404 Not Found
404 Not Found
牡丹種間遠緣雜交不親和的細胞學與生理機制研究
張翔宇,賈文慶,何松林,邱永杰,王喬健,胡 緩,史來琨,劉會超
(河南科技學院園藝園林學院,河南 新鄉 453003)
遠緣雜交是新優種質培育的主要途徑,野生種質與栽培種進行遠緣雜交在改良栽培品種以及培育新異種質中起著重要作用[1-3],如我國小麥育種專家李振聲團隊利用小麥(Triticum aestivum L.)與長穗偃麥草(Elytrigia elongata (Host) Nevski)雜交培育出抗病性強的‘小偃6號’等系列遠緣雜交小麥品種,成功解決抗“小麥癌癥”條銹病的世界育種難題[4],Moshe Feldman和Emest R.Sears也曾提出“未來的谷物改良寄希望于野生基因資源的利用”[5]。牡丹(Paeonia suffruticosa Andr.)為世界著名花卉,也是中國的一張名片,在中國影響之深遠是其他花卉所不可企及的;但牡丹現有栽培品種中純正花色少,黃色、橙色花缺乏,株型較低,大多對白粉病易感。長期以來,由于牡丹遠緣雜交不親和或親和性差,牡丹育種多集中在品種群內進行常規雜交,遺傳基礎狹窄和多樣性親本資源的缺乏成為了限制牡丹育種取得突破的瓶頸。
栽培品種中,由楊山牡丹(P.ostii T.Hong et J.X.Zhang sp.nov.)演化出的‘鳳丹’(P.ostii‘Feng Dan’)植株高大、生長勢強、花量大,生態適應性極廣泛,是雜交育種最常用的親本[6-7];牡丹野生種保康牡丹(Paeonia baokangensis Z.L.Dai et T.Hong sp.Nov.)[8],花桃紅,色澤純正、明亮,花期長,抗白粉病,是亟待開發的優良野生資源之一。解析牡丹‘鳳丹’與保康牡丹遠緣雜交的不親和性機制,對于克服受精障礙,完善牡丹遠緣雜交技術體系,培育花色純正、抗白粉病的牡丹新品種具有重要意義。
植物遠緣雜交研究發現,花粉萌發及花粉管生長情況、內源激素水平、保護酶活性及可溶性蛋白含量等是遠緣雜交親和性強弱的重要指標。郭朋輝等[9]在紫斑百合(Lilium nepalense D.Don)遠緣雜交親和性研究中發現,紫斑百合遠緣雜交過程中多出現受精前障礙,胼胝質沉積,花粉管扭曲纏繞,阻礙伸長生長是其受精前障礙的主要表現;王保成等[10]在蕓芥(Eruca sativa Mill)親和性研究中發現,SOD、POD及CAT三種保護酶的活性變化與自交親和基因的調控相關;楊曉玲等[11]在百合(Lilium brownie F.)親和性研究中發現,高水平的IAA、GA3、ZR對花粉在柱頭的識別黏附及萌發有促進作用;Kovaleva L等[12]表明,ABA含量較高是導致矮牽牛(Petunia hybrida L.)授粉不親和的原因;牡丹與芍藥(Paeonia lactiflora Pall.)雜交后也顯示相關保護酶活性及內源激素含量與授粉親和性相關[13]。目前,有關牡丹種間雜交不親和機制尚未見報道,基于此,本研究以牡丹‘鳳丹’為母本,以野生保康牡丹為父本進行了種間雜交,探討花粉萌發、花粉管生長、雌蕊保護酶活性、可溶性物質及內源激素含量變化與遠緣雜交不親和性的關系,以期為解析牡丹種間遠緣雜交不親和性機制以及完善牡丹遠緣雜交技術體系提供理論及試驗依據。
