山校區(qū)的溫室大棚內(nèi)自然光下生長(zhǎng),每?jī)商鞚惨淮螤I(yíng)養(yǎng)液,每盆500 mL。營(yíng)養(yǎng)液配方:KNO32.5 mmol/L、Ca(NO3)22.5 mmol/L、KH2PO40.5 mmol/L、MgSO41 mmol/L、H3BO310 μmol/L、MnCl22 μmol/L、ZnSO42 μmol/L、CuCl20.5 μmol/L、(NH4)6Mo7O240.065 μmol/L 和 Fe-EDTA 20 μmol/L。
移栽6周后,對(duì)柑橘實(shí)生苗進(jìn)行不同濃度銅處理,銅濃度分別為 0.5 μmol/L (對(duì)照,Cu0.5)和 300 μmol/L (銅過(guò)量,Cu300),營(yíng)養(yǎng)液 pH 用 1 mol/L HCl或NaOH調(diào)至4.30~4.50。處理18周后,柑橘苗按主根、側(cè)根、莖和葉分類(lèi)采集。其中,柑橘根樣品用自來(lái)水洗凈沙粒后用預(yù)冷的0.5 mol/L EDTANa2浸泡10 min再用蒸餾水清洗3次,經(jīng)吸水紙吸干后采集。取部分柑橘苗側(cè)根樣品立刻放入固定液中用于透射電鏡觀察;另取部分柑橘苗側(cè)根鮮樣用液氮固定,保存于–80℃冰箱,用于亞細(xì)胞組分分離和銅含量測(cè)定。剩余側(cè)根和主根鮮樣放入70℃烘箱烘干后,用于粗細(xì)胞壁的提取及銅含量的測(cè)定。柑橘地上部樣品用蒸餾水清洗3次,表面干燥后放入70℃烘箱烘干至恒重用于銅含量的測(cè)定。
1.2 試驗(yàn)方法
1.2.1 柑橘幼苗主根、側(cè)根、莖和葉中銅含量的測(cè)定
烘干至恒重的柑橘幼苗主根、側(cè)根、莖和葉粉末,經(jīng)濃硝酸–濃高氯酸(V∶V=5∶1)過(guò)夜消化后,用原子吸收光譜法 (PerkinElmer Singapore Pte Let.,Singapore)測(cè)定雪柑和沙田柚主根、側(cè)根、莖和葉中的銅含量[32],每個(gè)樣品4個(gè)重復(fù)。
1.2.2 亞細(xì)胞組分的分離與銅含量的測(cè)定 參考周麗珍等[33]的方法分離亞細(xì)胞組分。稱(chēng)取適量柑橘根尖新鮮樣品于預(yù)冷的勻漿液(50 mmol/L順丁烯二酸鹽緩沖液,pH 7.8;250 mmol/L 蔗糖;1 mmol/L 二硫赤蘚糖醇)中,研磨成勻漿,轉(zhuǎn)入50 mL離心管;于 4℃ 高速冷凍離心機(jī) (Thermo Scientific Sorvall Legend Micro 17R,USA)中,以 3000 r/min 離心1 min,底部沉淀為細(xì)胞壁組分;上清液轉(zhuǎn)入新的離心管后,以 15000 r/min 離心 45 min,底層為細(xì)胞器組分;上清液轉(zhuǎn)入新的離心管,為可溶性部分(含細(xì)胞質(zhì)及液泡),所得所有組分均于4℃保存。分離得到的亞細(xì)胞組分細(xì)胞壁、細(xì)胞器及可溶性部分,經(jīng)濃硝酸–濃高氯酸(V∶V=5∶1)消化后,利用NexION 300XICP-MS (PerkinElmer,Shelton,CT,USA)測(cè)定3個(gè)組分的銅含量,每個(gè)樣品含3個(gè)重復(fù)。采用柑橘葉國(guó)家標(biāo)準(zhǔn)物質(zhì)GBW10020 (GSB–11)進(jìn)行分析質(zhì)量控制。其中,柑橘葉標(biāo)準(zhǔn)品銅含量為6.6±0.5(μg/g, DW),質(zhì)控結(jié)果為 6.83±0.48 (μg/g, DW)。
