多菌靈在果園白三葉青貯中的降解及其對微生物群落的影響
2022-08-05 05:10:16戈建珍傅文慧張露藺寶珺趙帥白瑪噶翁寇建村
草業學報 2022年7期
戈建珍,傅文慧,張露,藺寶珺,趙帥,白瑪噶翁,寇建村
(西北農林科技大學草業與草原學院,陜西楊凌 712100)
果園生草是草本植物與果樹協調共生的生態果園地面管理模式,作為現代果園土壤管理先進技術,近年來大量研究表明果園生草在農業生態系統中具有十分重要的作用[1-3]。隨著現代果園生草種植在我國快速發展,果園生草資源利用得到廣泛的關注,研究主要集中在果園生草刈割后返園或堆肥[4-5]。但果園生草不及時刈割,或直接返園后易引發病蟲害[6],果園白三葉(Trifolium repens)刈割后通過埋置或覆蓋的方式返園,其降解速度較慢,需時較長[7]。因此,高效科學地利用這些物質資源具有重要意義,而將果園覆蓋白三葉作為動物飼料來利用是一種重要的途徑。果園生草是重要的飼料來源,由于果樹管理過程中大量使用農藥,使得果園覆蓋植物作為飼料有很大的健康風險。因此,采用科學有效的方法降低果園覆蓋植物白三葉上的農藥殘留,將其作為動物飼料來利用具有重要的意義。
多菌靈廣泛應用于果園生產中的病害防治,能有效防治由真菌(如多子囊菌、半知菌)引起的病害,但也是一類典型的降解緩慢、水溶性低、持久性強的有機污染物[8],因其結構穩定性較高,半衰期較長以及不科學合理地使用,導致土壤環境、動物飼草料和動物源食品中多菌靈農藥殘留含量較高[9]。青貯可以降解多種農藥,青貯微生物在飼料發酵過程中起著決定性的作用,是飼草調制成高品質青貯飼料的關鍵,對青貯過程中原料營養成分的保留[10]、pH 的降低[11]、青貯有害菌的抑制[12]、青貯飼料有氧穩定性[13]以及污染物降解轉化[14]等有重要意義。發酵過程中微生物對化學物品的生物降解是許多有機化合物分解的最重要機制之一[15]。同時,青貯微生物多樣性及其代謝產物也反映了青貯飼料發酵的基本狀況[16]。而多菌靈作為殺菌劑,在青貯發酵過程中能否對青貯微生物產生一定的影響,進而影響青貯發酵的品質,尚未報道。
目前,關于多菌靈農藥與微生物關系的研究主要集中在土壤中多菌靈降解菌的篩選[17]、多菌靈降解的微生物群落[18],以及多菌靈對土壤微生物的毒性[19]等方面。關于青貯發酵對多菌靈農藥的降解及其對青貯發酵過程中微生物群落的影響研究甚少。本研究以黃土高原優勢果園草—白三葉為材料,噴施不同濃度多菌靈為處理,在青貯60 d 后對多菌靈殘留、乳酸、乙酸、丙酸、pH、氨態氮、干物質、粗蛋白和粗纖維等一系列指標進行了檢測,同時,測定了青貯發酵關鍵期(0,3,60 d)的微生物群落。旨在研究青貯發酵條件下,不同濃度多菌靈的降解情況及其對微生物群落的影響,為果園覆蓋植物資源作為青貯飼料利用的安全性以及為種養結合、“林-草-畜”可持續發展循環農業模式提供新思路和理論依據。
1 材料與方法
1.1 試驗區概況