1 材料與方法
1.1 材料
母本‘鳳丹’種植于河南科技學院牡丹資源圃,父本保康牡丹采自洛陽國家牡丹園野生資源圃,二者植株健壯,均可正常開花結實。于露色期對母本材料進行去雄,掛上標簽并做隔離處理,4 d后用準備好的花粉進行人工授粉。
1.2 花粉萌發及花粉管生長熒光觀察
分別于授粉后1、3、6、8、12、24、36、48、72、96 h,取雌蕊10~15個(分別取自不同花朵)迅速放入卡諾固定液中,固定24 h以上轉入70%酒精于4 ℃ 冰箱存放,之后用8 mol·L?1NaOH 60 ℃條件下軟化4 h,漂洗后于0.5%苯胺藍染液避光染色6 h以上,壓片并于熒光顯微鏡進行觀察、拍照(圖1A~I及L×40倍;J及K×100倍)。
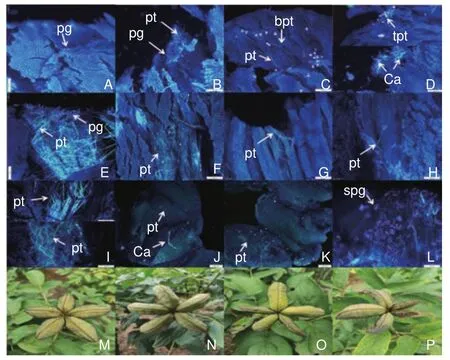
圖1 花粉萌發及花粉管生長熒光觀察Fig.1 Fluorescence observation of pollens germination and pollen tubes growth
1.3 生理指標測定
取未授粉及授粉后1、3、6、8、12、24、36、48、72、96 h的雌蕊,每處理0.2 g,重復3次,液氮速凍后于?80 ℃冰箱中保存。分別采用氮藍四唑(NBT)光還原法測定超氧化物歧化酶(SOD)活性,硫代巴比妥酸(TBA)法測定丙二醛(MDA)含量,考馬斯亮藍G-250法測定可溶性蛋白含量,苯酚法測定可溶性糖含量。
1.4 內源激素測定
取樣同1.3節,每處理0.3 g。采用酶聯免疫吸附法(ELISA),參照試劑盒附帶說明,分別測定吲哚乙酸(IAA)、赤霉素(GA3)、異戊烯基腺苷(IPA)、脫落酸(ABA)及茉莉酸(JA)含量。
2 結果與分析
2.1 花粉萌發及花粉管生長熒光顯微觀察
遠緣雜交與自交授粉后不同時期雌蕊熒光顯微觀察如圖1,授粉后1 h,遠緣雜交授粉雌蕊花粉粒黏附極少,未見萌發(圖1A);自交授粉花粉粒黏附較多,花粉萌發花粉管生長正常(圖1B)。授粉6 h,遠緣雜交花粉粒黏附增多,花粉粒萌發,花粉管有扭曲、分枝現象,伴有大量胼胝質沉積(圖1C、D);自交雌蕊花粉管生長迅速,有序向花柱生長(圖1E)。授粉后12 h,遠緣雜交授粉花粉管多扭曲,部分伸長到柱頭1/2到2/3部分(圖1F);自交授粉花粉管生長正常,伸長到花柱部分(圖1G)。授粉36 h,遠緣雜交授粉個別花粉管伸長到子房(圖1H);自交授粉花粉管成束有序穿過花柱,伸長到子房(圖1I)。授粉72 h,遠緣雜交授粉少量花粉管伸長到胚珠,伴有胼胝質沉積(圖1J),柱頭上花粉粒大多開始干癟(圖1L);自交授粉花粉管正常生長到胚珠(圖1K)。后期觀察發現,自交授粉能正常結實,結實率高,種莢飽滿(圖1M、N);遠緣雜交授粉結實率低,種莢大多干癟(圖1O、P)。整體表明,遠緣雜交受精前障礙明顯,花粉粒萌發晚,花粉管內胼胝質沉積是其不親和原因。
