北京海坨山典型林分土壤固氮菌群落特征研究
2022-08-03 10:57:56聶立水魏一凡張潤哲聶浩亮薄慧娟
林業科學研究 2022年4期
關鍵詞:差異
史 策,聶立水*,魏一凡,祝 琳,楊 昊,張潤哲,聶浩亮,王 江,薄慧娟
(1.北京林業大學林學院,北京 100083;2.河北農林科學院農業資源與環境研究所,河北 石家莊 050051;3.成都市青白江區農業農村局,四川 成都 610300;4.山西農業大學資源環境學院,山西 晉中 030000)
固氮菌是森林生態系統中重要的功能菌群,它通過固氮酶的作用把大氣中分子態氮還原為植物可利用態氮[1-2],為植物生長提供氮素營養。固氮菌在森林生態系統氮循環,尤其是土壤氮循環中具有重要作用,其群落多樣性及結構組成是土壤氮素固定及維持氮循環平衡的重要指標[3-4]。研究森林土壤固氮菌群落多樣性及組成可對提高土壤氮素水平和森林土壤氮循環機理提供新認識,可為森林植被恢復和森林經營提供科學依據。
已有研究表明固氮菌群落與植被類型、土壤特性等因素有互作效應[5-7]。植物群落通過凋落物和根系分泌物等影響固氮菌的種類和豐度,而固氮菌通過改變土壤環境、土壤養分礦化等方式影響植物群落[8]。Templer 研究表明不同樹種土壤氮保留能力顯著差異[9],并且在不同植被類型下土壤固氮菌群落存在顯著差異[10]。土壤中的碳、氮含量及碳氮比顯著影響土壤固氮菌的活性及分布[5,7,11-13]。相較傳統研究方法,高通量測序技術由于其精確性而被廣泛應用于微生物的研究中[14-15],對于高通量測序檢測土壤固氮菌通常選用nifH作為分子標記基因,因其是編碼鐵蛋白組分的基因且高度保守,在固氮微生物中都可以檢測到[16]。
北半球溫帶森林在調節全球碳平衡和減緩全球氣候變化進程中扮演著重要角色[17-19],華北地區是我國暖溫帶落葉闊葉林的重要分布區域,北京松山自然保護區是華北地區內保持完好,植被類型典型且豐富的自然保護區之一[20],油松林(Pinus tabuliformis Carriere.)是保護區最具代表性的植被類型,區內還分布有核桃楸(Juglans mandshurica Maxim.)、山楊(Populus davidiana Dode.)、白樺(Betula platyphylla Suk.)、蒙古櫟(Quercus mongolica Fisch.)等常見暖溫帶落葉闊葉林[21]。目前已有對該地區不同林分類型下土壤細菌和真菌的研究報道[22],然而對不同林分下土壤固氮微生物群落特征及其與土壤理化性質之間相互關系的研究未見報道。