1.2.3 細(xì)胞壁的提取、多糖組分的分離與銅含量測(cè)定
參考Zhong等[34]的方法,提取細(xì)胞壁并分離細(xì)胞壁多糖組分。稱(chēng)取適量烘干并磨碎成粉末的柑橘側(cè)根樣品于離心管中,采用75%乙醇、丙酮、甲醇–氯仿(V∶V=1∶1)和甲醇4種提取劑(固液比為0.1 g∶10 mL)依次分別提取。加入提取劑充分混勻后,靜置 20 min,在 12000 r/min 的離心機(jī)中離心 10 min,棄去上清液,并加入去離子水清洗,混勻并離心,棄去上清液,得到粗細(xì)胞壁。含粗細(xì)胞壁的離心管覆蓋單層保鮮膜并扎孔,于55 ℃烘箱烘干至恒重,稱(chēng)量粗細(xì)胞壁的質(zhì)量,計(jì)算細(xì)胞壁提取率。稱(chēng)取0.01 g干燥的細(xì)胞壁樣品于離心管中,加入2 mL 0.5%草酸銨 (含 0.1% NaBH4,pH=4.00),沸水浴 1 h 后12000 r/min離心 10 min,取上清液于新的離心管中,重復(fù)3次合并上清液得到果膠組分。沉淀加入4% KOH (含 0.1% NaBH4),并于室溫下在 200 r/min的搖床振蕩 8 h 后 12000 r/min 離心 10 min,取上清液于新的離心管中,重復(fù)3次提取(共24 h,每次需8 h),合并上清液,上清液即為半纖維素Ⅰ類(lèi)。沉淀加 24% KOH (含 0.1% NaBH4)后重復(fù)上步提取操作得到半纖維素Ⅱ類(lèi)組分。最終沉淀為纖維素,烘干至恒重并稱(chēng)重。得到的不同細(xì)胞壁多糖組分保存于4℃冰箱備用。分離得到的3種細(xì)胞壁多糖組分和纖維素,經(jīng)濃硝酸–濃高氯酸(V∶V = 5∶1)消化后,用ICP-MS測(cè)定各組分中的銅含量,每個(gè)樣品5個(gè)重復(fù)。
1.2.4 側(cè)根透射電鏡的觀察 參考Huang等[35]的方法,處理柑橘側(cè)根根尖鮮樣。柑橘側(cè)根在裝有3%戊二醛–1.5%多聚甲醛固定液的注射器中抽真空,并于3%戊二醛–1.5%多聚甲醛的固定液中4℃預(yù)固定3 h,用 0.1 mol/L 磷酸緩沖液 (pH 7.2)清洗 3 次,每次間隔15 min,在1%鋨酸–1.5%亞鐵氰化鉀的固定液中固定2 h,用蒸餾水清洗3次,每次15 min,最后用30%、50%、70%、80%、90%、95%、100%的乙醇進(jìn)行梯度脫水,每級(jí)15 min,其中100%酒精洗脫3次,加丙酮清洗2次,每次間隔10 min,加樹(shù)脂–丙酮(V∶V=1∶1)的混合液置換1次,在搖床上搖2 h,加樹(shù)脂–丙酮(V∶V=3∶1)的混合液滲透1次,在搖床上搖2 h,取出,在濾紙上吸去殘余丙酮,加全樹(shù)脂過(guò)夜。取出樣品,將單個(gè)樣品分開(kāi)放入含全樹(shù)脂的PCR小管中,用環(huán)氧樹(shù)脂618包埋,并做好標(biāo)記,凝固后用超薄切片機(jī)切片,在日立HT7700透射電鏡下觀察并拍照,每個(gè)樣品4個(gè)重復(fù)。