2.2 自交與遠緣雜交授粉后雌蕊SOD活性變化
授粉后不同時間,自交與遠緣雜交授粉雌蕊SOD活性變化(圖2)表明:授粉后1 h,自交雌蕊SOD活性急速上升,遠緣雜交雌蕊活性稍有上升,此時自交雌蕊花粉萌發,花粉管生長迅速,遠緣雜交雌蕊未見花粉萌發;1~3 h自交雌蕊SOD活性繼續急速顯著上升(P<0.05),在3 h達到頂峰84.57 U·g?1,3 h之后活性稍有下降,此后保持在較高水平,并在72 h達到另一高峰75.05 U·g?1,此時花粉管伸入胚珠部分。遠緣雜交雌蕊SOD活性1~3 h同樣急速上升,在3 h達到一個峰點74.78 U·g?1,此時遠緣雜交雌蕊個別花粉萌發;3 h后稍有下降,在24 h達到另一峰值81.97 U·g?1,之后保持在較高水平。表明高活性SOD對花粉萌發及花粉管生長有促進作用,尤其在授粉前期,高活性SOD有助于花粉-柱頭的識別作用,對自交授粉花粉粒在柱頭的黏附萌發促進作用顯著。

圖2 授粉后雌蕊 SOD 活性及MDA 含量變化Fig.2 Changes of SOD activity and MDA content in pistils after pollination
2.3 自交與遠緣雜交授粉后雌蕊MDA含量變化
授粉后,自交與遠緣雜交授粉雌蕊不同時間MDA含量變化(圖2)表明:授粉后1 h,自交雌蕊MDA含量顯著上升(P<0.05),1 h達最大67.03 μmol·g?1,此時自交雌蕊花粉萌發,花粉管生長迅速;1 h之后MDA含量下降,并在8 h之后保持在較低水平,隨后波動不大,顯微特征表現為花粉管有序向花柱及子房生長,推測此時雌蕊內部達到平衡狀態,雌蕊自身適應花粉落在柱頭所帶來的刺激作用。遠緣雜交授粉雌蕊MDA含量在授粉后1~3 h急速下降,此時花粉未見萌發;6 h之后MDA含量開始急速顯著上升(P<0.05),12 h含量達到最大 72.87 μmol·g?1,此階段花粉管出現扭曲,并伴有胼胝質沉積,花粉管生長受阻,可能原因在于柱頭對外來花粉識別后排斥作用明顯,誘導MDA急速增加,進而打破雌蕊內部動態平衡,細胞功能受損;在授粉后24 h之后MDA快速下降,并在48 h之后保持穩定,推測此時雌蕊應激響應達到平衡,整體處于穩定狀態。
2.4 自交與遠緣雜交授粉后雌蕊可溶性蛋白、可溶性糖含量變化
授粉后不同時間,自交與遠緣雜交授粉雌蕊可溶性蛋白、可溶性糖含量變化(圖3)表明:自交與遠緣雜交授粉0~36 h,可溶性蛋白及可溶性糖含量均表現為波動上升趨勢,自交與遠緣雜交的可溶性蛋白含量分別在24、36 h達最大,分別為52.90、50.48 mg·g?1,可溶性糖含量分別在 12、8 h 達到最大,分別為 30.17、31.64 mg·g?1;授粉36 h后,二者含量相對穩定。由此說明,可溶性蛋白及可溶性糖在柱頭識別及應對外來刺激作用中起著重要作用,其中,可溶性糖較可溶性蛋白含量變化小,表明在柱頭識別及應對外來刺激作用中,可溶性蛋白起著主導作用。出現此現象可能的原因是,在柱頭識別階段,可溶性蛋白作為識別蛋白的重要組成,積極參與柱頭的識別作用。此外,授粉作為外來刺激,雌蕊內部平衡破壞,加速雌蕊代謝,進一步提高了可溶性蛋白及可溶性糖含量,而表現出升高的趨勢,隨著刺激減弱,二者含量也相對保持平穩。