本研究以北京松山國家級自然保護區內4種典型林分(油松林、針闊混交林、蒙古櫟林、核桃楸林)下土壤為研究對象,以固氮酶鐵蛋白nifH 基因為分子靶標[23-24],采用Illumina MiSeq技術分析不同林分類型下土壤固氮菌的群落特征,探討該地區固氮菌群落的多樣性和結構組成及與其土壤特性的關系,為了解土壤氮循環提供基礎數據,同時為發揮暖溫帶森林土壤潛力和森林植被恢復重建等提供科學依據。
1 研究區概況
研究區位于北京市延慶區松山國家級自然保護區(115°43′44″~115°50′22″ E,40°29′9″~40°33′35″ N)。該區域位處燕山山脈海坨山南麓,海拔628~2 198 m,屬暖溫帶大陸性氣候,年平均氣溫8.9 ℃,年平均日照時數2 836 h,年蒸發量1 772 mm,年降水量493 mm[20]。保護區內成土母巖多為花崗巖,主要土壤類型為山地棕壤、山地褐土和山地草甸土。保護區內植被類型豐富,包括草甸、灌木、喬木等,森林覆蓋率高達87.6%。保護區內主要喬木為:白樺林、油松林、山楊林、核桃楸林、針闊混交林等。
2 研究方法
2.1 樣地設置
依據研究區內油松林、針闊混交林、蒙古櫟林和核桃楸林4種典型林分類型分布情況,在每種林型下設面積為20 m×20 m 3塊標準地,共12塊。分別對所選樣地進行相關因子調查,樣地基本概況見表1。樣地中土壤類型均為山地棕壤,石礫較少,多為壤土、碎屑類結構,成土母質均為坡積母質,林齡均為成熟林,林分起源均為天然林。
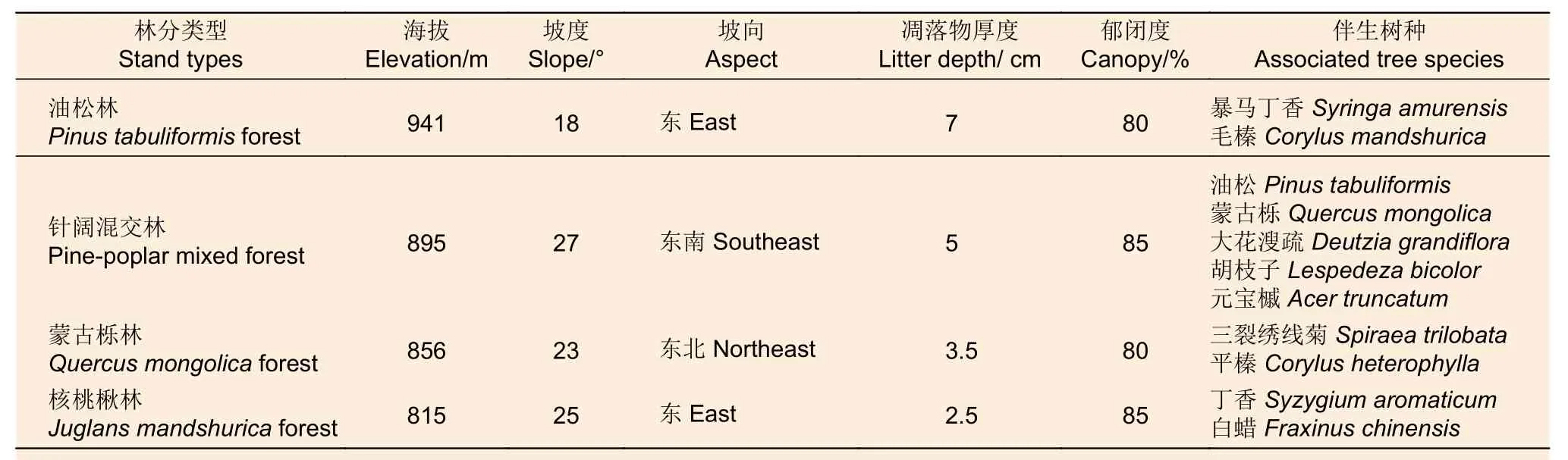
表1 樣地基本概況Table 1 General characteristics of forest stands
2.2 野外調查與采樣
于2019年7月進行野外調查和樣品采集。在不同林型下隨機設置3塊20 m×20 m 的標準樣地,按五點法采集0~20 cm土樣,去除土樣內凋落物和石礫,混合均勻后用四分法取100 g左右的土,裝入無菌自封袋中,共計12份土樣放入便攜式冰箱帶回實驗室。一部分放入?60 ℃冰箱保存,余下風干研磨用作土壤特性分析。
2.3 土壤理化性質測定
土壤特性采用常規分析方法:土壤pH 測定采用酸度計法;土壤全氮測定采用凱氏定氮法;土壤堿解氮測定采用堿解擴散法;土壤有效磷測定采用鉬藍比色法;土壤速效鉀測定采用醋酸銨-火焰光度計法;土壤有機質測定采用重鉻酸鉀-外加熱法[25]。
2.4 DNA提取及PCR擴增
