基于線粒體Cyt b基因的雅魯藏布江中游短尾高原鰍遺傳多樣性研究
2022-07-29 05:05:12劉艷超馬聰敏李小兵
淡水漁業 2022年4期
鞏 政,劉艷超,馬聰敏,李小兵
(1.棗莊學院生命科學學院,山東棗莊 277160;2.水利部中國科學院水工程生態研究所水利部水工程生態效應與生態修復重點實驗室,武漢 430072;3.西藏自治區高原生物研究所,拉薩850000;4.西南大學水產學院,重慶 402460)
雅魯藏布江是西藏地區最大的河流,也是世界上海拔最高的大河,它發源于喜馬拉雅西段北麓的杰馬央宗冰川,由西向東橫貫西藏南部,隨后沿喜馬拉雅山東麓急轉南下,在巴昔卡流出國境進入印度。雅魯藏布江全長2 229 km,流域面積達239 228 km,源頭至里孜段為其上游,派鎮以下至巴昔卡為下游,從里孜至派鎮全長1 340 km的河段則屬于雅魯藏布江中游。中游江段流域環境獨特且復雜多樣,孕育出異齒裂腹魚(′)、巨須裂腹魚()和黑斑原鮡()等雅魯藏布江流域特有物種。然而,近十幾年來,作為西藏人口和經濟社會發展最為密集的區域,在人類活動干擾、外來魚類入侵、水電開發和全球氣候變化等因素的影響下,雅魯藏布江中游魚類的生活史特征、遺傳多樣性及群落結構等方面均發生重要變化,引發廣泛關注。
高原鰍屬()隸屬于鯉形目(Cypriniformes)條鰍科(Nemacheilidae),現知FishBase數據庫中已收錄超過180個有效種或亞種,是條鰍科中最大的類群,并與鯉形目鯉科的裂腹魚類和鲇形目鮡科的鰋鮡魚類共同組成了適應青藏高原高寒環境的三大魚類類群。已知分布于雅魯藏布江流域的高原鰍屬魚類包括短尾高原鰍()、東方高原鰍()、細尾高原鰍()、異尾高原鰍()、斯氏高原鰍()和西藏高原鰍()等6種。短尾高原鰍的模式產地為青海省達布遜鹽湖水系,目前被認為分布于包括雅魯藏布江在內的青藏高原及其毗鄰的各水系中,相關魚類資源調查研究表明短尾高原鰍廣泛分布于雅魯藏布江中游流域且種群規模較大。然而,目前對于該流域短尾高原鰍包括遺傳多樣性在內的基礎生物學研究報道十分匱乏。
