晾曬、添加物及不同溫度對象草青貯發酵品質和微生物的影響
2022-08-05 05:10:16田靜尹祥樊楊李鑫琴張建國
草業學報 2022年7期
田靜,尹祥,樊楊,李鑫琴,張建國
(華南農業大學南方草業中心,廣東 廣州 510642)
象草(Pennisetum purpureum)是熱帶、亞熱帶地區廣泛栽培的牧草,具有栽培成本低、產量高、營養價值較高等特點[1],但其生長具有季節性,且調制干草較為困難,因此,象草常被調制成青貯飼料以保障反芻動物飼料的常年供應[2-3]。一般象草收獲時水分含量(80%~84%)和緩沖能力較強,青貯飼料的生產受到限制[4]。高水分含量會導致青貯發酵不良和可溶性營養成分損失[5],通過晾曬或添加吸附材料增加原材料的干物質含量,或添加糖源增加水溶性碳水化合物以改善其發酵品質[6]。除此之外,青貯飼料的發酵品質可能會受環境溫度的影響[7]。有研究報道,20~30 ℃下青貯飼料的主要發酵產物是乳酸[8],而高于30 ℃青貯飼料的主要發酵產物是乙酸[8-9]。在青貯初期,植物體不斷呼吸和有氧微生物活動可能使溫度升至40 ℃或更高,導致青貯期間產生大量乙酸、丁酸和酒精[10]。因此,有必要采取適當措施改善溫度對象草青貯發酵的影響,從而為農戶帶來益處。
近幾年,隨著高通量測序的發展,關于不同溫度下青貯飼料微生物群落的研究日益增多[11-12]。Li 等[7]研究表明乳桿菌屬(Lactobacillus)和腸桿菌屬(Enterobacter)可能是引起熱帶青貯飼料產生乙酸的主要原因。而Zhang等[13]在40 ℃條件下青貯苜蓿(Medicago sativa),硫酸鹽還原菌屬(Garciella)是優勢菌屬。不同的溫度條件和原材料本身附著的微生物可能也會影響青貯飼料中的微生物,從而影響發酵過程[14]。然而,有關溫度對象草青貯發酵品質和微生物的影響以及如何改善的研究較少,了解細菌多樣性可能有助于了解獨特的發酵條件并改善象草的青貯飼料質量。
因此,本試驗通過將象草晾曬,或不晾曬直接添加玉米粉或蔗糖,再分別置于20、30 和40 ℃的環境下青貯,探究晾曬和添加物處理下環境溫度對象草青貯發酵品質與微生物的影響,為提高象草發酵質量提供理論支持。
1 材料與方法
1.1 象草種植與試驗設計
試驗所用象草于2020 年8 月在華南農業大學第五教學試驗田收獲。收回實驗室后用鍘草機切割,長度為1~2 cm,將切碎的象草混合均勻。一部分放置室外晾曬(WS),另一部分不晾曬,設置為無任何處理(對照,CK)、添加鮮草10%的玉米粉(CS)或2%的蔗糖(SS),其中,玉米粉干物質(dry matter,DM)含量為93.79%,粗蛋白(crude protein,CP)含量為10.22% DM,中性洗滌纖維(neutral detergent fiber,NDF)和酸性洗滌纖維(acid detergent fiber,ADF)含量分別為8.44% DM 和5.00% DM,可溶性碳水化合物(water-soluble carbohydrate,WSC)含量為20.41% DM。然后將晾曬和添加物處理的象草分別混合均勻后裝至20 cm×30 cm 的聚乙烯塑料袋中,每袋約200 g,每個處理9 個重復,用真空封口機抽真空密封保存,最后分別放置于20、30 和40 ℃的恒溫培養箱中貯藏,即4 個處理×3 個溫度×3 個重復,共36 袋。60 d 后開袋取樣分析,檢測其青貯飼料的發酵品質、微生物數量和細菌多樣性。
1.2 試驗指標測定