使用Omega Mag-bind soil DNA kit提取土壤微生物總DNA,合格后樣品保存于?80 ℃冰箱用于PCR擴增。擴增引物選擇nifH-F(5′-AAAGGYG GWATCGGYAARTCCACCAC-3′)和nifH-R(5′-TTGTTSGCSGCRTACATSGCCATCAT-3 ′)[26],對fungene-nifH區域進行擴增。每個樣品重復擴增3次,擴增后的序列委托上海派森諾生物有限公司進行測定。PCR 反應體系:5×緩沖液 5 μL;dNTP(2.5 mM)2 μL;正向引物(10 uM)1 μL;反向引物(10 uM)1 μL;DNA 模板 1 μL;超純水 14.75 μL;快速 pfu DNA 聚合酶 0.25 μL。PCR擴增條件:98 ℃ 預變性 5 min;98 ℃ 變性 30 s,63.7 ℃ 退火 30 s,72 ℃ 延伸 45 s,共 35個循環;72 ℃ 延伸 5 min。PCR 反應后,取 5 μL 產物進行1.0% 瓊脂糖凝膠電泳檢測。
2.5 數據處理與統計分析
根據序列質量初步篩選出高通量測序的原始離線數據,并把數據上傳至NCBI平臺,序號為:SUB10898702。QIIME2 DADA2分析進行OTU聚類。采用CD-HIT方法,根據97% 的相似性對OTU進行分類,以獲得每個OTU對應的物種分類信息,利用QIIME分析微生物α和β多樣性,在門和屬分類水平上比較不同林型土壤固氮菌群落結構組成差異。試驗數據處理采用Excel、SPSS24.0和R完成。采用單因素方差分析(one-way ANOVA),Duncan 法比較不同林型的土壤特性、固氮菌多樣性的差異(p<0.05)。采用 Pearson相關系數評價固氮菌多樣性與土壤特性之間的相關關系。采用冗余分析(RDA)檢驗固氮菌結構組成與土壤特性的關系。采用 Spearman 相關性檢驗,分析優勢固氮菌和土壤特性的相關性,運用關聯熱圖展現微生物群落組成與土壤特性的關系。
3 結果
3.1 不同林分類型下土壤特性
不同林分類型的土壤特性見表2。方差分析和多重比較表明,土壤有機質、全氮及速效鉀在4種林型之間均有顯著差異,堿解氮含量表現為:針闊混交林 > 油松林 > 核桃楸林 > 蒙古櫟林,有效磷含量表現為:核桃楸林 > 蒙古櫟林 > 針闊混交林 > 油松林。

表2 不同林分類型下土壤特性Table 2 Soil properties of forest stands
3.2 不同林分類型下土壤固氮菌多樣性