白三葉采自陜西省咸陽市楊凌區現代農業創新園蘋果試驗基地(108°02'E,34°18'N)。試驗區蘋果園年平均氣溫12.9 ℃,無霜期211 d,年平均降水量635.1 mm,年均日照時數2163 h,年總太陽輻射量480.90 kJ·cm-2,日照充足,晝夜溫差大。蘋果園占地5.3 hm2,土壤為黑壚土。試驗區種植果樹品種為“長富2 號”,果樹行間距4 m,株間距2 m,行間種植白三葉。
1.2 青貯前處理
由于缺乏適當的監督和監管,多菌靈農藥的使用量遠超出建議施用的水平[20],所以本試驗用2.0(RU-)、2.5(RU)、3.0 g·L-1(RU+)多菌靈對處于盛花期白三葉進行農藥噴施,噴施等量水作為對照(CON),待各組葉面所噴液體自然揮發后,割取白三葉放到無菌袋中,迅速帶回實驗室,將所取材料切至2~3 cm 長,添加2%蔗糖(20 g)混合均勻后,裝入1 L 青貯罐中,每瓶約800 g,壓實后密封,重復3 次,放置到室溫下青貯,用于檢測農藥殘留及營養指標的測定。對噴施3 g·L-1多菌靈處理進行微生物群落測定,在青貯當天(0 d)取樣標記為A 組;第3天取樣,不噴多菌靈記為B 組,噴施多菌靈記為C 組;第60 天開罐取樣,噴多菌靈記為D 組,不噴施多菌靈記為E組,3 次重復,用16S MetaVx?方法檢測各處理中主要微生物菌群在青貯過程中的群落動態變化。
1.3 青貯后取樣
白三葉青貯60 d 時,從瓶中取出青貯飼料、混勻,稱20 g,加180 mL 蒸餾水,攪拌均勻,用封口膜封口,于4 ℃靜置24 h,四層紗布過濾,再用漏斗過濾,用于pH、有機酸、可溶性糖(water soluble carbohydrates,WSC)及氨態氮的測定;取樣150 g 于65 ℃烘箱烘至恒重后粉碎,用于測定干物質(dry matter,DM),粗蛋白(crude protein,CP),粗纖維(crude fiber,CF),酸性洗滌纖維(acid detergent fiber,ADF),中性洗滌纖維(neutral detergent fiber,NDF),含水量。在青貯第0,3,60 天開罐取樣,用超純水浸泡并不斷攪拌,用細菌濾膜過濾,全程在無菌條件下進行,在-80 ℃冰箱保存用于測定微生物群落。
1.4 測試及分析
通過梅特勒-利多DELTA320 pH 計(上海儀電科學儀器股份有限公司,上海)測定pH,通過高效液相色譜法測定青貯飼料中的有機酸含量,采用苯酚-次氯酸鈉比色法測定氨態氮含量,采用蒽酮-硫酸比色法測定可溶性碳水化合物含量,通過凱氏定氮法測定青貯飼料的粗蛋白含量,酸堿消煮法、中性洗滌劑法、酸性洗滌劑法測定青貯飼料粗纖維、ADF、NDF 含量,烘干法測定干物質含量,用殘余法(SOX406 脂肪測定儀)測定粗脂肪含量,進行青貯后化學成分分析[21],采用液相色譜-串聯質譜法測定多菌靈農藥殘留量[22]。高通量測序文庫構建和Illumina MiSeq 測序是由GENEWIZ 公司(中國蘇州)使用MetaVx?文庫構建試劑盒(GENEWIZ,南平原,新澤西州,美國)來構建測序文庫。采用包含“CCTACGGRRBGCASCAGKVRVGAAT”序列的上游引物和包含“GGACTACNVGGGTWTCTAATCC”序列的下游引物擴增V3 和V4 區。另外,通過PCR 向16S rDNA 的PCR 產物末端加上帶有Index 的接頭,以便進行高通量測序(next generation sequencing,NGS)測序。