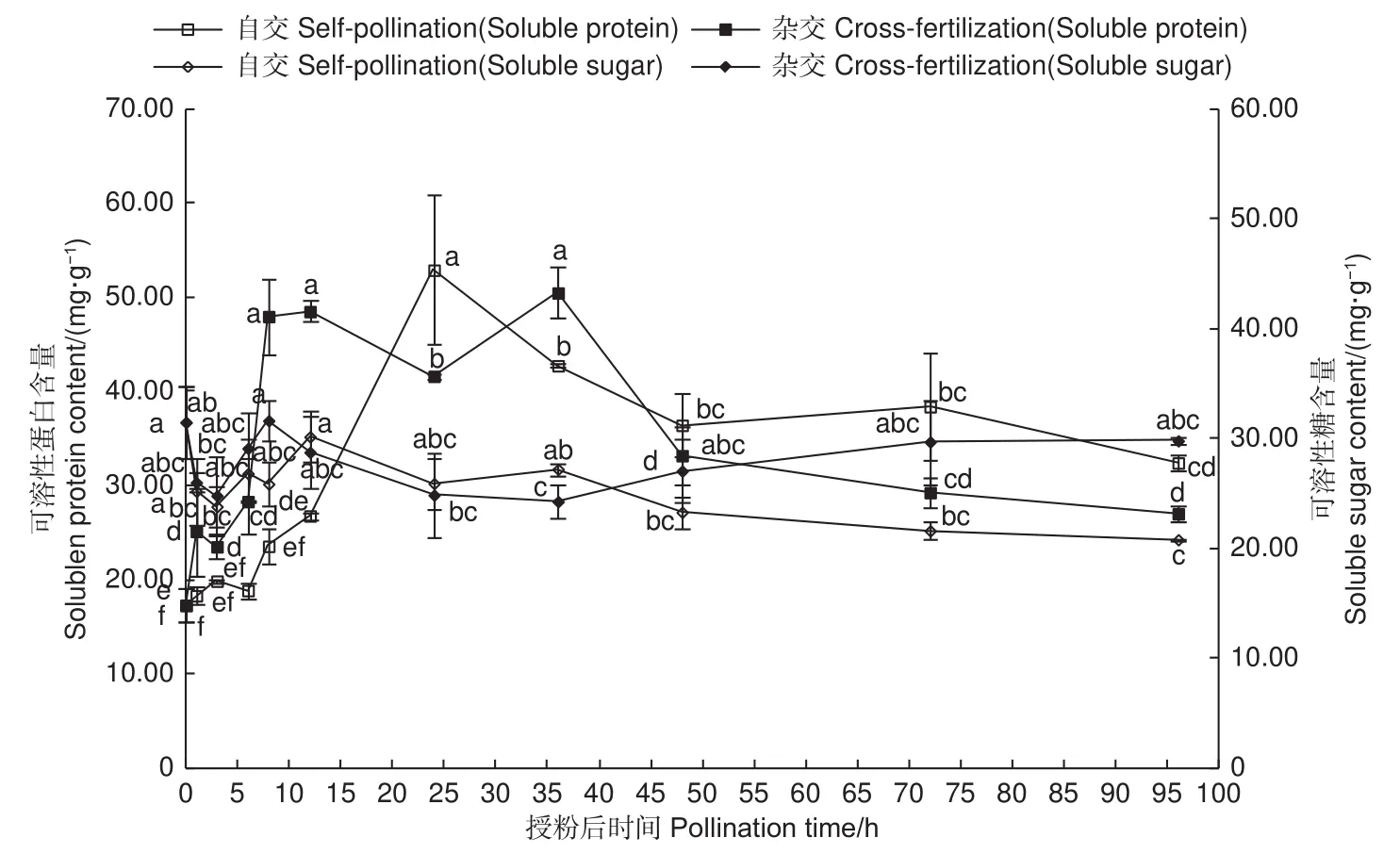
圖3 授粉后雌蕊可溶性蛋白含量及可溶性糖含量變化Fig.3 Changes of soluble protein content and soluble sugar content in pistils after pollination
2.5 自交與遠緣雜交授粉后雌蕊IAA、GA3含量變化
圖4表明:自交與遠緣雜交授粉雌蕊IAA、GA3含量變化整體表現為上升-下降-上升趨勢。整體上自交授粉雌蕊較遠緣雜交授粉雌蕊IAA含量高,尤其在授粉前期表現較顯著。自交授粉雌蕊IAA、GA3含量上升趨勢較遠緣雜交早,授粉后3~6 h自交授粉雌蕊IAA含量便急速顯著上升(P<0.05),并在 6 h 達最大 45.28 ng·g?1;自交授粉GA3含量在授粉后1~3 h快速上升,高于遠緣雜交授粉雌蕊,此階段自交授粉花粉萌發多,花粉管生長迅速。隨著遠緣雜交授粉GA3含量的升高,遠緣雜交授粉雌蕊花粉開始萌發,花粉管伸長生長。表明高水平IAA、GA3對花粉在柱頭的識別及花粉萌發、花粉管的生長起促進作用,IAA、GA3作為中間物質,起著傳遞生長信號的作用。
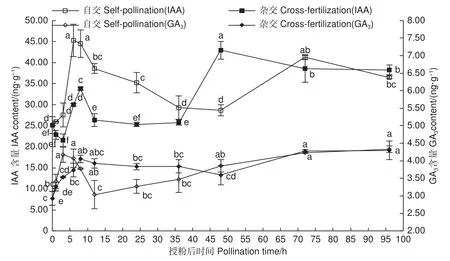
圖4 授粉后雌蕊 IAA 及GA3 含量變化Fig.4 Changes of IAA and GA3 content in pistils after pollination
2.6 自交與遠緣雜交授粉后雌蕊IPA含量變化
圖5表明:自交與遠緣雜交雌蕊授粉后IPA含量均表現出先升高后下降趨勢。授粉后1~36 h,自交授粉雌蕊IPA含量處于較高水平且保持相對穩定,遠緣雜交授粉雌蕊IPA含量呈波動上升,同樣保持在較高水平,其中,授粉后3~8 h,自交授粉雌蕊IPA含量高,顯微特征表現為花粉管迅速伸長生長,遠緣雜交授粉雌蕊IPA含量較低,顯微鏡觀察發現花粉管有胼胝質沉積及扭曲纏繞現象,推測高水平IPA對花粉管的伸長生長有促進作用;36 h之后均急速下降,之后保持相對穩定,花粉管生長也趨于穩定狀態,自交授粉含量依然高于遠緣雜交授粉。
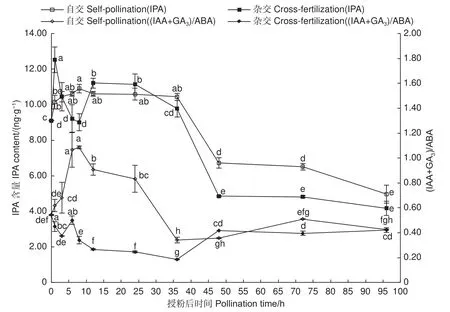
圖5 授粉后雌蕊 IPA 含量及(IAA+GA3)/ABA 變化Fig.5 Changes of IPA content and (IAA+GA3)/ABA in pistils after pollination
2.7 自交與遠緣雜交授粉后雌蕊(IAA+GA3)/ABA變化