3.2.1 不同林分類型下土壤固氮菌α多樣性 不同林分類型下土壤固氮菌α多樣性指數的統計結果見表3,Chao1指數由高到低表現為:油松林 >針闊混交林 > 核桃楸林 > 蒙古櫟林,核桃楸林和蒙古櫟林之間差異不顯著(p<0.05)。Shannon指數由高到低表現為:油松林 > 針闊混交林 > 核桃楸林 > 蒙古櫟林,針闊混交林和核桃楸林之間差異不顯著(p<0.05)。
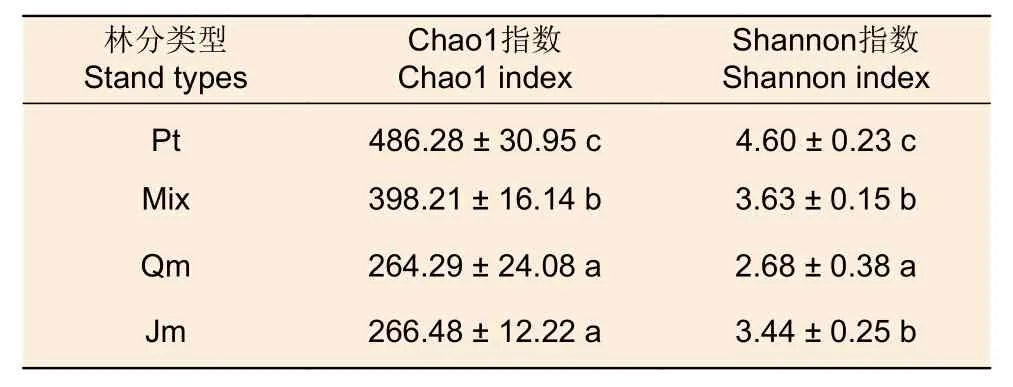
表3 不同林分類型下土壤固氮菌α多樣性指數Table 3 Alpha diversity index of soil nitrogen-fixing bacteria of forest stands
3.2.2 不同林分類型下土壤固氮菌β多樣性 為進一步明晰樣本間在群落物種組成上的差異性,基于Bray-Curtis距離算法并采用 PCoA 分析衡量4種林型土壤固氮菌群落物種組成的相似度。圖1表明,每種林型的3個重復均聚類于同一象限,且每種林分集合間距較大,PCo1 和PCo2分別解釋變量方差的41.7%、21.5%,兩者累計貢獻率達63.2%。表明4種林分類型下土壤固氮菌群落組間差異遠大于組內差異。
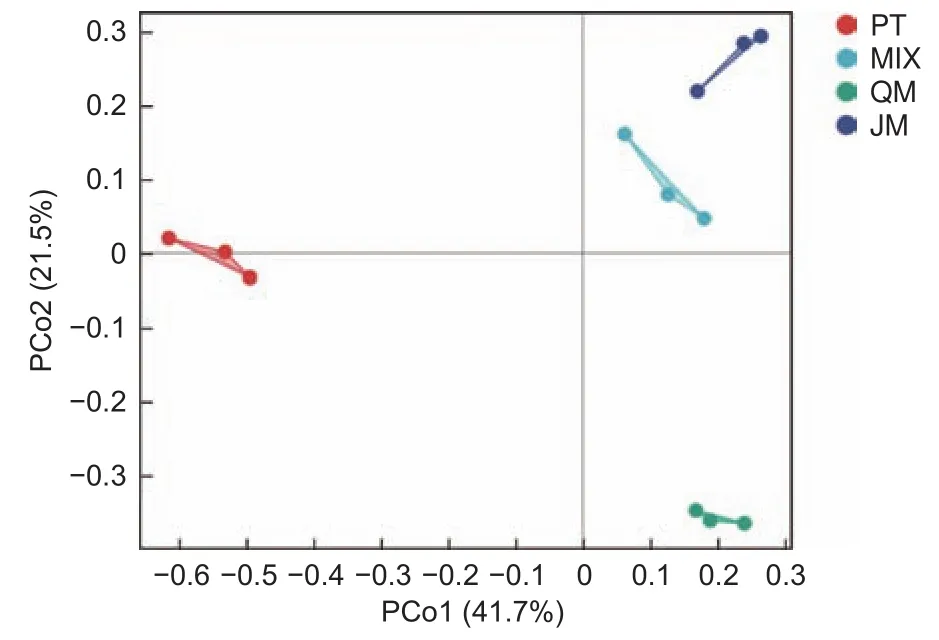
圖1 不同林型下土壤固氮菌群落主坐標分析(PCoA)Fig.1 Principal coordinate analysis (PCoA) analysis of soil nitrogen-fixing bacteria of different forest stands
3.3 不同林分類型下土壤固氮菌群落組成及豐度差異物種分析
為解析不同林型下土壤固氮菌群落結構的變異規律,對聚類后的 OTU 代表序列進行物種注釋。4種林型土壤樣品中的 OTU 隸屬于 4 門 6 綱 13目 16 科 29 屬。將相對豐度低于 1% 以及在該水平上沒有注釋結果的歸于 Others。4種林型土壤固氮菌在門和屬水平上的群落組成見圖2。
在門分類水平上(圖2A),4種林型土壤固氮菌均以變形菌門 (Proteobacteria)為主,其相對豐度范圍是91.87%~73.89%。變形菌門的組成比例在針闊混交林土壤中最高(91.87%),在蒙古櫟林中比例最低(73.89%)。同時在4種林型中還含有酸桿菌門(Acidobacteria),其相對豐度范圍是7.91%~0.40%。另外,厚壁菌門(Firmicutes)僅存在于油松林和針闊混交林土壤中,含量分別為4.97% 和1.76%。雖然群落組成相似,但是各菌門含量差異顯著。