1.5 數據處理
用SPSS 22 和Microsoft Office Excel 2010 軟件對數據進行處理,單因素方差分析采用One-way ANOVA,采用Duncan 平均值多重比較分析法對農藥降解率、品質指標等數據進行分析。對原始數據進行去接頭和低質量過濾處理,然后去除嵌合體序列,得到有效序列后進行聚類分析,每一個聚類稱為一個物種操作單元(operational taxonomic units,OTU),對OTU 的代表序列作分類學分析,得到各樣本的物種分布信息。基于OTU 分析結果,對各個樣本進行多種α 多樣性指數分析,得到各樣本物種豐富度和均勻度等信息;基于分類學信息,在各個分類水平上進行群落結構的統計分析;通過計算Unifrac 距離、構建UPGMA 樣本聚類樹、繪制PCoA 圖等,直觀展示不同樣本或分組之間群落結構差異。
2 結果與分析
2.1 不同濃度多菌靈對白三葉青貯品質及其營養成分的影響
本研究中,沒有檢測到丁酸,與對照相比,不同濃度多菌靈處理均顯著提高了白三葉青貯發酵乳酸、乙酸、丙酸的含量(P<0.05),乳酸在多菌靈RU-、RU、RU+濃度下分別增加了47.55%、63.24%、71.08%,乙酸分別增加了24.49%、44.90%、46.94%,丙酸分別增加了66.67%、187.50%、250.00%,并隨著多菌靈噴施濃度的增加而增加。在RU+濃度下乳酸、乙酸、丙酸含量最高,pH 的變化與乳酸、乙酸、丙酸含量變化趨勢相吻合,隨著有機酸的增加pH 值減小(表1)。
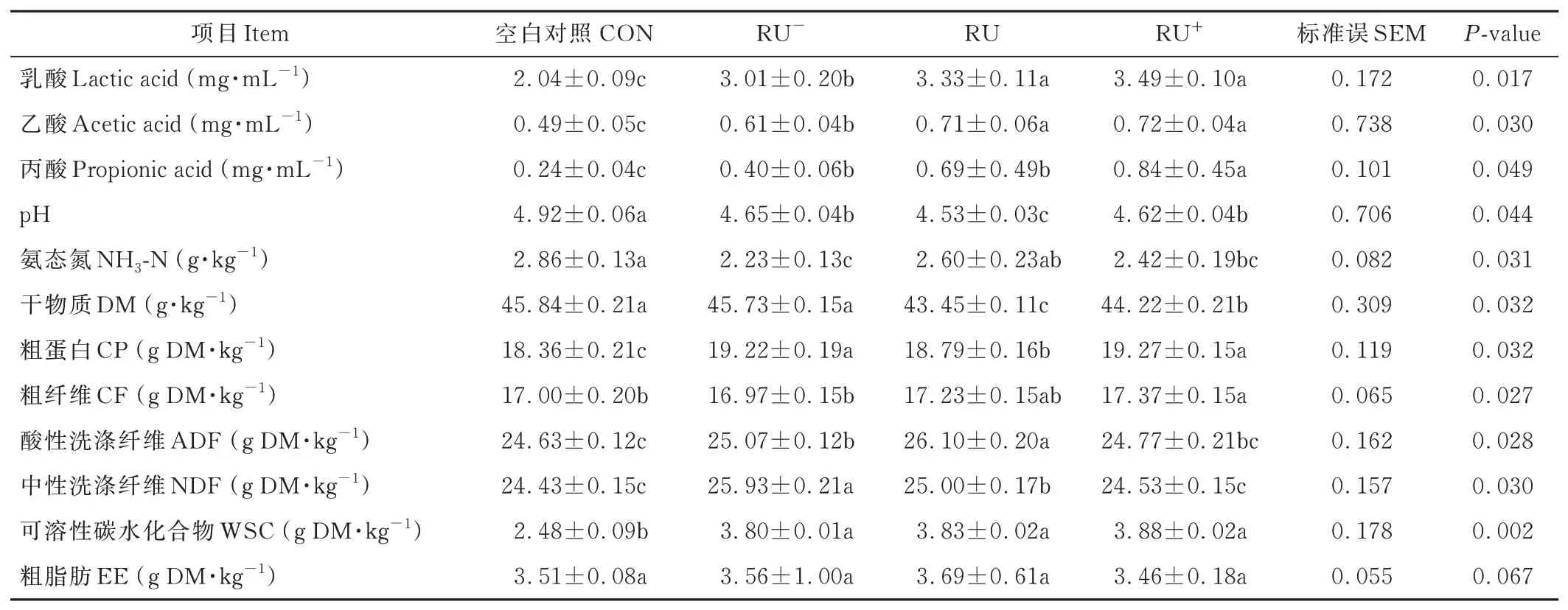
表1 多菌靈對果園白三葉青貯飼料品質及營養價值的影響Table 1 Effect of carbendazim on the quality and nutritional value of white clover silage in orchard
噴施多菌靈的白三葉青貯60 d 后的DM 含量表現為RU、RU+處理顯著低于對照,RU-處理低于對照,但差異不顯著(P>0.05)。不同濃度處理的CP 含量均顯著高于對照(P<0.05),表現為RU+>RU->RU>對照。氨態氮含量與CP 含量變化趨勢相反,RU-、RU+處理下氨態氮含量均顯著低于對照(P<0.05)。CF 表現為RU+處理顯著高于對照。在不同濃度多菌靈處理下,ADF、NDF 含量均高于對照;ADF 含量隨著農藥濃度的增加先增后減,在RU 處理下含量最高;NDF 含量隨著農藥濃度的增加逐漸減小,噴施多菌靈對白三葉青貯粗脂肪無顯著影響。
2.2 白三葉青貯發酵后不同濃度多菌靈的降解