1.2.5 FTIR分析 參考Liu等[36]的方法,稱(chēng)取約2 mg干燥細(xì)胞壁與200 mg KBr放入瑪瑙研缽中研磨混合均勻后倒入壓模中,在真空條件下得樣品薄片,放入 FTIR 采樣器 (Thermo Scientific Nicolet iS50,USA)測(cè)定紅外光譜,在 4000~400 cm–1光譜范圍內(nèi),分辨率為4 cm–1,掃描累加次數(shù)32次,以各吸收峰的位置和吸收率為指標(biāo),構(gòu)建數(shù)據(jù)庫(kù),即測(cè)得各組樣品的原始紅外光譜數(shù)據(jù)。在OMNIC 8.0軟件上進(jìn)行自動(dòng)基線校正和平滑。為了提高譜圖的分辨率,以便減少譜圖疊加,對(duì)不同處理選擇含有多糖、酰胺和酯等特征信息的1800~800 cm?1光譜范圍進(jìn)行精細(xì)表征,每個(gè)樣品3個(gè)重復(fù)。
1.3 統(tǒng)計(jì)分析
采用完全隨機(jī)進(jìn)行試驗(yàn)設(shè)計(jì),4個(gè)不同處理組合間采用 2 (柑橘品種)×2 (銅處理水平)雙因素方差分析進(jìn)行差異性檢測(cè),利用SPSS 16.0軟件(Chicago,USA)進(jìn)行統(tǒng)計(jì)分析,統(tǒng)計(jì)結(jié)果表示為平均數(shù)±標(biāo)準(zhǔn)誤差。利用 Image-Pro Plus 6.0 (Media Cybernetics,USA)軟件處理透射電鏡照片,并使用Origin 2018軟件(OriginLab,USA)制圖。
2 結(jié)果與分析
2.1 過(guò)量銅在兩個(gè)柑橘品種幼苗主根、側(cè)根、莖和葉中的分布
如表1所示,在柑橘幼苗的組織水平上,不論是對(duì)照 (Cu 0.5 μmol/L)還是過(guò)量銅處理 (Cu 300 μmol/L),側(cè)根中的銅含量均最高。以含量最低的柑橘主根銅含量(雪柑和沙田柚分別為4.46和2.34 μg/g, DW)作為標(biāo)準(zhǔn)(數(shù)值為1),對(duì)照組雪柑和沙田柚主根、側(cè)根、莖和葉的銅含量相對(duì)值分別為 1、6.16、2.02、1.49 和 1、8.06、3.37、3.23。相比對(duì)照,300 μmol/L銅處理顯著提高了兩個(gè)柑橘品種幼苗各部位的銅含量。以含量最低的柑橘葉銅含量(雪柑和沙田柚分別為29.21和34.86 μg/g, DW)作為標(biāo)準(zhǔn)(數(shù)值為1),300 μmol/L銅處理的雪柑和沙田柚主根、側(cè)根、莖和葉的銅濃度相對(duì)值分別為16.3、190、1.18、1 和 14.7、194、1.26、1。對(duì)照組的雪柑和沙田柚側(cè)根銅分布率分別為57.32%和51.31%。300 μmol/L 銅處理的雪柑和沙田柚側(cè)根銅分布率分別提高到90.87%和91.99%。但過(guò)量銅處理降低了銅在柑橘莖和葉的分布率。品種間比較結(jié)果顯示,銅處理下沙田柚側(cè)根銅含量顯著高于雪柑。
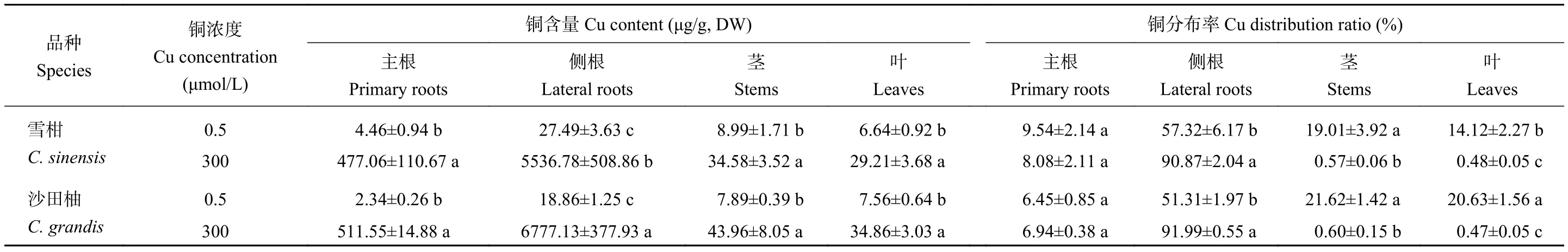
表1 不同銅處理的兩個(gè)柑橘品種幼苗主根、側(cè)根、莖和葉中的銅含量與分布率Table 1 Cu content and distribution ratio in the primary roots, lateral roots, stems and leaves of two citrus species under different Cu concentrations
2.2 過(guò)量銅在兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁、細(xì)胞器和可溶性部分的分布