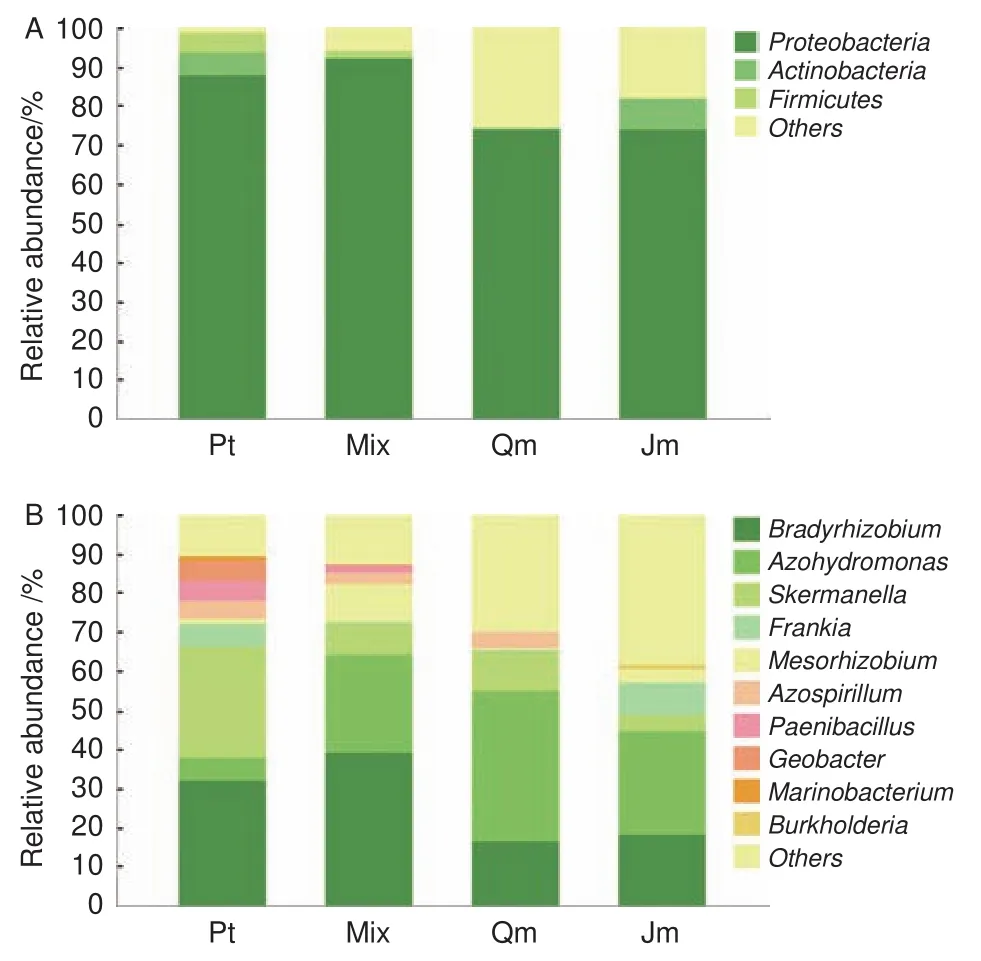
圖2 不同林型土壤固氮菌在門和屬水平上的群落組成Fig.2 The soil nitrogen-fixing bacteria community compositions of different forest stands on phylum(a) and genus(b) level
在屬水平上,取所有樣本中平均相對豐度前10 位的菌群,結果表明4種林型下共有菌屬為慢生根瘤菌屬(Bradyrhizobium),相對豐度為39.24%~16.36%、廣泛固氮氫自養單胞菌屬(Azohydromonas),相對豐度為38.48%~5.98%、斯克爾曼氏菌屬(Skermanella),相對豐度為28.24%~4.41%、弗蘭克氏菌屬(Frankia),相對豐度為7.91%~0.40%、中生根瘤菌屬(Mesorhizobium),相對豐度為9.69%~0.34%、固氮螺菌屬(Azospirillum),相對豐度為4.64%~0.17%。地桿菌屬(Geobacter)僅存在于油松林和針闊混交林下土壤中,海細菌屬(Marinobacterium)僅存在于油松林下土壤中,伯克霍爾德菌屬(Burkholderia)僅存在于核桃楸林下土壤中。
按照97%序列相似性劃分 OTU,根據 Bray-curtis 指數計算距離,得到相似性矩陣,進行群落結構差異分析。由 Venn 圖可見(圖3),4 種林型下共有 OTUs 數目為53個。其中,油松林與針闊混交林共有OTUs數目為124個,針闊混交林與蒙古櫟林、核桃楸林共有OTUs數目78個,同為闊葉林的蒙古櫟林與核桃楸林共有OTUs數目為181個。油松林土壤中所含特有OTUs數量最多為448個,核桃楸林土壤中特有OTUs最少為71個。
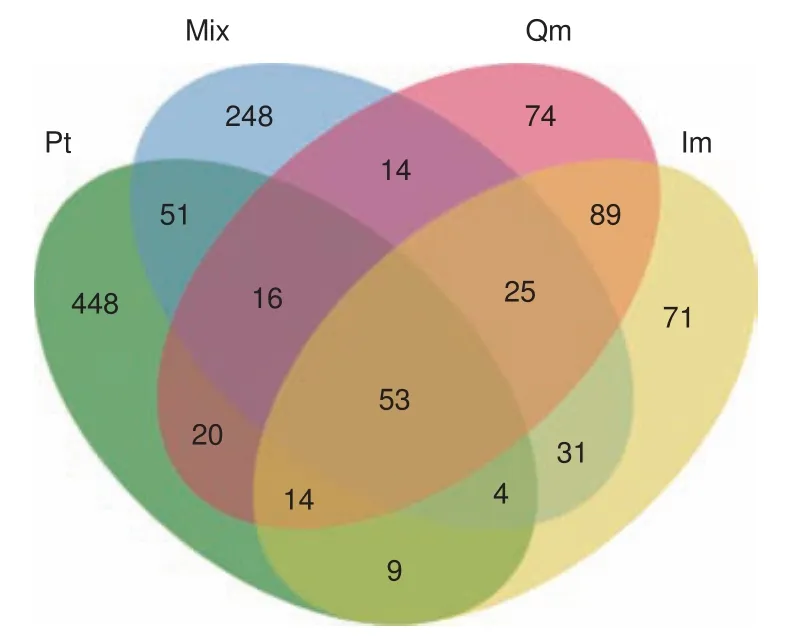
圖3 不同林型下土壤固氮菌的群落差異分析Fig.3 The variance analysis of soil nitrogen-fixing bacteria community of different forest stands
3.4 不同林型下土壤固氮菌群落與土壤理化因子的關聯分析
3.4.1 土壤特性與土壤固氮菌群落多樣性的關系土壤特性與固氮菌的α 和β 多樣性的相關性分析如表4 所示。固氮菌α多樣性與土壤有機質、全氮和速效鉀呈極顯著正相關,堿解氮與Chao1指數呈極顯著正相關,與Shannon指數呈顯著正相關,土壤有效磷與Chao1指數呈顯著負相關。β多樣性數值為相應兩個樣本之間距離(距離矩陣的類型依據算法參數選擇而定)。固氮菌的 β 多樣性與土壤有機質和全氮呈極顯著負相關,這與α多樣性相反,而β 多樣性與土壤速效鉀呈及顯著正相關。