脊椎動物的線粒體DNA(mtDNA)基因組分子結構簡單,進化速率較核基因快且嚴格遵循母系遺傳,在遺傳多樣性分析中可選取其中特定基因片段,基于堿基位點的突變分析其遺傳變異的程度并推測群體的遺傳結構。線粒體細胞色素b(b)基因以其進化速率較快和易于擴增測序等優點已被廣泛應用于種內或種間的遺傳變異分析中。本實驗通過對雅魯藏布江中游不同支流中生活的短尾高原鰍b基因序列的測定,分析其遺傳多樣性現狀,檢測群體間分化水平,追溯其種群歷史動態,進而為該物種提供生物學資料,為物種保護提供理論依據。
1 材料與方法
1.1 樣本采集
分別于2020年5-6月和2021年8月在雅魯藏布江中游10條支流中采集短尾高原鰍樣本,共計125尾。剪取樣本背部右側肌肉或將其整體固定于無水乙醇中,帶回實驗室后保存于-20 ℃冰箱中備用。采樣點的樣本量及分布信息見表1和圖1。
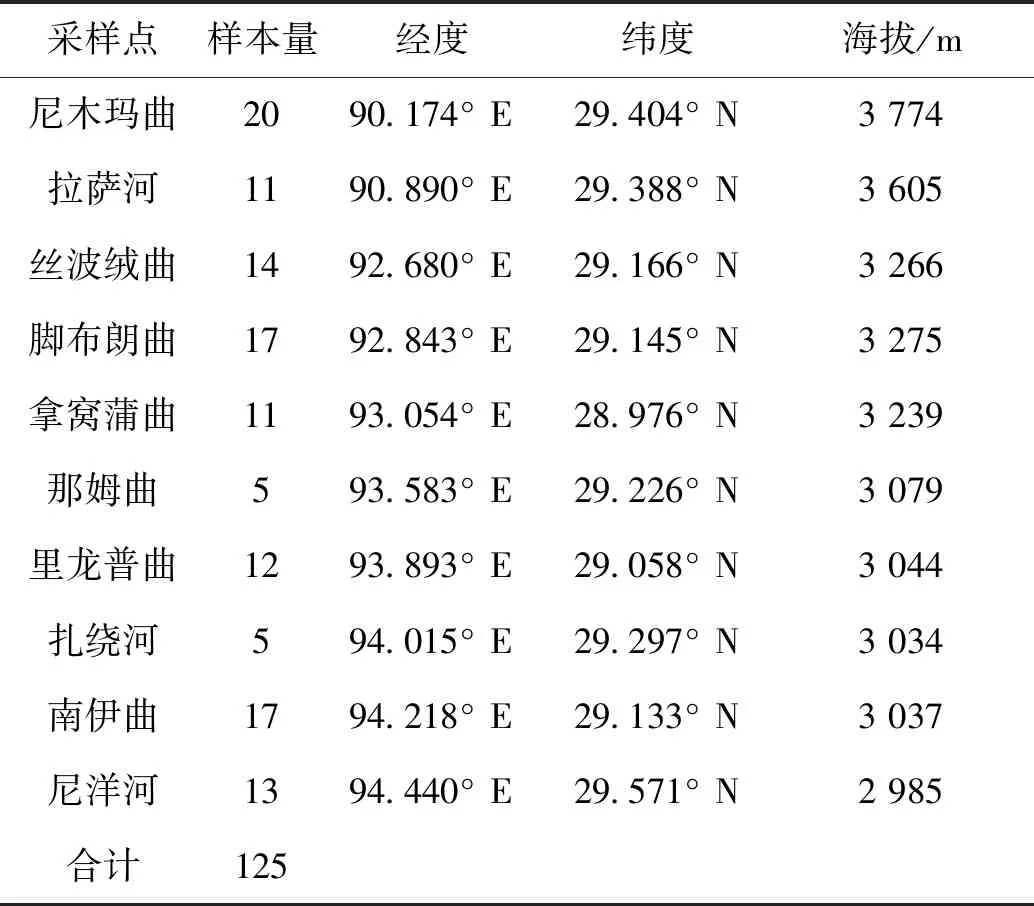
表1 雅魯藏布江中游短尾高原鰍的樣本采集信息Tab.1 Sample data of T.brevicauda collected from the middle Yarlung Tsangpo River
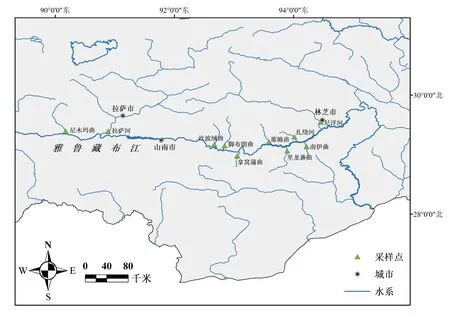
圖1 雅魯藏布江中游短尾高原鰍的采樣點示意圖Fig.1 Map showing the sampling sites of T.brevicauda in the middle Yarlung Tsangpo River
1.2 DNA提取和PCR擴增
DNA提取采用略微改進的高鹽抽提法,分子標記b基因擴增和測序的引物為L14724和H15915。PCR反應體系為30 μL,包括3 μL的10 × Buffer,1.5 μL dNTPs,上下游擴增引物各1 μL,0.25 μL的TaqDNA聚合酶和30~50 ng DNA模板,然后加入雙蒸水標定體積。PCR反應條件如下:95 ℃預變性4 min;進入反應循環:95 ℃變性45 s,54~56 ℃退火45 s,72 ℃延伸1 min,共循環35次;72 ℃延伸10 min。PCR產物使用8%的瓊脂糖電泳檢測目標條帶,擴增成功后送至武漢擎科生物有限公司進行純化和雙向測序。
1.3 數據分析
測定的短尾高原鰍b序列使用Clustal X v2.0軟件進行比對,并在SEAVIEW軟件中參照序列峰圖進行人工校正。使用MEGA v6.0軟件將基因序列翻譯成氨基酸序列進行勘誤,并計算序列中堿基組成、變異位點以及轉換()顛換()比值等參數。利用DnaSP v5.1軟件分析短尾高原鰍的遺傳多樣性指數,包括單倍型數目()、單倍型多樣性()和核苷酸多樣性()等。