如表2所示,對(duì)照組中的柑橘幼苗側(cè)根細(xì)胞器銅濃度最高,細(xì)胞壁中銅濃度最低。對(duì)照組的雪柑和沙田柚側(cè)根細(xì)胞器銅分布率分別為58.61%和39.26%;300 μmol/L銅處理顯著提高了兩個(gè)柑橘品種幼苗側(cè)根各亞細(xì)胞組分中的銅含量。過(guò)量銅處理的柑橘幼苗側(cè)根亞細(xì)胞水平上可溶性部分銅濃度最高,細(xì)胞器濃度最低,過(guò)量銅處理的雪柑和沙田柚側(cè)根細(xì)胞器的銅分布率分別為14.34%和11.49%。相比對(duì)照,過(guò)量銅處理提高了細(xì)胞壁和可溶性部分的銅分布率,但顯著降低了細(xì)胞器的銅分布率。過(guò)量銅處理的沙田柚細(xì)胞壁中的銅含量與分布率均顯著高于雪柑。
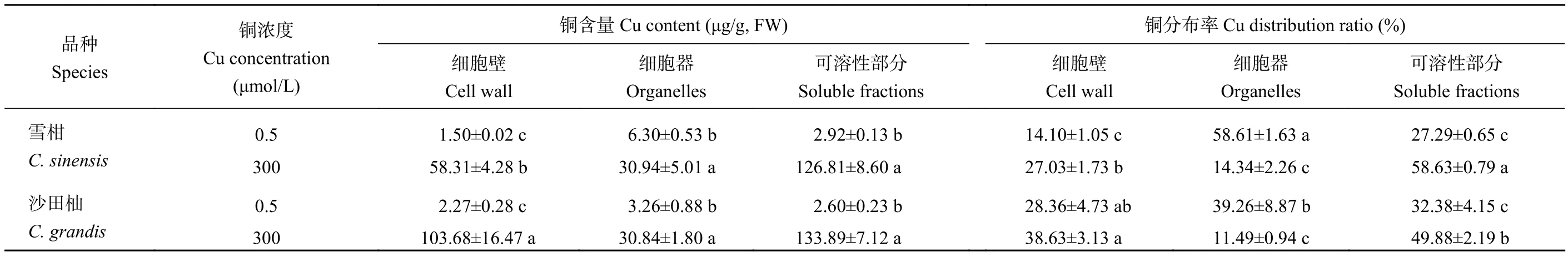
表2 不同銅處理的兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁、細(xì)胞器和可溶性部分的銅含量與銅分布率Table 2 Cu content and distribution ratio in the cell wall, organelles and soluble fractions of lateral roots of two citrus species under different Cu concentrations
2.3 過(guò)量銅在兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁果膠、半纖維素Ⅰ、半纖維素Ⅱ和纖維素中的分布
由表3可知,對(duì)照和過(guò)量銅處理,銅在細(xì)胞壁多糖組分中的分布均為果膠>纖維素>半纖維素Ⅰ>半纖維素Ⅱ。與對(duì)照相比,過(guò)量銅處理柑橘側(cè)根細(xì)胞壁多糖組分中的銅含量均顯著提高,果膠銅含量上升尤為顯著。對(duì)照組的雪柑和沙田柚側(cè)根細(xì)胞壁果膠銅分布率分別為44.69%和37.15%;過(guò)量銅處理后,雪柑和沙田柚側(cè)根銅分布率分別提高到98.45%和98.48%。過(guò)量銅處理的雪柑和沙田柚側(cè)根細(xì)胞壁果膠銅含量較對(duì)照分別提高了170和290倍。品種間比較發(fā)現(xiàn),過(guò)量銅處理后的沙田柚側(cè)根細(xì)胞壁果膠銅含量顯著高于雪柑。從銅在柑橘側(cè)根細(xì)胞壁多糖組分中的分布率來(lái)看,過(guò)量銅處理的半纖維素Ⅰ、半纖維素Ⅱ和纖維素中的銅分布率顯著降低,而果膠銅分布率顯著提高。過(guò)量銅處理的兩個(gè)柑橘品種側(cè)根細(xì)胞壁果膠銅分布率均高于98%。
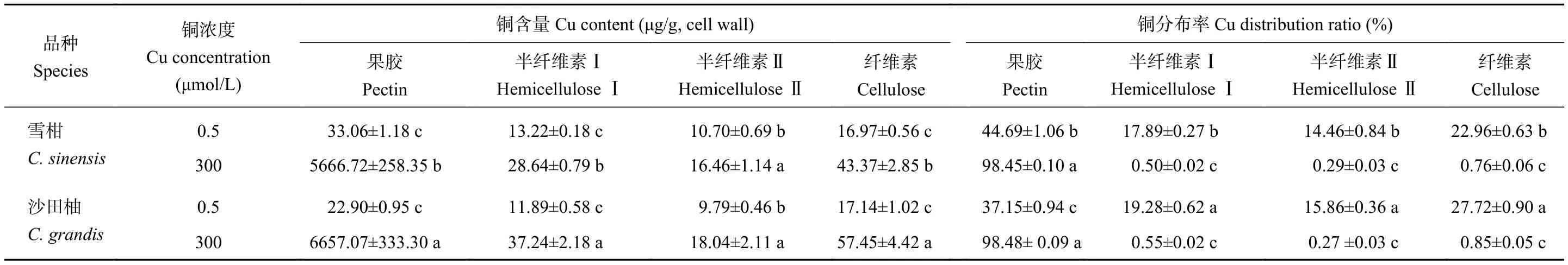
表3 不同銅處理的兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁多糖組分中銅含量與銅分布率Table 3 Cu content and distribution ratio in polysaccharide compositions of lateral roots cell wall of two citrus species under different Cu concentrations
2.4 過(guò)量銅對(duì)兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁的影響