為了確定青貯后不同濃度多菌靈農藥的降解程度,在實驗室進行了模擬筒倉青貯發酵的試驗。將含有初始濃度為37.52,46.88,56.28 mg·kg-1多菌靈的白三葉進行青貯發酵,在發酵后,多菌靈降解情況如圖1 所示。可以看出,RU-濃度多菌靈處理下經過60 d 青貯發酵后多菌靈殘留為15.15 mg·kg-1,RU 處理濃度下多菌靈殘留濃度為16.30 mg·kg-1、RU+處理為16.50 mg·kg-1。3 個處理濃度下(RU-、RU、RU+)多菌靈的降解率分別為59.62%、65.23%、70.68%,并隨著多菌靈濃度的增加降解率逐漸增加。
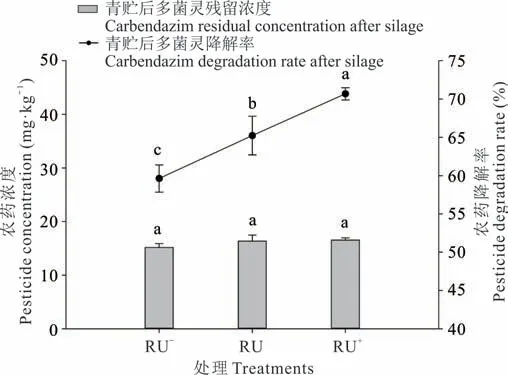
圖1 果園生草白三葉青貯發酵過程對不同濃度多菌靈降解率的影響Fig. 1 The effect of the fermentation process of orchard grass white clover silage on the degradation rate of different concentrations of carbendazim
2.3 果園生草白三葉青貯過程中微生物群落動態變化
2.3.1序列數據預處理及微生物群落結構多樣性分析 分別取青貯第0,3,60 天的青貯樣品DNA,用16S MetaVx?方法檢測各處理中主要微生物菌群在青貯過程中的群落動態變化。為了得到更高質量及更準確的生物信息分析結果,對測序原始數據進行優化處理。本試驗中的5 個樣品共獲得了337551 條有效序列,經過優化選擇了301819 條去除嵌合體后有效序列,平均每個樣品約60364 條序列。在97%的序列相似性標準下,共得到了433 個OTUs,分屬于5 個門,6 個綱,17 個目(表2),A、B、C、D 和E 組分別有2、0、2、1 和1 個獨特的OTU(圖2)。
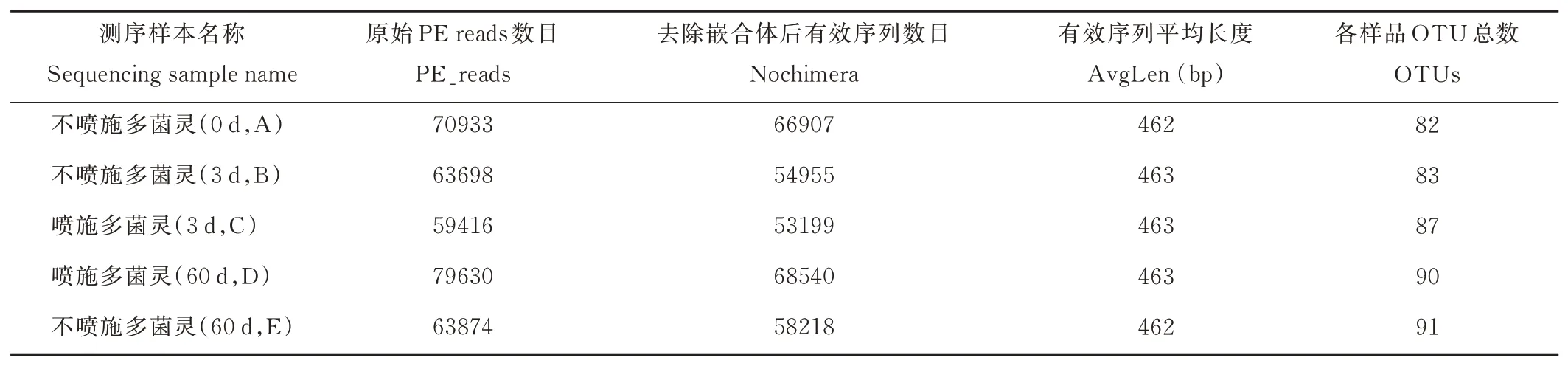
表2 優化后數據質量統計Table 2 Data quality statistics after optimization
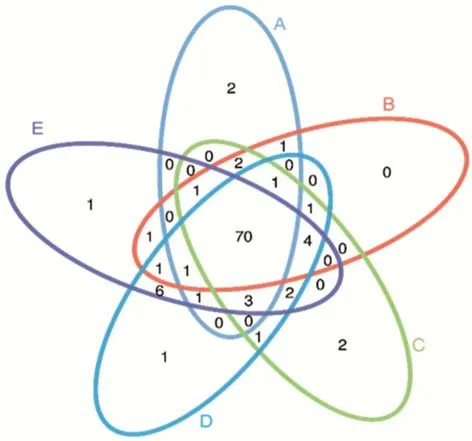
圖2 OTU 韋恩圖Fig.2 OTU Venn diagram
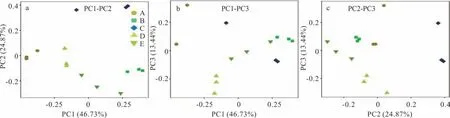
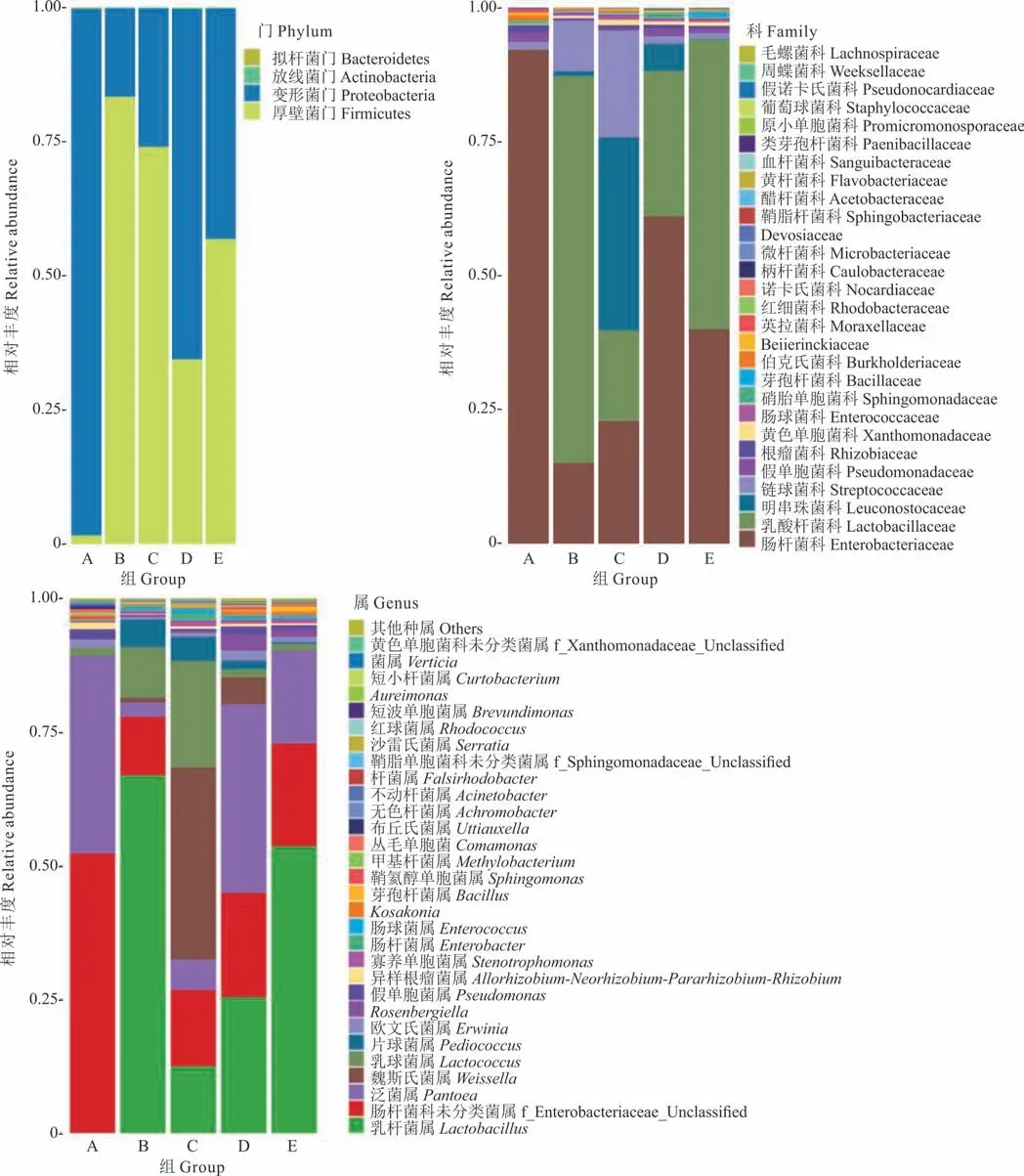
2.3.2微生物群落α 多樣性分析 在本研究中,用Chao 1 指數顯示菌群豐富度(community richness)(圖3a),用Shannon 指數顯示菌群多樣性(community di‐versity)(圖3b)。結果表明,青貯3,60 d 時菌群豐度、多樣性均高于0 d,青貯3 d 時噴施多菌靈處理組(C)菌群豐度、多樣性均高于不噴多菌靈處理組(B),青貯60 d 時噴施多菌靈處理組(D)菌群豐度與不噴多菌靈處理組(E)無差異,噴施多菌靈處理組菌群多樣性顯著高于不噴施多菌靈處理組。