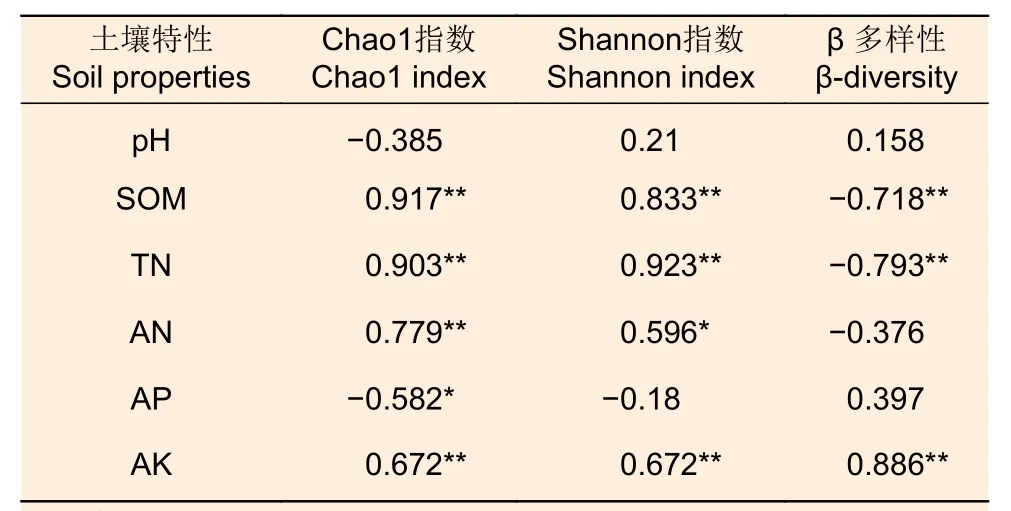
表4 土壤性質和固氮菌群落α和β多樣性的相關性系數Table 4 Correlation coefficients between soil properties and α,β diversity of soil nitrogenfixing bacteria community
3.4.2 土壤特性與土壤固氮菌群落組成的關系采用冗余分析(Redundancy analysis,RDA)探究影響4種不同林型下土壤固氮菌群落組成的環境因子(圖4)。主成分1(48.85%)和主成分2(16.63%)累計解釋土壤固氮菌群落結構變異的65.43%。土壤全氮和有機質是影響土壤固氮菌群落組成的主要土壤特性因子。
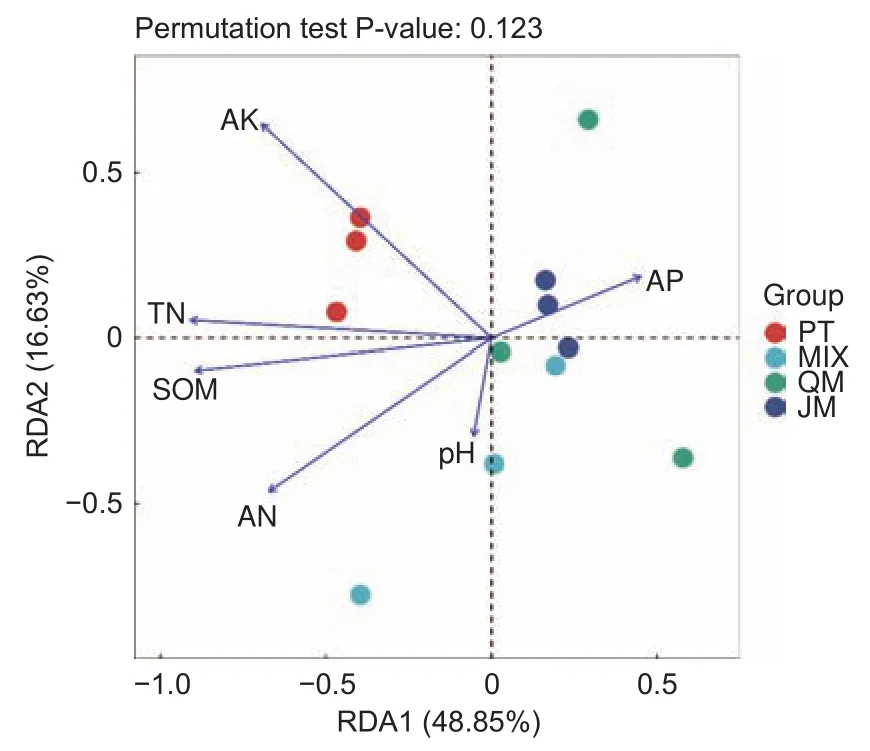
圖4 不同林型下土壤固氮菌群落與土壤特性的冗余分析Fig.4 Redundancy analysis(RDA)between soil properties and soil nitrogen-fixing bacteria community of different forest stands based on Bray-Curtis
采用Spearman相關性系數分析4種林型土壤特性與土壤固氮菌群落組成的相關性,豐度前20菌屬的相關性熱圖(圖5)。其中與土壤速效鉀含量呈顯著相關的菌屬有9個,全氮8個,有機質5個,堿解氮3個,pH 2個。
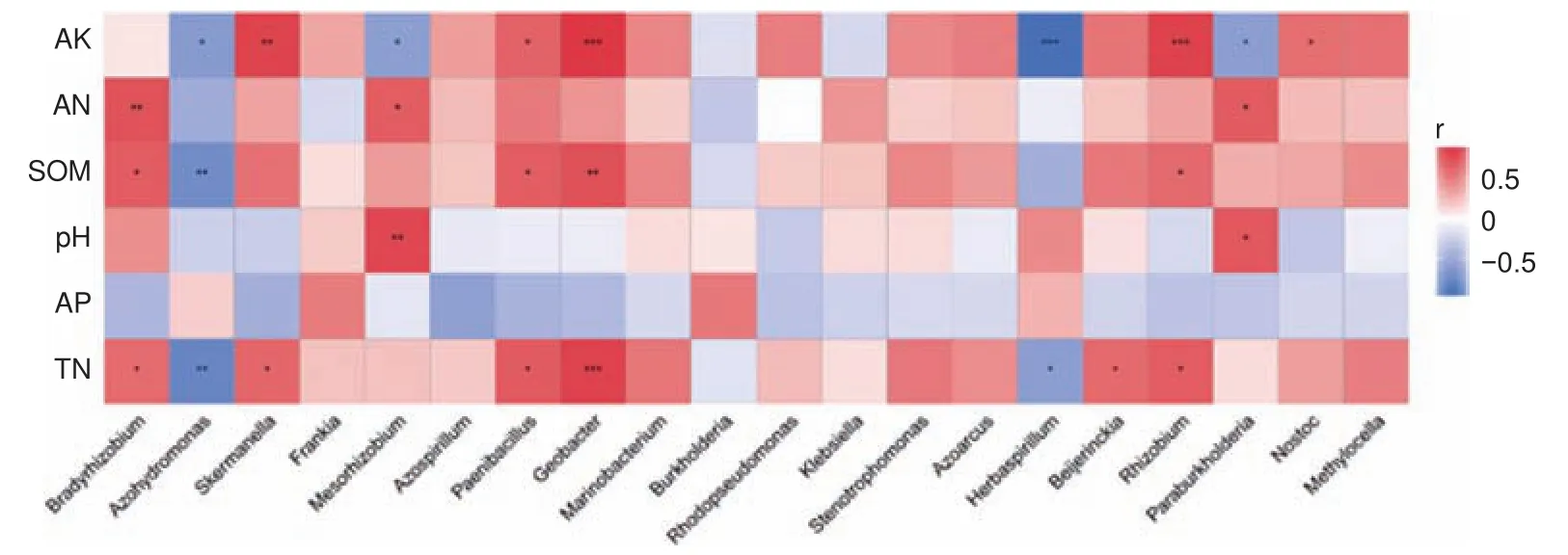
圖5 土壤固氮菌菌屬與環境因子的相關性熱圖Fig.5 Correlation heat maps between the genus of soil nitrogen-fixing bacteria and soil chemical factors.
4 討論
4.1 不同林分類型下土壤固氮菌多樣性及群落結構