相比對(duì)照,過(guò)量銅處理顯著提高了兩個(gè)柑橘品種幼苗的細(xì)胞壁提取率,且過(guò)量銅處理的沙田柚細(xì)胞壁提取率顯著高于雪柑(圖1)。透射電鏡觀察表明(圖2),過(guò)量銅處理的雪柑側(cè)根線粒體模糊,但沙田柚側(cè)根線粒體膜結(jié)構(gòu)清晰可見(jiàn)。相比對(duì)照,過(guò)量銅處理的兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁增厚。不論是對(duì)照還是過(guò)量銅處理,沙田柚側(cè)根細(xì)胞壁厚度均顯著高于雪柑(圖3)。
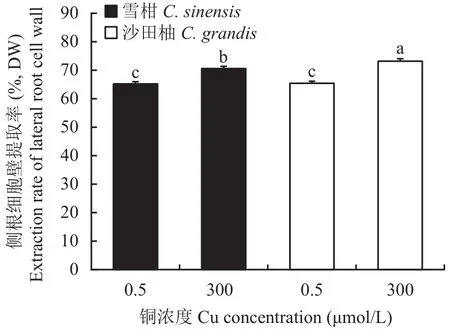
圖1 不同銅濃度處理下兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁提取率Fig.1 The extraction rate of lateral roots cell wall of two citrus species under different Cu concentrations
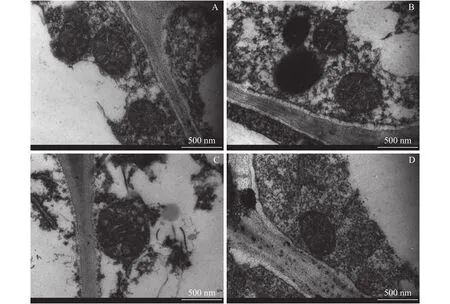
圖2 不同銅濃度處理下兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁微觀形態(tài)Fig.2 The micromorphology of lateral roots cell wall of two citrus species under different Cu concentrations
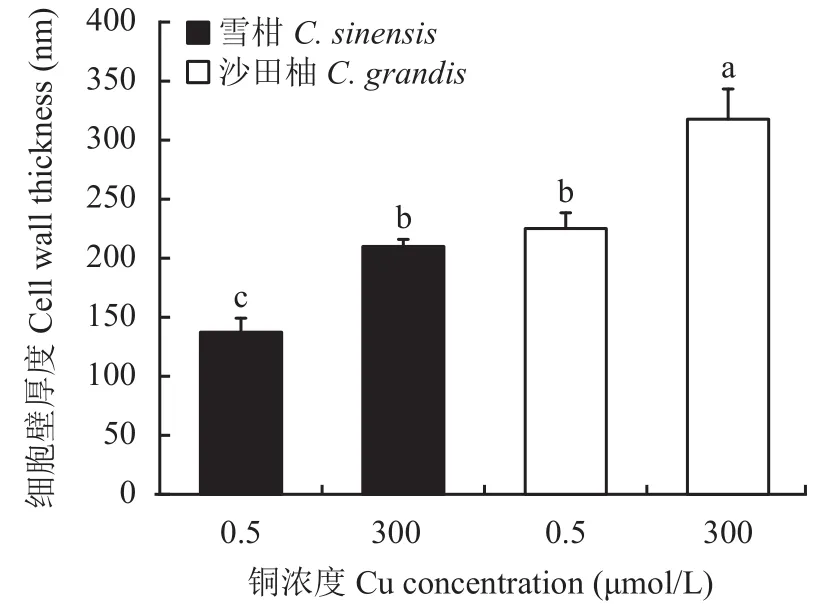
圖3 不同銅濃度處理下兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁厚度Fig.3 The lateral root cell wall thickness of two citrus species under different Cu concentrations
2.5 銅處理的兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁FTIR分析
對(duì)照和過(guò)量銅處理后的雪柑和沙田柚幼苗側(cè)根細(xì)胞壁的 FTIR 譜圖如圖4 所示。3338 cm–1(No.1)附近的吸收峰為—OH和—NH伸縮振動(dòng)峰的疊加,主要來(lái)自于根細(xì)胞壁中脂肪酸、蛋白質(zhì)和碳水化合物(果膠、半纖維素、纖維素、多糖),2921 cm–1(No.2)附近的吸收峰為甲基C―H鍵的對(duì)稱(chēng)伸縮振動(dòng),主要來(lái)自于細(xì)胞壁中親水脂分子,1736 cm–1(No.3)附近的吸收峰歸屬為酯化果膠中酯基C=O的伸縮振動(dòng),1652 cm–1(No.4)處的吸收峰為酰胺Ⅰ帶C—N伸縮,是細(xì)胞壁蛋白質(zhì)上的特征峰,1508 cm–1(No.5)附近的吸收峰為蛋白質(zhì)上的酰胺Ⅱ帶N―N彎曲和伸縮特征吸收峰,1423 cm–1(No.6)附近的吸收峰歸屬為果膠質(zhì)糖醛酸中羧酸根離子COO–的特征吸收峰,1374 cm–1(No.7)附近的吸收峰為纖維素中甲基的伸縮振動(dòng)吸收峰,1337 cm–1(No.8)附近的吸收峰對(duì)應(yīng)于纖維素中醇羥基的C―O弱吸收峰,1247 cm–1(No.9)處對(duì)應(yīng)硫酸鹽C―O―S或羧基C―O或磷酸鹽 C―O―P 的特征吸收峰,1150 cm–1(No.10)左右為果膠的多糖環(huán)結(jié)構(gòu)的C―C或C―O吸收峰,1035 cm–1(No.11)附近的吸收峰主要為纖維素的多糖鏈C―C的特征吸收峰。