采用貝葉斯推斷(Bayesian inference,BI)和最大似然法(Maximum likelihood,ML)分別重建單倍型的系統發育樹。首先在PhyloSuite v1.2軟件PartitionFinder v2.1模塊中基于修正的赤池信息準則(corrected Akaike information criterion,AICc)和密碼子分區篩選核苷酸替換模型,得到的最佳模型用于下一步系統樹的構建中。貝葉斯樹的構建在MrBayes v3.1軟件中實現;最大似然樹的構建在RaxML v8.0軟件中實現。利用popart 1.7軟件中的TSC方法繪制單倍型的進化網絡圖,分析單倍型之間的進化關系。使用Arlequin v3.5軟件檢測不同地理群體間遺傳分化指數(),基于分子方差分析(analysis of molecular variance,AMOVA)估算群體遺傳結構和地理群體遺傳變異的分布。
種群歷史動態分析使用Arlequin v3.5軟件對各群體和總體進行中性檢驗,計算Tajima′s和Fu′s值兩項參數;錯配分布分析基于粗糙指數(Harpending′s raggedness index,)和偏差平方和()衡量結果與預期模型是否相符合及其顯著性水平。使用BEAST v1.8軟件進行種群動態的貝葉斯天際線(Bayesian Skyline Plot,BSP)分析:采用鯉形目魚類常用的進化速率(0.01/ma),堿基替換模型選擇GTR模型,Number of Gamma Categories設為6,選擇Coalescent Bayesian Skyline模型作為點圖模型,馬爾科夫鏈(MCMC)的長度為100 000 000;最后在Tracer v1.5軟件中構圖。
2 結果
2.1 Cyt b基因序列堿基組成與變異
對125尾短尾高原鰍的線粒體b基因序列進行比對校正后,獲得長度為1 140 bp的序列集,起始密碼子為ATG,中止密碼子為GCT,未檢測到堿基的插入或缺失。