不同林分類型的群落結構不同導致凋落物的質量和根系分泌物存在差異,對土壤特性造成影響,從而間接影響固氮菌種間的競爭力,使整個固氮菌的多樣性及群落結構發生改變[27-30]。本研究發現,不同林分類型下土壤固氮菌多樣性存在顯著差異,且油松林下土壤固氮菌多樣性最高,這與牛艷芳對賀蘭山區和大青山的油松林下土壤固氮菌研究一致[31-32]。固氮菌大部分是異養型微生物,充足的凋落物和豐富的有機質能為固氮菌提供利于它們生長繁殖的養分[33-34],本研究中,油松林下凋落物層厚度厚達7.0 cm,這為固氮菌生長繁殖提供了大量的營養物質。同為落葉闊葉林的蒙古櫟林與核桃楸林Shannon指數存在顯著差異,PCoA分析固氮菌β多樣性也明顯區分出不同林型下土壤固氮菌,這說明不同樹種形成的林分是決定土壤固氮菌多樣性的關鍵因素。
本研究結果顯示,不同林分類型下土壤固氮菌群落組成結構存在顯著差異,固氮菌優勢門、屬各占其總種群的40%以上,不同林型下土壤固氮菌群落組成結構差異主要是由其豐度不同和稀少種不同導致的。不同微生物群落對環境變化和適應能力存在差別,森林的生物固氮功能也會對不同環境做出差別響應,因此造成不同區域優勢固氮微生物產生差異。本研究區固氮菌優勢菌門為變形菌門,呂雪麗發現亞熱帶季風氣候森林土壤固氮菌優勢菌門為酸桿菌門,溫帶大陸性山地氣候蒙古櫟林下土壤固氮菌優勢菌門為酸桿菌門和變形菌門。本研究中固氮菌優勢菌屬為慢生根瘤菌屬、廣泛固氮氫自養單胞菌屬和斯克爾曼氏菌屬。慢生根瘤菌可以在寄主植物根部膨大生成根瘤,它們可固定大氣氮為寄主植物可利用氮,而有些固氮菌會在特定條件下以游離狀態固定大氣氮[35-36]。盡管斯克爾曼氏菌的基因組內含nifH 基因,但是目前沒有發現斯克爾曼氏菌屬的物種能夠固定N2[37]。
4.2 土壤特性對土壤固氮菌的影響
在相同的氣候環境條件下,不同樹種的根系分泌物不同及凋落物化學組分和分解速率不同[38-40],導致輸入到土壤中的有機養分不同,造成林下土壤特性差異顯著。相關研究表明,落葉針葉林的凋落物中含有難以分解的木質素、丹寧、蠟等高分子有機物,導致凋落物分解較為緩慢,累積起來凋落物質量較多,厚重的凋落物層覆蓋在土壤表面,從而降低了土壤養分被利用的速率,加速了土壤養分的積累并為固氮菌生長提供養分[41]。本研究中,蒙古櫟林下土壤pH為5.87,與其它闊葉林相比,蒙古櫟凋落物質量較少,且凋落物中N含量較低、C/N較高、木質素含量較高[42]等因素導致蒙古櫟林下土壤pH值偏低。沈秋蘭[43]等對闊葉林改種毛竹后土壤固氮菌多樣性變化的研究表明,不同樹種下土壤固氮菌豐度和群落多樣性差異顯著,導致這種結果這不僅是地上植物類型的改變,土壤性質差異也是原因之一。森林土壤固氮菌群落多樣性和結構的差異主要是不同的土壤條件所造成的,如土壤pH、有機質、全氮和有效磷等[11,44-47]。