添加多菌靈白三葉青貯60 d 與3 d 相比,菌群豐度表現為C>D、多樣性表現為C 圖3 DNA 序列數據和微生物多樣性指數分析Fig.3 DNA sequence data and microbial diversity index analysis 2.3.3噴施多菌靈的白三葉青貯發酵過程中菌群結構分析 圖4 中PC1、PC2 和PC3 軸分別可以解釋46.73%、24.87% 和13.44% 的微生物群落結構差異,3 個主成分累計解釋了樣品中細菌群落組成變化的85.04%。5 個不同處理青貯樣品點分布較為分散,說明各處理間微生物群落構成存在差異。青貯3 d時,噴施多菌靈處理組(C)與不噴施多菌靈處理組(B)樣本點距離較遠,說明微生物群落結構差異較大。隨著時間的推移,在青貯60 d 時,噴施多菌靈處理組(D)與不噴施多菌靈處理組(E)樣本點距離出現顯著聚合現象,說明微生物群落結構差異減小。 圖4 基于Unifrac 距離的PCoA 分析Fig.4 PCoA analysis based on Unifrac distance 在不同的分類水平上分析了多菌靈對白三葉青貯中細菌的群落組成和結構的影響(圖5)。青貯細菌菌群門水平分布包括4 個門(phylum),主要由擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)組成。5 個處理組中變形菌門、厚壁菌門占比較多,青貯3 d 時,B與C 組相比,多菌靈的噴施減少了厚壁菌門的相對豐度,增加了變形菌門的相對豐度。青貯60 d 時,D 與E組相比,菌群豐度變化與青貯3 d 時變化趨勢一致。青貯60 d 與3 d 相比,噴施多菌靈處理主要菌群表現為變形菌門豐度增加,厚壁菌門豐度減少。 圖5 細菌群落分布Fig.5 Bacterial community distribution 青貯細菌菌群科水平分布包括28 個科(family),包括乳酸桿菌科(Lactobacillaceae)、明串珠菌科(Leuconostocaceae)、鏈球菌科(Streptococcaceae)、腸桿菌科(Enterobacteriaceae)、假單胞菌科(Pseudomonadaceae)和芽孢桿菌科(Bacillaceae)等主要菌群。青貯3 d時,B 與C 組相比,多菌靈的噴施減少了乳酸桿菌科、鏈球菌科的相對豐度,增加了明串珠菌科、腸桿菌科、假單胞菌科、芽孢桿菌科的相對豐度。青貯60 d 時,D 與E 組相比菌群豐度變化與青貯3 d 時趨勢一致。青貯60 d 與3 d相比,噴施多菌靈處理主要菌群表現為乳酸桿菌科、腸桿菌科豐度增加,假單胞菌科、芽孢桿菌科、鏈球菌科、明串珠菌科豐度減少。 青貯細菌菌群屬水平分布包括53 個屬(genus),主要包括:泛菌屬(Pantoea)、乳桿菌屬(Lactobacillus)、魏斯氏菌屬(Weissella)、乳球菌屬(Lactococcus)、片球菌屬(Pediococcus)、Rosenbergiella、假單胞菌屬(Pseudomonas)、寡養單胞菌屬(Stenotrophomonas)、腸桿菌屬(Enterobacter)、芽孢桿菌屬(Bacillus)等菌群。