基于短尾高原鰍的b序列集檢測到堿基變異位點共95個,其中70個為簡約信息位點;全部個體的平均堿基組成為A=27.0%,T=29.4%,C=26.9%,G=16.7%,A+T的含量(56.4%)高于C+G(43.6%),堿基組成具有強烈的反G偏倚(bias),其中堿基G在密碼子第三位的含量尤其低(10.7%);序列的轉換和顛換均未達飽和,轉換數明顯大于顛換數,平均值為10.26。
2.2 遺傳多樣性
從125尾個體中檢測到52個單倍型,總體單倍型多樣性與核苷酸多樣性分別為0.953±0.010和0.010 51±0.001 04。具體而言,單倍型多樣性最低的是扎繞河群體(0.600±0.175),最高的是那姆曲群體(0.900±0.161);核苷酸多樣性最低的是尼洋河群體(0.001 87±0.000 40),最高的是那姆曲群體(0.019 65±0.003 36)。短尾高原鰍各地理群體的序列變異情況和遺傳多樣性指數見表2。
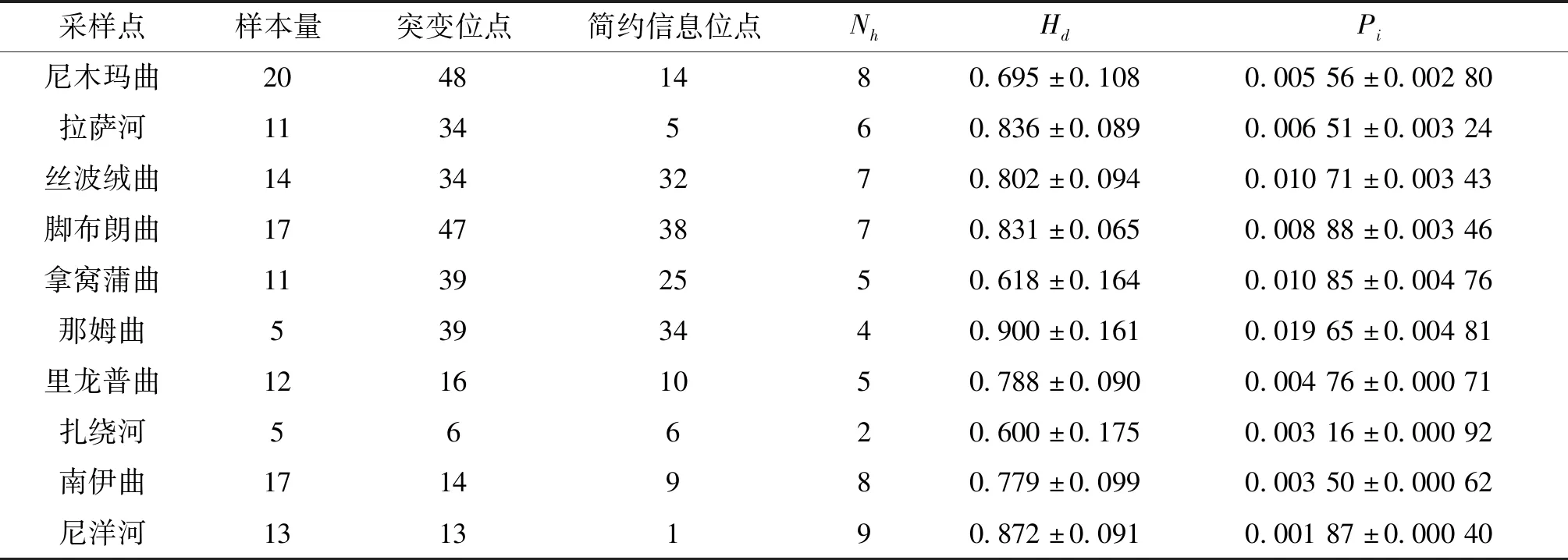
表2 基于Cyt b基因序列的雅魯藏布江中游短尾高原鰍的遺傳多樣性Tab.2 Genetic diversity of T.brevicauda in the middle Yarlung Tsangpo River based on Cyt b gene sequences
以分布于雅魯藏布江中游的另外兩種高原鰍屬魚類:細尾高原鰍和異尾高原鰍作為外類群,采用BI法和ML法重建的短尾高原鰍單倍型系統發育樹的拓撲結構基本一致,在系統樹中所有單倍型可被劃分為5支譜系(圖2)。其中,譜系I規模最大,其中包含的33個單倍型分布于除拿窩蒲曲和絲波絨曲以外的其他所有群體中;此外,譜系I中還包含若干亞譜系。譜系II中包含的5個單倍型分布于絲波絨曲、腳布朗曲和里龍普曲3個群體中。譜系III中包含的6個單倍型僅分布于尼洋河群體中。譜系IV中包含的5個單倍型分布于尼木瑪曲、拉薩河和絲波絨曲3個群體中。譜系V中包含的3個單倍型分布于尼木瑪曲、腳布朗曲、拿窩蒲曲和那姆曲4個群體中。
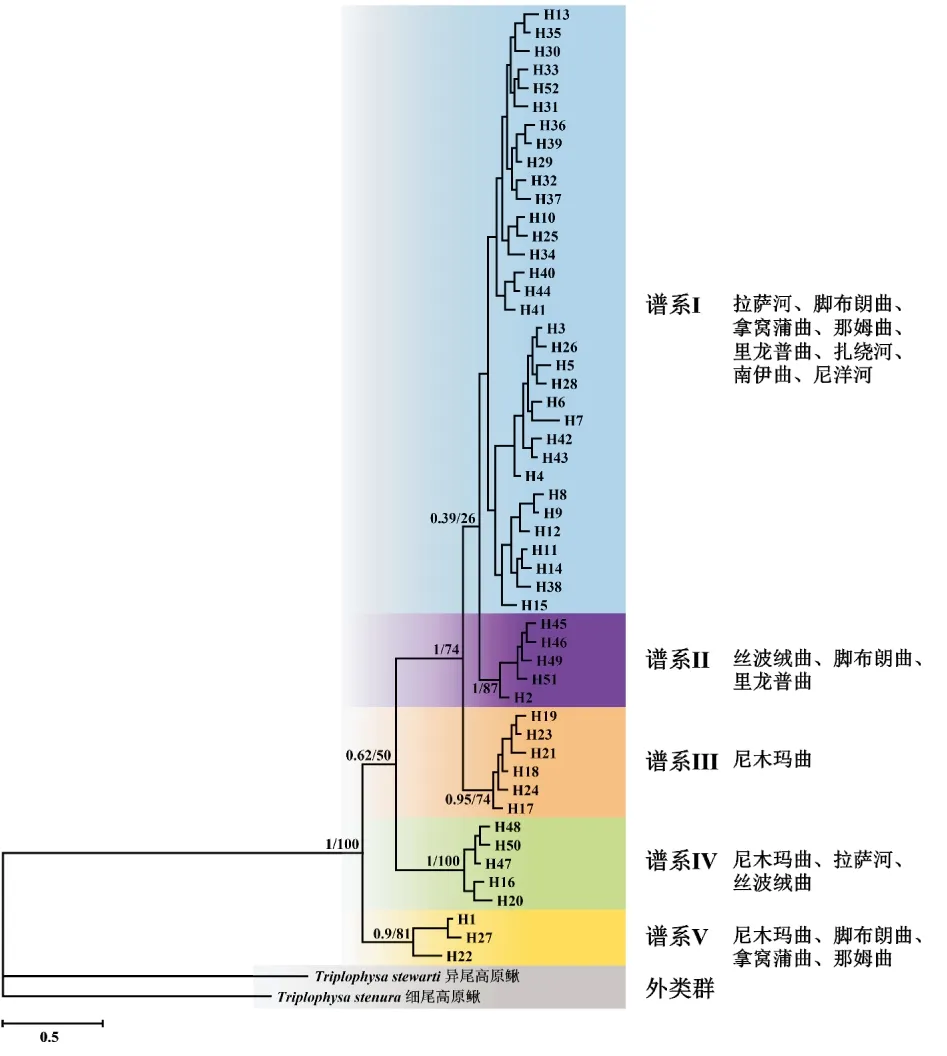
圖2 基于Cyt b基因序列單倍型構建的雅魯藏布江中游短尾高原鰍的系統發育樹Fig.2 Phylogenetic tree of T.brevicauda in the middle Yarlung Tsangpo River based on Cyt b gene sequence haplotypes