圖5表明:自交授粉雌蕊(IAA+GA3)/ABA顯著高于遠緣雜交授粉雌蕊。授粉0~36 h,遠緣雜交授粉雌蕊(IAA+GA3)/ABA僅在授粉后6 h有上升趨勢,此時遠緣雜交授粉雌蕊處于花粉萌發、花粉管快速生長階段,此時ABA含量依然較高,花粉管扭曲嚴重;自交授粉雌蕊(IAA +GA3)/ABA僅在授粉后36~48 h較低,此時花粉管已伸入子房。
2.8 自交與遠緣雜交授粉后雌蕊ABA、JA含量變化
圖6表明:授粉0~24 h,自交與遠緣雜交授粉雌蕊ABA含量變化趨勢相反,自交授粉雌蕊ABA含量稍下降,遠緣雜交授粉雌蕊則急速顯著上升(P<0.05),并保持在較高水平,此階段自交授粉花粉萌發,花粉管生長迅速,遠緣雜交授粉花粉管扭曲嚴重,胼胝質沉積較多;授粉0~3 h,自交與遠緣雜交授粉雌蕊JA含量均上升,遠緣雜交授粉上升趨勢更顯著,此時自交授粉花粉少量萌發,遠緣雜交授粉花粉未見萌發;授粉3~12 h,JA含量均呈下降趨勢,此時花粉管處于快速生長階段;12 h之后,自交授粉雌蕊JA含量保持相對穩定,遠緣雜交授粉雌蕊JA含量再次上升,授粉72 h之后保持穩定,此時花粉管已伸長到胚珠部分;24~36 h期間,自交與遠緣雜交授粉雌蕊ABA含量均有大幅度上升,48 h之后呈現輕微下降趨勢,顯微鏡觀察發現花粉管伸長生長到子房部分。整個授粉受精期間,遠緣雜交授粉雌蕊ABA含量遠高于自交雌蕊,JA含量整體上也較自交授粉雌蕊高。推測在花粉-柱頭識別階段,JA含量的升高影響了花粉黏附及花粉的水合作用,進而阻礙了遠緣雜交雌蕊花粉的萌發;同時,伴隨著ABA含量的快速增加,高含量ABA致使遠緣雜交授粉花粉管胼胝質沉積較多,阻礙花粉管進一步伸長生長,表現出扭曲纏繞生長停滯。
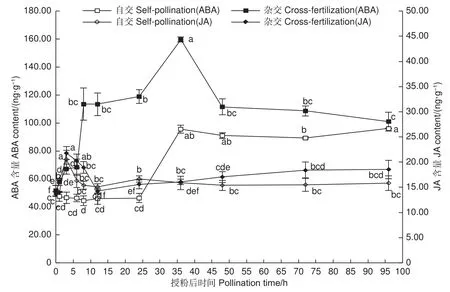
圖6 授粉后雌蕊 ABA 及JA 含量變化Fig.6 Changes of ABA and JA content in pistils after pollination
3 討論
受精前障礙是多數植物遠緣雜交天然生殖障礙的主要原因[14-15],相關研究表明,胼胝質大量積累是遠緣雜交不親和受精前障礙的主要細胞學原因之一,絨氈層的花粉外壁蛋白與柱頭乳突細胞質膜蛋白相互識別,親和性授粉,柱頭會分泌囊泡,提供花粉萌發所需的水合物質;不親和授粉,胼胝質產生積累在乳突細胞,擾亂花粉水合作用,進而抑制花粉萌發[16]。李辛雷等[17]在菊屬(Chrysanthemum L.)植物遠緣雜交研究中發現,不親和雜交組合柱頭上花粉粒黏附少,花粉萌發率低,花粉管生長異常,難以進入花柱;高山杜鵑(Rhododendron lapponicum (L.) Wahl.)遠緣雜交后,花粉管、花柱、胚珠等產生大量胼胝質,導致受精障礙而無法結實是其不親和的主要原因[18];牡丹芍藥遠緣雜交也因花粉管扭曲、纏繞、分枝、胼胝質沉積等表現為受精前障礙[19-21]。本試驗研究結果表明,‘鳳丹’與保康牡丹遠緣雜交受精前障礙明顯,胼胝質沉積在其中起著主導作用,原因可能是遠緣雜交授粉引起的脅迫反應,干擾了花粉管細胞發育過程中細胞壁內層胼胝質的正常合成,引起胼胝質異常累積,從而造成花粉管生長紊亂,出現扭曲纏繞現象。