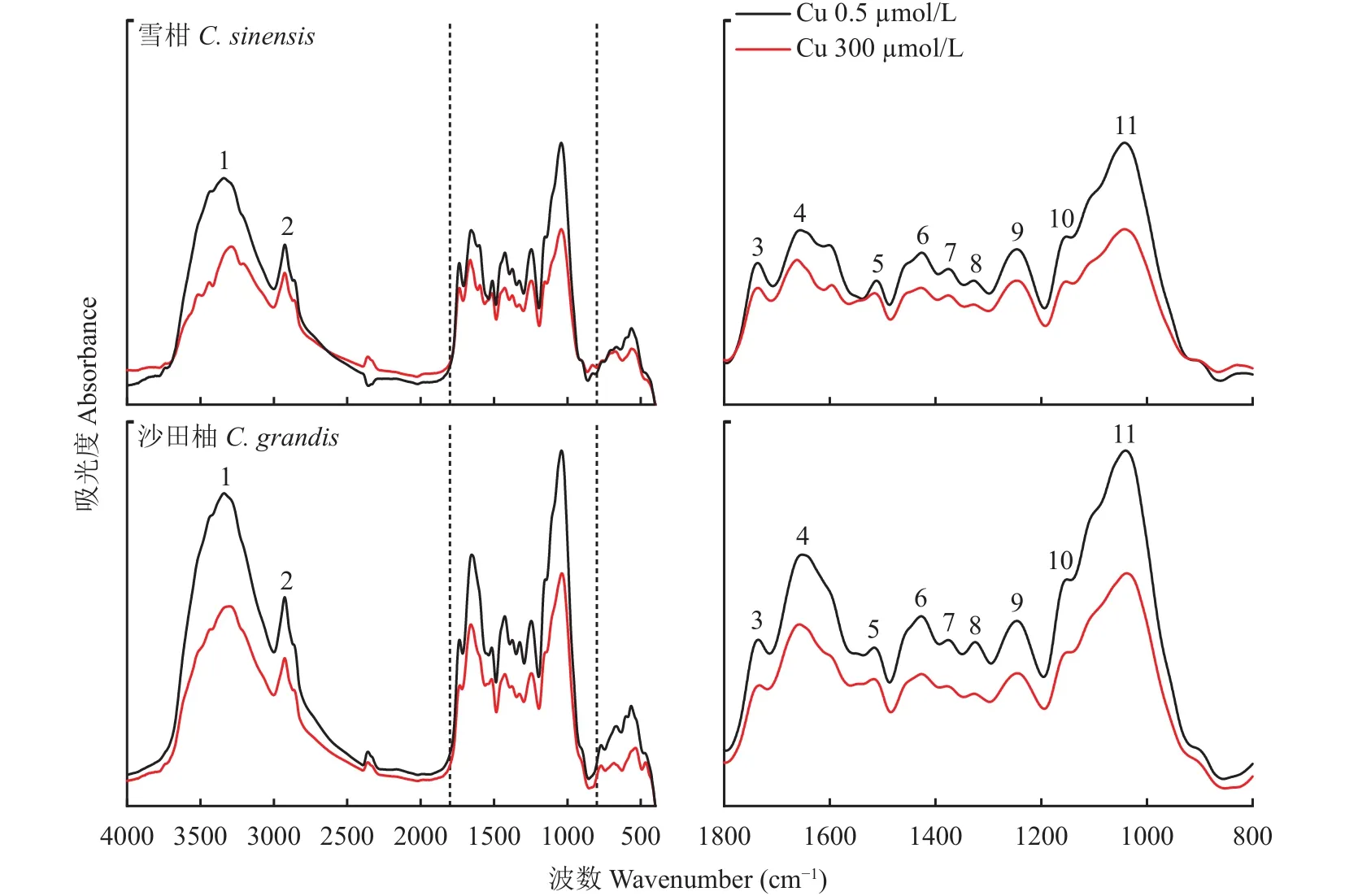
圖4 不同銅濃度處理下的兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁紅外光譜Fig.4 The FTIR spectra of Cu-treated lateral roots cell wall of two citrus species
參考張虹等[37]的方法,以 2921 cm–1處―CH3中C―H的特征吸收峰的吸光度A2921為標(biāo)準(zhǔn)值,計(jì)算其它特征峰的吸光值與A2921的比值,通過(guò)該比值的大小變化半定量分析過(guò)量銅處理的雪柑和沙田柚側(cè)根細(xì)胞壁官能團(tuán)特征峰變化(表4)。雪柑和沙田柚幼苗經(jīng)銅處理后,根細(xì)胞壁上的官能團(tuán)在No.1特征峰(—OH/—NH)、No.6特征峰(羧酸鹽C=O不對(duì)稱(chēng)伸縮振動(dòng))和No.10特征峰(果膠多糖環(huán)結(jié)構(gòu)C—C或C—O)發(fā)生了較明顯的偏移,且雪柑主要向高頻偏移,沙田柚主要向低頻偏移。另外,雪柑在No.4特征峰(酰胺Ⅰ帶C—N)處向高頻偏移了3 cm–1,沙田柚在No.3特征峰(酯基C=O)、No.5特征峰(酰胺Ⅱ帶N—N)及No.9特征峰(硫酸鹽C—O—S或羧基C—O或磷酸鹽C—O—P)處向低頻偏移,表明—OH、果膠、蛋白質(zhì)、羧酸鹽、硫酸鹽、磷酸鹽及纖維素在一定程度上參與了銅吸附。與對(duì)照相比,過(guò)量銅處理后沙田柚A1734/A2921高于雪柑A1737/A2921,表明過(guò)量銅處理的沙田柚根細(xì)胞壁中果膠質(zhì)甲基酯化程度上升明顯;過(guò)量銅處理后,沙田柚 A1653/A2921和 A1509/A2921上升值分別高于雪柑A1655/A2921和A1509/A2921,表明過(guò)量銅處理的沙田柚根細(xì)胞壁蛋白質(zhì)含量升高顯著;與對(duì)照相比,過(guò)量銅處理后,沙田柚A1373/A2921和A1244/A2921比值升高,而過(guò)量銅處理后雪柑的A1373/A2921和A1247/A2921比值較對(duì)照組降低,可知過(guò)量銅使沙田柚根細(xì)胞壁中的纖維素、硫酸鹽、羧酸鹽及磷酸鹽含量增加。過(guò)量銅處理后,相比沙田柚,雪柑根細(xì)胞壁A3293/A2921和A1426/A2921的比值有明顯降低,意味著其根細(xì)胞壁中―OH和果膠在銅處理下顯著減少;過(guò)量銅處理后,相比沙田柚,雪柑的A1337/A2921、A1156/A2921和A1035/A2921的比值下降較大,可能與過(guò)量銅處理后的雪柑根細(xì)胞壁中主要組分纖維素含量減少有關(guān)。
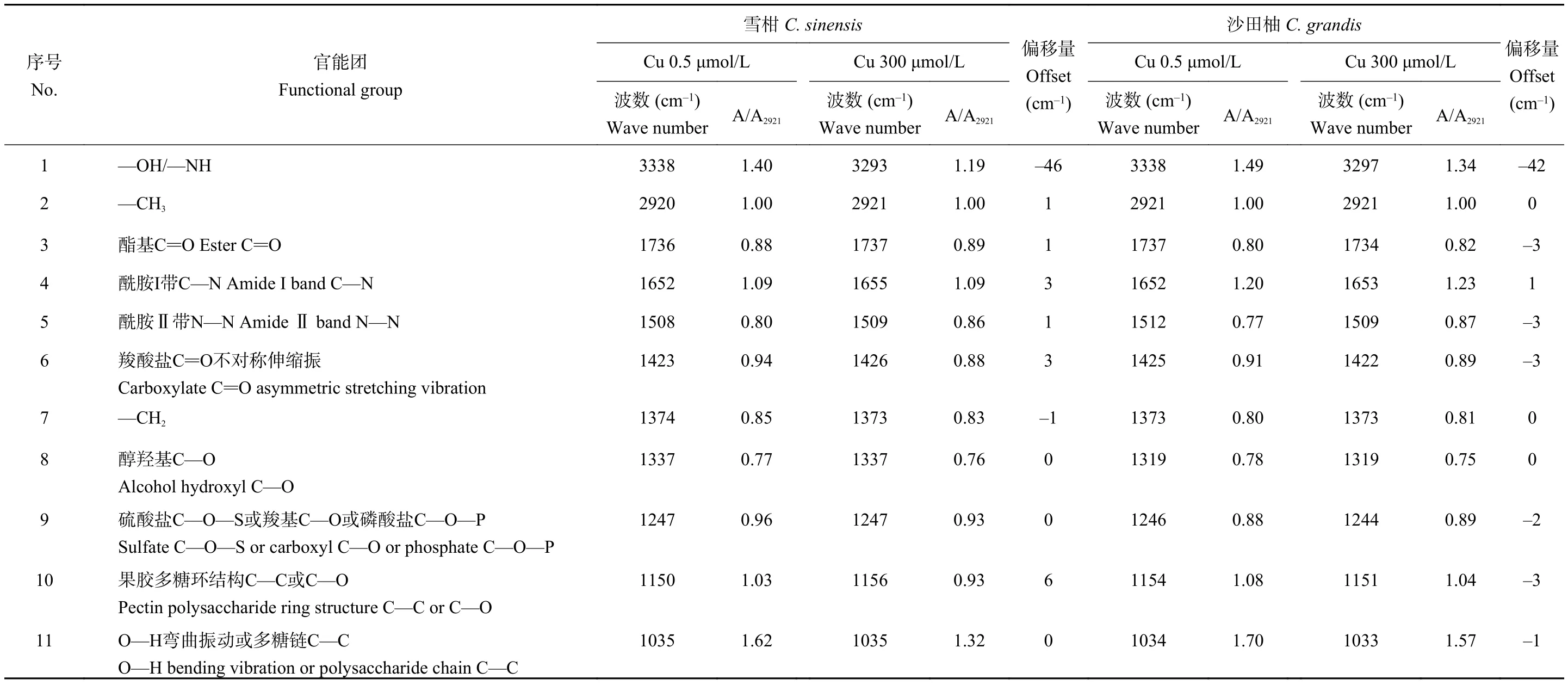
表4 不同銅濃度處理下兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁紅外光譜特征峰的半定量分析Table 4 The semi-quantitative analysis of FTIR spectra of Cu-treated lateral roots cell wall of two citrus species
3 討論