基于TCS法構建的單倍型進化網絡圖顯示:52個單倍型中有47個單倍型為單一群體所獨享,僅有少數單倍型(例如H2)為多個群體所共享(圖3)。系統發育分析檢測出的5支譜系所屬單倍型之間堿基突變大都超過十步;而在譜系內部相鄰單倍型之間往往僅經歷一到兩步的堿基突變,多個單倍型通過缺失的中間單倍型連接。
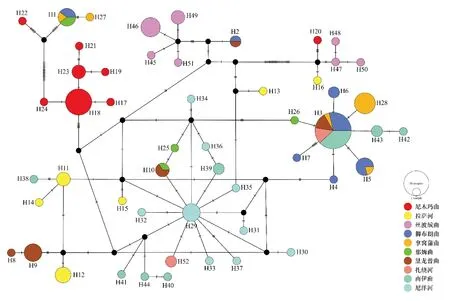
圖3 基于Cyt b基因序列的雅魯藏布江中游短尾高原鰍單倍型的進化網絡圖Fig.3 Evolutionary network of T.brevicauda in the middle Yarlung Tsangpo River inferred from Cyt b gene haplotype frequencies
2.3 群體間遺傳分化
不同地理群體兩兩配對計算得到的范圍為-0.057~0.605(表3),大部分群體之間的遺傳分化指數呈現示出顯著或極顯著水平,少數群體間未檢測出遺傳分化。其中,尼木瑪曲群體與其他群體之間的最大(0.467~0.605)且均為顯著或極顯著水平;拿窩蒲曲群體、扎繞河群體與其他群體之間遺傳分化指數最小(-0.016~0.525,-0.057~0.566),且部分群體間的檢測結果未達到顯著性水平。
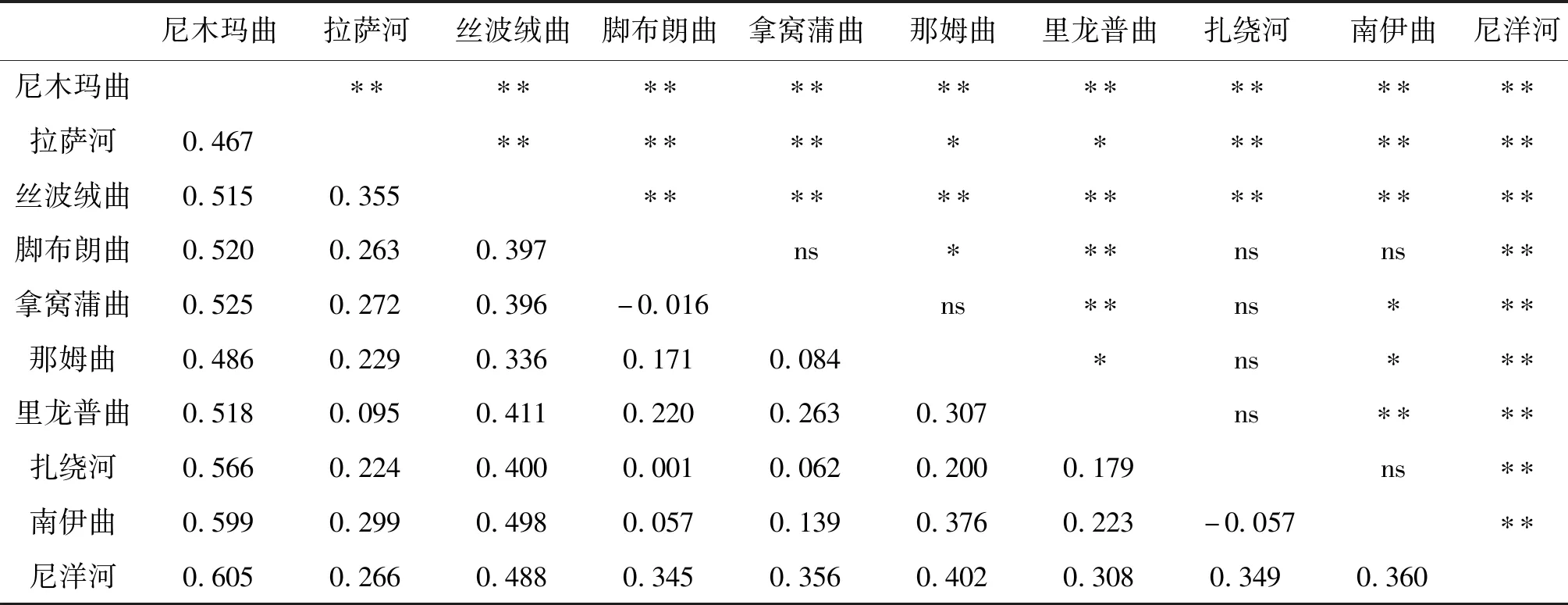
表3 雅魯藏布江中游10個地理群體短尾高原鰍的FSTTab.3 The FST between 10 populations of T.brevicauda in the middle Yarlung Tsangpo River
AMOVA分析結果顯示,在不分組的情況下遺傳變異主要來自于群體內部,占比為62.1%;群體間的遺傳變異相對較小,占比為37.9%,表明遺傳多樣性主要產生于個體間的遺傳變異,群體間的分化亦占有一定比例。根據楊漢運等對雅魯藏布江中游不同江段河流形態的劃分,將10個群體劃分為3組(尼木瑪曲和拉薩河群體為一組,分布于曲水寬谷段;絲波絨曲、腳布朗曲、拿窩蒲曲、那姆曲和里龍普曲群體為一組,分布于加查峽谷段;扎繞河、南伊曲和尼洋河群體為一組,分布于米林寬谷段),此時得到的組間變異占9.23%,組內群體間變異占30.28%,群體內變異占60.89%(表4)。