青貯3 d 時,B 與C 組相比,多菌靈的噴施減少了乳桿菌屬、片球菌屬的相對豐度,增加了乳球菌屬、魏斯氏菌屬、泛菌屬、Rosenbergiella、假單胞菌屬、寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬的相對豐度。青貯60 d 時,D 與E 組相比,菌群豐度變化與青貯3 d 時趨勢一致。青貯60 d 與3 d 相比,噴施多菌靈處理主要菌群表現為泛菌屬、乳桿菌屬、Rosenbergiella豐度增加,片球菌屬、乳球菌屬、魏斯氏菌屬、寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬豐度減少。 青貯發酵過程中產生有機酸的含量是評價青貯飼料發酵品質的重要因素[23],而青貯過程中pH 值的降低是由一些菌株通過發酵產酸來實現的[24]。研究結果中不同濃度多菌靈處理均顯著提高了白三葉青貯發酵乳酸、乙酸、丙酸的含量,可能是因為多菌靈噴施增加了乳桿菌屬等產酸微生物活性導致乳酸含量增加,例如有研究表明殺菌劑百菌清對蘋果酸-乳酸發酵具有一定的促進作用[14,25]。前人研究表明,青貯發酵開始的前幾天,好氧細菌比較活躍,隨著時間推移,發酵體系中氧氣被消耗完,乳桿菌屬大量繁殖產生乳酸并降低pH,有利于抑制青貯的好氧細菌[24]。本研究中,青貯60 d 與3 d 相比,噴施多菌靈處理主要菌群表現為泛菌屬、乳桿菌屬、Rosenbergiella豐度增加,與前人研究一致。同時殺菌劑可以降低酵母的活性,促進乳酸菌等產酸微生物的代謝,例如He 等[26]研究表明殺菌劑百菌清可以有效抑制酵母發酵產生酒精。 CP 的降解是評估青貯飼料質量的重要標準之一,CP 降解脫氨生成氨態氮[27],芽孢桿菌屬中專性厭氧菌的生長或蛋白水解酶的活性是導致青貯飼料CP 降解的主要因素[28]。本研究中噴施多菌靈農藥處理較對照中芽孢桿菌屬的相對豐度減少,這可能是氨態氮含量低于對照,而CP 含量高于對照的原因。而在不同濃度多菌靈處理下,CF、ADF、NDF 的研究結果可能是因為多菌靈農藥的噴施影響了降解纖維微生物的代謝活性,或者是因為多菌靈的噴施導致乙酸含量增加,間接影響降解纖維微生物的活性。前人研究表明,梭菌屬、纖維桿菌屬、桿菌屬、瘤胃球菌屬是可以有效降解纖維的微生物[29],Fang 等[30]的研究表明殺菌劑戊唑醇可以顯著抑制土壤中梭菌屬微生物活性,Kato 等[31]的研究表明乙酸可以抑制5 種能有效降解纖維的微生物(Clos-tridium straminisolvensCSK1,Clostridiumsp. strain FG4,Pseudoxanthomonassp. strain M1-3,Brevibacillussp. strain M1-5,Bordetellasp.strain M1-6)的降解作用,本研究中多菌靈處理乙酸含量顯著高于對照與之相吻合,但具體原因還需進一步研究。 多菌靈農藥廣泛應用于各種作物,農藥使用不當、過量造成環境污染并對牲畜及人類健康產生危害。