授粉作為一種外來刺激,會在柱頭及子房內引起酶活性及滲透調節物質含量變化。SOD作為植物防御系統里重要的抗氧化酶,通過清除過量的活性氧來保護植物免受外界刺激產生的傷害,高水平SOD有利于花粉-柱頭的識別、黏附及花粉在柱頭的萌發[13]。吳能表等[22]在對甘藍(Brassica oleracea var.capitata L.)雜交授粉研究中發現,授粉后雌蕊SOD活性會顯著上升,親和性授粉較不親和性授粉上升更顯著;在蕓芥(Eruca sativa Mill.)自花授粉研究中,自交親和授粉雌蕊SOD活性變化較自交不親和變化更顯著[10]。本研究結果表明,授粉親和雌蕊SOD活性升高明顯,特別在授粉前期花粉管迅速生長階段,SOD活性急速顯著升高(P<0.05),推測此時雌蕊內部清除活性氧系統迅速加強,外界刺激誘發的活性氧被迅速清除,為花粉管正常生長提供了便利環境。MDA作為膜脂過氧化最終產物,其含量高低可直接反映細胞受傷害程度[23]。張雪梅等[24]在蘋果(Malus pumila Mill.)自花授粉研究中發現,授粉前期自交親和雌蕊MDA含量較自交不親和高,48 h之后隨著自交親和雌蕊MDA含量下降,花粉管持續生長,自交不親和雌蕊MDA含量升高,花粉管在花柱內停止生長;Ren等[25]對芍藥花粉冷凍保存研究發現,冷凍過程中細胞膜脂過氧化加劇促使MDA含量的增加,細胞功能受損,細胞凋亡率升高,花粉萌發率下降。本試驗結果顯示,花粉萌發、花粉管的生長誘導MDA含量快速升高,同時伴隨MDA含量的急速顯著升高(P<0.05),遠緣雜交授粉花粉管扭曲纏繞并伴有大量胼胝質沉積,推測出現此現象的原因可能是雌蕊細胞膜脂過氧化,導致柱頭及花柱細胞功能受阻,引導花粉管有序生長的信號物質減少,致使花粉管無序生長,進而出現扭曲纏繞,無法順利穿過花柱進入子房完成受精。可溶性蛋白和可溶性糖既是滲透調節物質又是柱頭識別物質的主要成分,在植物授粉過程中有著重要作用。此外,蛋白質代謝是連接植物體內三大代謝的樞紐,在植物抗逆生理中具有重要作用[26]。吳能表等[22]在甘藍雜交研究中發現,自花或異花授粉,均能引起甘藍體內生理生化反應,伴隨花柱內蛋白含量的升高,促進甘藍體內代謝改變。本研究結果表明,授粉后自交與遠緣雜交雌蕊可溶性蛋白含量均表現為先升高后下降趨勢,可溶性糖含量在小范圍波動,整體遠緣雜交雌蕊可溶性糖含量較自交雌蕊含量高,原因可能是遠緣雜交花粉對柱頭刺激作用更加明顯,柱頭對花粉識別及排異作用加強,加速了雌蕊代謝作用,排異物質增加,花粉管生長受阻,顯微特征表現為花粉管內胼胝質的沉積及花粉管扭曲纏繞。