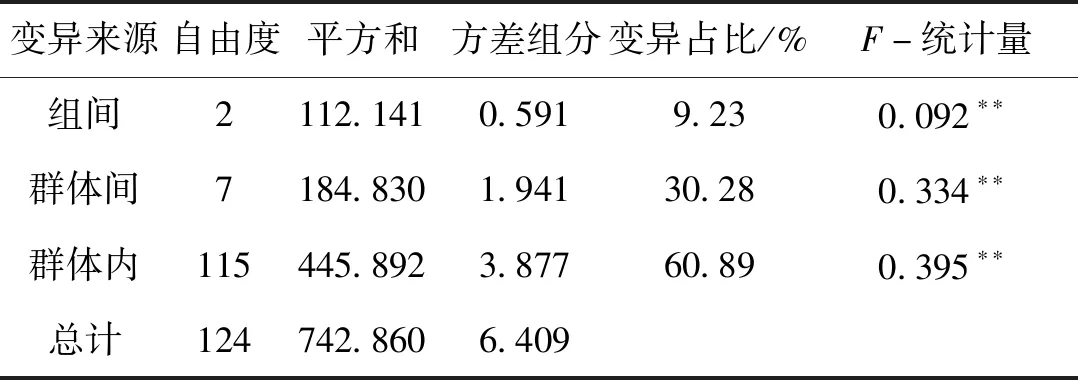
表4 雅魯藏布江中游10個地理群體短尾高原鰍的AMOVA分析Tab.4 Analysis of molecular variance analysis among ten populations of T.brevicauda in the middle Yarlung Tsangpo River
2.4 種群歷史動態
中性檢驗與核苷酸錯配分析的結果見表5,中性檢驗的結果顯示尼木瑪曲、拉薩河、尼洋河群體及總體的Tajima′值或Fu′s值均為顯著或極顯著的負值,而其他群體的Tajima′值或Fu′s值檢驗結果均不顯著。核苷酸錯配分析顯示,絲波絨曲、拿窩蒲曲、那姆曲、里龍普曲和扎繞河群體檢測出具有顯著或極顯著水平的和,其他群體的參數檢驗均未達到顯著水平。對10條支流短尾高原鰍總體進行BSP分析,結果顯示短尾高原鰍在1.12 kya出現過快速的種群擴張,隨后又發生了急速的種群收縮(圖4)。
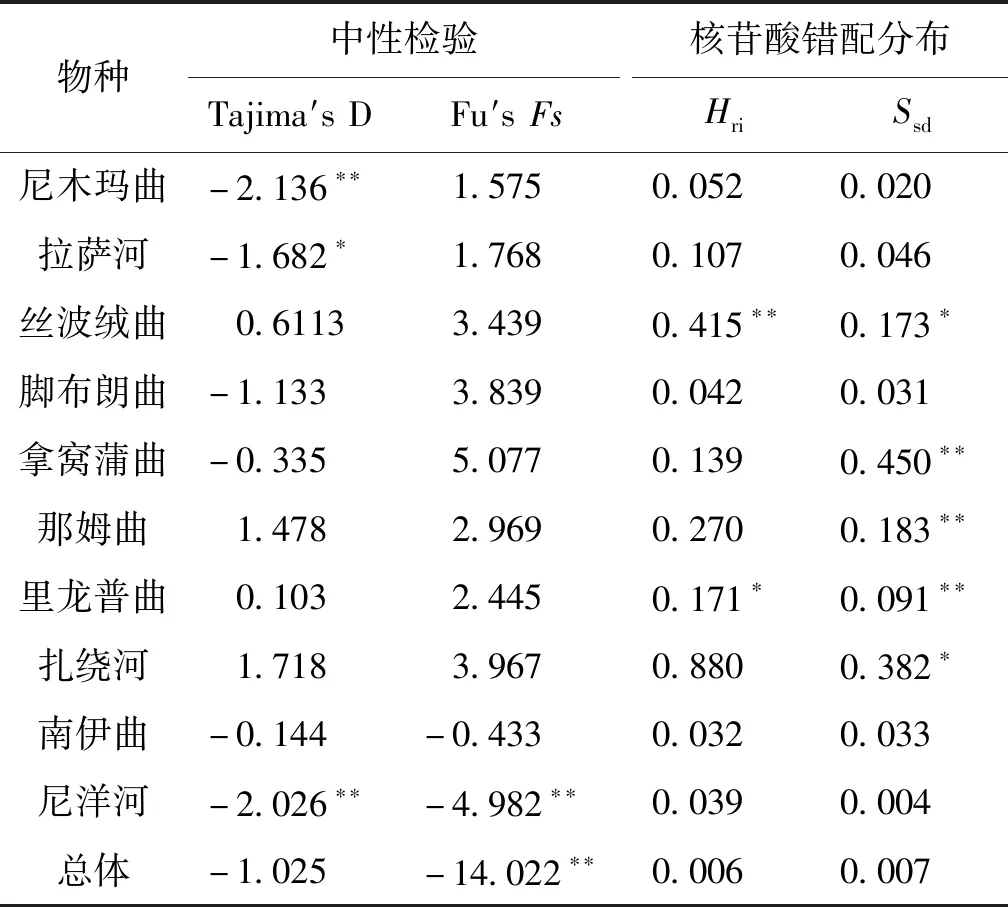
表5 雅魯藏布江中游短尾高原鰍地理群體的中性檢驗和核苷酸錯配分析Tab.5 Indices of neutral test and mismatch distribution analysis for T.brevicauda in the middle Yarlung Tsangpo River
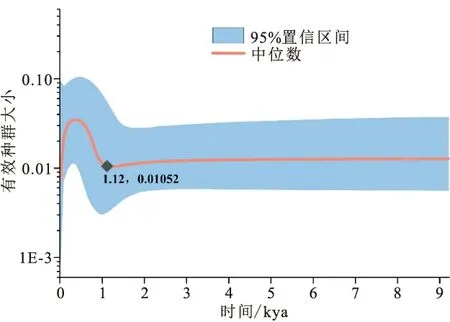
圖4 基于Cyt b基因序列的雅魯藏布江中游短尾高原鰍的貝葉斯天際線分析Fig.4 Bayesian skyline plot analysis based on Cyt b gene sequences for T.brevicauda in the middle Yarlung Tsangpo River
3 討論
3.1 遺傳多樣性與群體分化
單倍型多樣性指數()與核苷酸多樣性指數()被認為是衡量物種或群體遺傳多樣性水平高低的重要指標。本研究中短尾高原鰍總體及尼木瑪曲、拉薩河、絲波絨曲、腳布朗曲、拿窩蒲曲和那姆曲群體≥0.5,≥0.005,表明這些地理群體和總體的遺傳結構較為穩定;而里龍普曲、扎繞河、南伊曲和尼洋河群體≥0.5,<0.005,表明這些群體在歷史上可能遭遇過遺傳瓶頸后又經歷了種群擴張。同雅魯藏布江中游異齒裂腹魚等其他魚類的基于b序列的遺傳多樣性指數相比,短尾高原鰍總體的單倍型多樣性和核苷酸多樣性均處于較高水平,遺傳結構較為穩定。