世界各國或各組織對多菌靈含量的最低要求不同,例如:歐盟的標準是200 μg·L-1,澳大利亞的標準則是不超過10 μg·L-1,美國的標準是不超過80 μg·L-1,我國標準高于歐美檢測,為5000 μg·L-1[32]。本研究中果園生草白三葉噴施多菌靈后經過青貯發酵,農藥殘留雖顯著減少(P<0.05),但殘留量還是高于安全標準。 本試驗結果可能是因為青貯發酵是多種微生物及化學物質共同作用的復雜反應過程,青貯微生物對農藥的吸附作用和一部分細菌將農藥作為氮源等能量利用,來促進微生物代謝過程,從而降解農藥,高濃度多菌靈較低濃度可以顯著降低pH,為微生物提供更好的降解環境,從而更好地降解多菌靈。例如:蒲丹等[33]從成都彭州蔬菜基地土壤中分離出能以多菌靈作為唯一碳氮源生長的細菌GRPD-1,鑒定其為假單胞菌屬(Pseudomonassp.),該菌株在以多菌靈為唯一碳氮源的基礎上,在培養基中培養6 d 對50 mL 多菌靈的降解率達60%。Xu 等[34]篩選出的紅球菌屬能以多菌靈為唯一碳、氮源生長,在2~3 d 內,可以完全降解100 mg·L-1的多菌靈,降解率高達55.56 mg·L-1·d-1。如Dord 等[35]研究發現與低濃度毒死蜱相比,高濃度毒死蜱可以更好地促進干酪乳桿菌的代謝,從而提高毒死蜱的降解率。李杰等[36]研究表明多菌靈易溶于酸水不溶于堿水,所以在酸性條件下比在堿性條件時降解的更好。 通過測定不同濃度多菌靈農藥對白三葉青貯發酵的影響以及發酵后不同濃度多菌靈降解情況,結果表明多菌靈農藥對白三葉青貯發酵無負面影響,但青貯后多菌靈農藥殘留較高,可能是因為多菌靈對微生物群落的變化及微生物生長活性有一定抑制作用[37]。為了探究青貯后多菌靈殘留量高是否與多菌靈對白三葉青貯過程中微生物群落動態變化影響有關,因此選擇RU+濃度多菌靈進行研究。在群落生態學中,α 多樣性可以反映微生物群落中的物種數量,并且可以通過一系列統計指標來探索環境群落的物種豐度和多樣性的估計[38]。研究結果中菌群豐度、多樣性增加可能是因為青貯前期一部分細菌將多菌靈作為氮源等能量來源來促進微生物繁殖[34],從而導致農藥降解的同時菌群豐度及多樣性得到增加。青貯60 d 噴施多菌靈處理組菌群多樣性顯著下降可能是多菌靈青貯后殘留濃度較高的原因之一。Zhang 等[39]的研究表明噴施毒死蜱顯著降低大腸桿菌和酵母數量,導致農藥殘留較高。 為進一步了解多菌靈對白三葉青貯發酵過程中菌群結構的影響,對不同處理樣品進行主成分分析后,發現隨著時間的推移,多菌靈被降解,殘留濃度減少,導致多菌靈對青貯細菌菌群的結構影響減小。肖麗等[40]的研究表明不同時間內多菌靈處理對土壤細菌群落結構影響有顯著差異,當多菌靈及其中間產物完全降解時,120 d 后多菌靈處理的細菌群落恢復到初始水平。青貯60 d 與3 d 相比,噴施多菌靈處理主要菌群表現為變形菌門豐度增加,厚壁菌門豐度減少,這與前人研究一致,Fang 等[41]的研究發現變形菌門是土壤中降解多菌靈農藥的主要細菌門類。而添加多菌靈使得厚壁菌門微生物豐度下降,這說明變形菌門微生物可能對多菌靈具有耐受性,多菌靈的添加有利于它們生長繁殖,而對厚壁菌門微生物有抑制作用。