根是植物最早感知根際重金屬濃度,啟動(dòng)脅迫響應(yīng)最關(guān)鍵的部位[38]。銅在根中的固定,減少了其向地上部的裝載,起到保護(hù)地上部同化器官的作用。如,Hippler等[39]研究表明,24 μmol/L 銅處理 90 天后的水培甜橙幼苗根中累積的銅占植株吸收銅總量的80.0%。本研究結(jié)果進(jìn)一步表明,根際銅過(guò)量時(shí),柑橘苗側(cè)根是銅主要的累積部位。這與史萍等[40]在銅處理的牡丹根中的研究結(jié)果一致。相比銅處理的主根,雪柑和沙田柚幼苗側(cè)根的銅濃度分別提高了10.6和13.2倍。銅處理下,柑橘苗組織水平上的銅分布特征表現(xiàn)為側(cè)根>主根>莖>葉。相比對(duì)照,過(guò)量銅處理顯著提高了柑橘幼苗不同組織的銅濃度。300 μmol/L砂培銅處理的沙田柚側(cè)根銅濃度顯著高于雪柑可能與沙田柚側(cè)根對(duì)銅的固定能力較強(qiáng)有關(guān)。
亞細(xì)胞組分中重金屬的分布特征與植物耐性機(jī)制有關(guān)[41]。本研究顯示,在亞細(xì)胞水平上,銅主要分布在對(duì)照組柑橘幼苗側(cè)根的細(xì)胞器組分。過(guò)量銅處理下,細(xì)胞器中的銅相對(duì)含量顯著降低,但細(xì)胞壁和可溶性部分銅含量占比顯著提高。這與柑橘銅響應(yīng)機(jī)制有關(guān)。當(dāng)柑橘幼苗根際銅濃度較低時(shí),銅主要存在于細(xì)胞器,參與細(xì)胞代謝反應(yīng)。當(dāng)銅濃度過(guò)高時(shí),過(guò)量銅主要分布于細(xì)胞可溶性部分與細(xì)胞壁,而細(xì)胞器銅相對(duì)濃度顯著降低可以減少過(guò)量銅對(duì)細(xì)胞器的結(jié)構(gòu)損傷和功能破壞。通常認(rèn)為,根細(xì)胞壁是植物響應(yīng)重金屬脅迫的“第一道屏障”[42]。例如,劉婷婷[43]揭示了水培條件下,銅濃度為100 μmol/L時(shí),耐銅植物海州香薷根細(xì)胞壁銅累積量在亞細(xì)胞水平占比高達(dá)68.6%。類(lèi)似的報(bào)道顯示,過(guò)量鋅處理的欒樹(shù)、臭椿和銀杏根細(xì)胞壁中鋅的累積量最高可占亞細(xì)胞鋅含量的82.6%[44]。胡蘿卜側(cè)根細(xì)胞壁是過(guò)量鉛富集的主要部位[45]。玉米根細(xì)胞壁也是銅[46]、鎘[47]和釩[48]等重金屬的主要分布區(qū)域。不同的是,過(guò)量鎘在油菜根中的分布特征表現(xiàn)為細(xì)胞液>細(xì)胞器>細(xì)胞壁[49]。本研究結(jié)果顯示,盡管300 μmol/L砂培銅處理都顯著提高了兩個(gè)柑橘品種幼苗根細(xì)胞壁和可溶性部分的銅含量,但相比細(xì)胞壁,可溶性部分結(jié)合的銅含量占比更高。這與經(jīng)鉛、鎘和鋅處理的馬纓丹[41]以及經(jīng)砷處理的狹葉香蒲[50]上的研究結(jié)果一致。亞細(xì)胞可溶性部分富含小分子有機(jī)酸以及螯合態(tài)蛋白等,可以有效結(jié)合并固定銅,通過(guò)液泡對(duì)過(guò)量銅的區(qū)隔化作用降低細(xì)胞銅損傷[51]。鑒于細(xì)胞可溶性成分種類(lèi)多,成分雜,有待通過(guò)質(zhì)譜分析與蛋白分離等技術(shù)進(jìn)一步研究。