單倍型系統發育關系與進化網絡圖顯示,短尾高原鰍單倍型大致聚為5支譜系且未按照地理分布聚類,即10個地理群體的單倍型混雜分布于5支譜系中,不符合各支流群體的空間分布格局(個別譜系僅由單一群體的單倍型構成:譜系III包含的單倍型僅分布于尼洋河群體中)。90.4%的單倍型僅出現在單一群體中,不同群體間共享的單倍型極少,這些暗示著雅魯藏布江中游的短尾高原鰍群體具有共同的祖先起源或曾經發生過充分的遺傳交流,在較近的時期內發生了種群收縮和遺傳分化。此外,單倍型進化網絡圖還顯示不同譜系的單倍型群組不具有占絕對優勢的進化網絡中心,僅在譜系局部存在少數單倍型呈中心放射狀分布,其他單倍型由它突變演化而來;例如譜系I中的H3和H29,推測這些單倍型為發揮過“奠基者效應”的古老單倍型。
遺傳分化指數()可在一定程度上指示群體間基因交流和遺傳漂變的程度,是反映群體進化歷史的重要參數。尼木瑪曲群體與其他9個群體之間均存在極顯著水平的高度分化(范圍為0.467~0.605);部分群體之間未檢測具有顯著水平的遺傳分化,主要出現在拿窩蒲曲和扎繞河群體與其他少數群體之間,原因可能是這兩個群體樣本數量較少,后期需要增加樣本量進行驗證。AMOVA分析同樣表明群體間的遺傳變異占據相當比例,支持遺傳分化的結論。短尾高原鰍在雅魯藏布江中游不同支流群體間普遍存在高度遺傳分化,究其原因可能是由于短尾高原鰍是一種小型定居型魚類,沒有遷移洄游的習性,不同地理群體間基因交流的機會很少,局域種群在長期獨立進化過程中朝著不同方向發生遺傳漂變。
3.2 種群歷史動態
中性檢驗顯示部分群體的Tajima′值和Fu′s值均為顯著的負值,偏離了中性突變,表明三種魚類在近期應該經歷過種群擴張事件,對應的核苷酸錯配分析中粗糙度指數和偏方差的檢驗結果均不顯著,支持種群擴張的推斷。對短尾高原鰍總體進行BSP分析的結果顯示其在很近的歷史時期(約1.12 kya)經歷過急速的種群擴張和種群收縮。