青貯60 d 與3 d 相比,噴施多菌靈處理主要菌群表現為乳酸桿菌科、腸桿菌科豐度增加,假單胞菌科、芽孢桿菌科、鏈球菌科、明串珠菌科豐度減少。這可能是因為青貯環境有利于明串珠菌科的生長。Gharechahi 等[25]的研究表明明串珠菌科在玉米(Zea mays)青貯中起主導作用。而腸桿菌科、假單胞菌科、芽孢桿菌科作為多菌靈的降解菌,可利用多菌靈作為能量來源來生長繁殖,從而提高豐富度。但隨著農藥的降解,以及青貯環境的改變,假單胞菌科、芽孢桿菌科豐度下降。這可能也是多菌靈殘留較高的原因。青貯60 d 與3 d 相比,噴施多菌靈處理主要菌群表現為泛菌屬、乳桿菌屬、Rosenbergiella豐度增加,片球菌屬、乳球菌屬、魏斯氏菌屬、寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬豐度減少。結果與科水平細菌菌群變化一致。多菌靈的噴施導致細菌群落結構的變化。由于青貯環境的影響以及青貯優勢微生物的競爭抑制作用,導致可以降解多菌靈,如寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬等的細菌豐度的減少,故在生產中可以通過添加一定寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬等多菌靈降解菌,在青貯前期最大限度地降低多菌靈的殘留濃度,從而達到安全標準。 1)青貯后,多菌靈降解率為59.6%~70.7%,隨著農藥濃度的增加降解率顯著增加,噴施多菌靈農藥的白三葉青貯后農藥殘留未達到安全飼用標準;與對照相比,不同濃度多菌靈處理對白三葉青貯發酵過程中乳酸、乙酸、丙酸的產生有顯著促進作用,均顯著降低了青貯環境的pH 值,與低濃度相比,高濃度多菌靈處理中乳酸、乙酸、丙酸含量均顯著增加;多菌靈處理的白三葉青貯后DM、CP、氨態氮含量與農藥濃度有關。2)高通量測序結果顯示多菌靈噴施顯著改變了白三葉青貯菌群群落構成,增加了青貯的菌群豐度、多樣性,多菌靈的噴施減少了乳桿菌屬、片球菌屬的相對豐度,增加了魏斯氏菌屬、泛菌屬、假單胞菌屬、寡養單胞菌屬、腸桿菌屬的相對豐度,不利于青貯發酵;隨著多菌靈的降解菌群豐度、多樣性慢慢恢復,多菌靈的噴施刺激了乳球菌屬、魏斯氏菌屬、泛菌屬、Rosenbergiella、假單胞菌屬、寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬菌群的繁殖。青貯過程中,寡養單胞菌屬、腸桿菌屬、芽孢桿菌屬等具有降解多菌靈功能的菌群隨著青貯時間的推移豐度減少。多菌靈通過對青貯發酵有害菌、多菌靈降解菌群豐度及青貯微生物多樣性產生影響,導致青貯后多菌靈殘留高于歐洲食品安全局規定的作物類動物飼料中農藥最大殘留量,達不到飼用標準。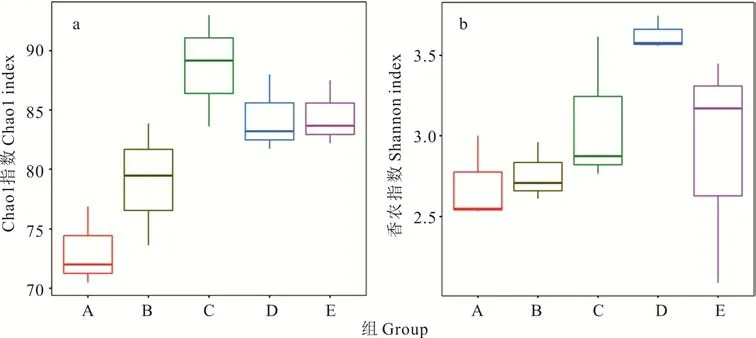
3 討論
3.1 不同濃度多菌靈對白三葉青貯發酵的影響
3.2 白三葉青貯發酵對多菌靈的影響
3.3 多菌靈對果園生草白三葉青貯過程中微生物群落的影響
4 結論