根細(xì)胞壁多糖組分由葡萄糖、阿拉伯糖以及半乳糖醛酸等單糖聚合而成[52]。半乳糖醛酸富含半乳糖殘基,可作為細(xì)胞壁的陽(yáng)離子吸附位點(diǎn)[53]。報(bào)道顯示,細(xì)胞壁各多糖組分的銅結(jié)合能力存在差異。例如,任超[54]報(bào)道了銅主要結(jié)合于蓖麻根細(xì)胞壁的半纖維素上。李芹等[55]的研究也表明,半纖維素Ⅰ是水稻根細(xì)胞壁蓄積鎘的主要場(chǎng)所。相反,周冉[56]和張涵[57]等分別報(bào)道了鎘和鋁主要結(jié)合在植物根細(xì)胞壁的果膠上。本研究通過(guò)差速離心法分離柑橘幼苗側(cè)根細(xì)胞壁多糖組分并分析各多糖組分的銅累積量。銅在柑橘側(cè)根細(xì)胞壁多糖組分的累積量表明,果膠是銅在柑橘側(cè)根細(xì)胞壁的主要結(jié)合位點(diǎn),其結(jié)合的銅濃度占細(xì)胞壁銅濃度的百分比高于98%。
研究表明,不同重金屬脅迫對(duì)細(xì)胞壁厚度的影響并不一致。例如,過(guò)量鉛[58]和鎘[59]使小麥根細(xì)胞壁變薄。相反,鎘毒害引起苔蘚[60]和鉤沙菜[61]細(xì)胞壁增厚。本研究發(fā)現(xiàn),過(guò)量銅處理的柑橘幼苗根細(xì)胞壁厚度顯著增加。細(xì)胞壁多糖組分富含大量可與金屬離子結(jié)合的官能團(tuán),細(xì)胞壁的增厚可結(jié)合更多的金屬離子,從而降低重金屬對(duì)植物的毒害。本研究還發(fā)現(xiàn),過(guò)量銅處理的兩個(gè)柑橘品種幼苗側(cè)根細(xì)胞壁提取率(圖1)較對(duì)照組都顯著提高。可見(jiàn),細(xì)胞壁受過(guò)量銅影響做出了應(yīng)激反應(yīng),通過(guò)增加細(xì)胞壁的量以結(jié)合更多的銅,對(duì)細(xì)胞內(nèi)部進(jìn)行保護(hù)。細(xì)胞壁含量提高且厚度增加可能是Kulikova等[62]提出的過(guò)量銅引起植物根尖膨大的主要原因。兩個(gè)柑橘品種比較發(fā)現(xiàn),過(guò)量銅處理的沙田柚側(cè)根細(xì)胞壁銅含量(表2)、細(xì)胞壁提取率(圖1)與厚度(圖3)都顯著高于雪柑。因此,柑橘幼苗側(cè)根細(xì)胞壁特性可能是影響兩個(gè)柑橘品種幼苗銅分布特征與耐銅差異的關(guān)鍵因素。
細(xì)胞壁果膠、纖維素和蛋白質(zhì)等組分對(duì)銅具有較高的吸附和固定能力,主要源于羥基、羧基和氨基官能團(tuán)[63]。FTIR譜圖是植物細(xì)胞壁各主要化學(xué)成分的綜合反映,可以根據(jù)相應(yīng)紅外光譜吸收峰找到對(duì)應(yīng)的官能團(tuán)。特征峰吸光度的比值越高,暗示該官能團(tuán)的含量越高,當(dāng)官能團(tuán)參與銅的吸附時(shí),根細(xì)胞壁上官能團(tuán)的特征吸收峰會(huì)發(fā)生不同程度的位移和強(qiáng)度的變化,官能團(tuán)沒(méi)有參與銅的吸附時(shí),其特征吸收峰的位移不變或變化較小[42]。在本研究中,兩個(gè)柑橘品種的特征吸收峰向低頻或高頻的位移程度表明,果膠、纖維素和蛋白質(zhì)處的吸光值與A2921比值大小變化顯著,這與劉思怡等[64]和曹升等[65]在茶樹(shù)上的研究報(bào)道類(lèi)似。
4 結(jié)論
相比對(duì)照,過(guò)量銅處理顯著提高了兩個(gè)柑橘品種幼苗各部位的銅含量。組織水平上,過(guò)量銅主要富集在柑橘苗側(cè)根;亞細(xì)胞水平上,過(guò)量銅主要結(jié)合于側(cè)根可溶性部分和細(xì)胞壁果膠組分。品種比較表明,過(guò)量銅處理的沙田柚側(cè)根銅含量、側(cè)根細(xì)胞壁銅含量以及側(cè)根細(xì)胞壁果膠銅含量均顯著高于雪柑。提高細(xì)胞壁含量、增厚細(xì)胞壁以及促進(jìn)細(xì)胞壁官能團(tuán)對(duì)銅的吸附可能是沙田柚耐銅性強(qiáng)于雪柑的主要原因。