種群的歷史動態對區域地質運動和氣候變化有著優良的指示作用。相關地質研究表明,青藏高原的隆升是決定雅魯藏布江河谷地貌形成的主要因素。雅魯藏布江中游寬谷與峽谷相間呈串珠狀分布,說明雅魯藏布江流域的地質構造復雜多變,具體表現在峽谷江段主要是由地殼上升塑造而成,而寬谷江段主要是由局部地殼下降所導致。本研究的10條支流從上至下依次分布于曲水寬谷、加查峽谷和米林寬谷段,各自經歷了不同類型和強度的地質運動,進而影響著各江段內的河流形態和水系格局。同時,全新世以來青藏高原出現過多次明顯的冰期和間冰期的冷暖波動,且平均氣溫變化幅度以西藏南部地區為最大。末次冰盛期以后,青藏高原氣溫雖然整體上逐步回升,但仍存在多次波動調整,氣候依舊不穩定;例如在最近的2 ky,青藏高原先后出現了8次暖期和7次冷期。
作為一種適應高原環境的小型魚類,短尾高原鰍主要棲息于雅魯藏布江各支流中,遷移能力弱,對于分布區內的冷暖波動、水系變遷等環境擾動的響應尤為敏感。結合種群動態分析結果,我們認為末次冰期以來青藏高原南部的氣候波動可能通過改變短尾高原鰍在不同支流中的適合度,進而影響其種群動態;同時推測地質活動導致的水系變遷也是影響短尾高原鰍種群波動的重要因素。
3.3 物種保護建議
本研究基于線粒體b基因序列分析了雅魯藏布江中游短尾高原鰍的遺傳多樣性和群體遺傳結構,結果顯示各地理群體呈現較高的遺傳多樣性,不同地理群體間高度分化,共享單倍型極少。隨著雅魯藏布江干流多座梯級水電站的建設運行,不同河段支流間的群體進行基因交流變得更加困難。因此,為了防止小種群遺傳資源的喪失,在設置保護單元時應該針對不同支流進行分別管理和保護。同時,作為雅魯藏布江各支流中常見的高原鰍屬魚類,短尾高原鰍主要棲息于淺水礫石灘緩流水或靜水小潭等生境中。然而,隨著雅魯藏布江流域沿線公路、鐵路等工程的影響和外來物種的侵入,部分支流河道生境遭到破壞,外來魚類數量泛濫,短尾高原鰍種群生存受到嚴重威脅。
因此,我們建議對于諸如拉薩河、尼洋河等流量較大、生境類型豐富且已建有水利工程設施的支流,應當識別短尾高原鰍典型棲息地并劃定重點保護河段,對已遭破壞魚類保護河段進行生態修復,同時進行嚴格監管和防范外來魚類進入自然水體;對于其他流量較小、生境類型較為單一的支流,建議進行整體保護,不做水利水電開發,盡可能減少人類活動對河流生態環境造成的影響。此外,本研究也發現短尾高原鰍的種群數量曾受到較近歷史時期內氣候波動的影響。因此,還應加強對該物種遺傳多樣性和遺傳結構的跟蹤監測,及時完善和調整物種保護策略。