GA3、IAA、ABA、JA等是植物花粉萌發及花粉管生長中的重要調控物質[27-29],張鵬等[30]在加楊(Populus×canadensis Moench)自交和與銀腺楊(P.alba×P.glandulosa)雜交授粉時發現,親和性授粉GA3、IAA、ZR含量較不親和授粉含量高,ABA含量較不親和授粉含量低;劉懿瑤[31]研究表明,茉莉酸信號轉導參與了油茶(Camellia oleifera Abel.))的自交授粉過程,進而參與植物防御過程,花柱中花粉管緩慢生長時,子房接收到JA信號,啟動抑制自交授粉親和的分子機制,花粉管生長受阻;低水平的IPA、ZR、GA3含量引起花粉管尖端膨大、扭曲折疊,是同源四倍體黑皮冬瓜(Benincasa hispida (Thunb.)Cogn.)自交低稔的原因之一[32]。本研究結果表明,在前期花粉-柱頭識別階段,自交授粉雌蕊IAA含量顯著高于遠緣雜交授粉雌蕊,隨IAA含量顯著升高(P<0.05),花粉萌發、花粉管生長迅速,表明高水平IAA對親和性授粉過程中花粉萌發有促進作用。授粉前期,自交授粉IPA、GA3含量較高,遠緣雜交授粉花粉開始萌發時IPA、GA3含量升高,推測高水平IPA、GA3有利于花粉管生長,這與賀丹等[13]在芍藥牡丹遠緣雜交研究結果相似。Kim等[33]研究表明,JA與ABA相互作用來調節植物反應和對非生物脅迫的耐受性,同時JA激活ABA合成基因,促進了ABA的生物合成。Rajesh 等[34]研究表明,ABA調控CALLOSE SYNTHASE 1(CALS1)基因表達上調,抑制分解胼胝質的葡聚糖酶,從而誘導細胞內胼胝質堵塞,限制獲得促進生長的信號達到對楊樹芽休眠的效果。本研究結果表明,在授粉過程中,遠緣雜交授粉雌蕊JA含量高于自交授粉雌蕊,遠緣雜交授粉雌蕊ABA含量遠高于自交授粉雌蕊,特別在授粉后6 h,ABA含量急劇顯著升高(P<0.05),此時遠緣雜交雌蕊胼胝質沉積較多,花粉管扭曲嚴重,推測ABA含量的升高是伴隨JA含量升高延遲發生的,二者共同作用,誘導花粉管細胞內胼胝質堵塞,對花粉管伸長生長起抑制作用。此外,低水平(IAA+GA3+ZR)/ABA是自交不親和的特征表現[35],高水平(IAA+GA3)/ABA有利于授粉親和[12],本研究也證實了這一點,親和性授粉(IAA+GA3)/ABA遠高于不親和授粉,這說明在雜交不親和中起決定作用的是生長素和赤霉素與脫落酸的比值,而不是含量的絕對值。
4 結論
受精前障礙是‘鳳丹’與保康牡丹遠緣雜交不親和的主要原因,具體體現在花粉-柱頭的識別作用及花粉管能否正常伸長生長。Fu等[14]通過鹽溶液處理柱頭以破壞柱頭識別作用,有效克服了亞洲百合(Lilium Asiatica Hybrida)與東方百合((Lilium spp.) Oriental hybrids)受精前障礙;Chen等[15]對煙草(Nicotiana tabacum L.)進行延遲授粉,避開柱頭識別功能強效階段,從而克服煙草雜交不親和受精前障礙;郝津藜等[36]通過對柱頭施加外源GA3,大幅提高牡丹遠緣雜交結實率,在一定程度上克服了牡丹遠緣雜交受精前障礙。結合本試驗,推測通過施加外源IAA、GA3、IPA類植物激素及通過對柱頭識別功能的破壞,或可有效克服‘鳳丹’與保康牡丹受精前障礙,有效提高育種效率,但實際效果有待進一步研究論證。