本研究通過Pearson相關性分析檢驗固氮菌多樣性和土壤環境因子的相關關系,發現土壤特性對不同林型下土壤固氮菌多樣性均有不同程度的影響,其中土壤有機質、全氮、堿解氮、有效磷和速效鉀均與固氮菌多樣性存在相關關系,且大部分土壤特性與固氮菌α多樣性呈正相關。這可能說明在一定范圍內,土壤有效養分含量與固氮菌α多樣性呈正比。以往研究表明土壤pH是影響土壤固氮菌群落結構的重要因子[48],但本研究中,固氮菌多樣性和群落結構與土壤pH并沒有相關性,有可能是因為試驗區土壤pH范圍較窄為5.87~6.93。RDA分析顯示,土壤特性對土壤固氮菌群落結構變化的解釋度為65.43%,土壤全氮和有機質是影響土壤固氮微生物群落組成的主要土壤特性因子。對于固氮菌來說,固氮過程會消耗大量用于生長的碳、氮等能源物質,土壤氮和碳含量不僅是影響土壤固氮菌群落結構和功能的重要因素,而且對調節土壤氮循環起到一定作用[49]。圖5表明,不同種類的固氮菌與土壤性質的相關關系結果差異很大,相關性分析表明慢生根瘤菌屬(Bradyrhizobium)與土壤全氮、堿解氮和有機質呈正相關關系。研究表明慢生根瘤菌是化能自養型細菌[50],且慢生根瘤菌具有很強的固定CO2的能力[51],并在所有土壤中存在。它可以使用還原態的無機物作為能源,例如NH4+、NO2?、S2?、H2S、H2和Fe2+等作為自養生長的能源。
綜上,林分類型、土壤特性、固氮菌三者之間相互影響,存在協同演變機制。然而影響土壤固氮菌的因子還有很多,例如凋落物、季節變化、地理位置、氮沉降等生態環境因子[52],并且微生物群落的分布并非是均勻的[53],不同樣點間局部植被和環境的差異可能引發土壤固氮菌性質差異,因此還需要增加重復樣品采集數量,多尺度地來探討其對土壤固氮菌的影響。在今后研究中,有必要綜合考慮微生物特征及其影響因素,具體分析固氮菌與植物、土壤之間的耦合機制。
5 結論
不同林分類型下土壤固氮菌群落多樣性及結構組成存在顯著差異。α多樣性分析顯示油松林下土壤固氮菌多樣性最高,β多樣性顯示4種林分類型下土壤固氮菌群落組間差異遠大于組內差異。本研究區優勢菌門為變形菌門,優勢菌屬為慢生根瘤菌屬、廣泛固氮氫自養單胞菌屬和斯克爾曼氏菌屬。冗余分析顯示土壤特性能解固氮菌群落結構變異的65.43%,土壤全氮和有機質是影響土壤固氮菌群落組成的主要土壤特性因子。林分類型、土壤特性和土壤固氮菌具有緊密的聯系,研究土壤固氮菌的群落多樣性及結構組成必須考慮林分類型和土壤性質的影響。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