1.2.1原材料營養成分及緩沖能的測定 取20 g 樣品,加入80 mL 蒸餾水,使樣品完全浸沒,放置4 ℃冰箱18 h 后,用中性濾紙過濾,得到浸提液,采用Mettler Toledo FE28 pH 計測定pH;采用鹽酸、氫氧化鈉滴定法測定緩沖能[15]。再取約150 g 樣品,稱重后置于70 ℃恒溫烘箱中,烘干至恒重稱重測定干物質含量;粉碎過1.0 cm 篩,用于測定營養成分,采用凱氏定氮法測定粗蛋白[16]含量;粗纖維(crude fiber,CF)、中性洗滌纖維和酸性洗滌纖維含量在范氏法(van Soest)的基礎上使用改進的濾袋分析法[17](ANKOM A200i,北京);可溶性碳水化合物含量采用硫酸-蒽酮法測定[18]。
1.2.2發酵品質和產氣的測定 青貯開袋前,采用排水法測定青貯飼料的產氣量。開袋后,用1.2.1 中的方法測定pH;采用凱氏定氮法測定氨態氮(ammonia nitrogen,NH3-N)含量;將2 mL 浸提液12000 r·min-1在4 ℃離心3 min,用0.45 μm 的微孔濾膜過濾后,用島津LC-20AT 型高效液相色譜儀(日本東京)分析有機酸含量。色譜條件:色譜柱為Eleven Organic Acids on Transgenomic COREGel 87H3;檢測器:RID-10A;流動相:0.1 mmol·L-1的磷酸溶液;流速:1 mL·min-1;柱溫40 ℃;檢測波長210 nm;進樣量20 μL。
1.2.3微生物計數及測序 無菌操作下稱取20 g 樣品,加入90 mL 無菌水,在200 r·min-1搖床中振蕩30 min。吸取1 mL 稀釋至不同濃度后,分別采用MRS 瓊脂培養基(MRS agar medium,MRS)[酪蛋白酶消化物10 g·L-1,牛肉膏粉10 g·L-1,酵母膏粉4 g·L-1,檸檬酸三銨2 g·L-1,乙酸鈉5 g·L-1,硫酸鎂0.2 g·L-1,硫酸錳0.05 g·L-1,磷酸氫二鉀2 g·L-1,葡糖糖20 g·L-1,吐溫-80 1.08 g·L-1,瓊脂15 g·L-1]、營養瓊脂培養基(nutrient agar,NA)[蛋白胨10 g·L-1,牛肉膏粉3 g·L-1,氯化鈉5 g·L-1,瓊脂15 g·L-1]、孟加拉紅培養基(rose bengal agar,RBA)[蛋白胨5 g·L-1,葡糖糖10 g·L-1,磷酸二氫鉀1 g·L-1,硫酸鎂0.5 g·L-1,瓊脂15 g·L-1,孟加拉紅0.033 g·L-1,氯霉素0.1 g·L-1]和亞硫酸鐵瓊脂培養基(iron sulfite agar,ISA)[胰蛋白胨15 g·L-1,大豆蛋白胨5 g·L-1,酵母浸膏5 g·L-1,偏重亞硫酸鈉1 g·L-1,枸櫞酸鐵銨1 g·L-1,瓊脂15 g·L-1]進行乳酸菌(lactic acid bacteria,LAB)、好氧性細菌、酵母菌、霉菌和梭菌劃板培養后計數。LAB 和梭菌用厭氧罐37 ℃培養1~2 d;好氧性細菌、酵母菌和霉菌在有氧條件下30 ℃培養2~3 d。
稱取的分析樣品添加至20 倍體積滅菌的磷酸鹽緩沖液(pH=7.4)中進行DNA 提取,再通過聚合酶鏈式反應(PCR,95 ℃持續2 min,然后在95 ℃持續30 s,55 ℃持續30 s,72 ℃的25 個循環)進行擴增細菌16S 核糖體RNA基 因 的V3~V4 區 域,引 物 為338F(ACTCCTACGGGAGGCAGCAG)和806R(GGACTACHVGGGTW TCTAAT),進行30 s 的擴增[7],最后在72 ℃延伸5 min。最后將PCR 產物在Illumina MiSeq 平臺上測序,使用Trimmomatic v0.33 軟件,對測序得到的Raw Reads 進行過濾;然后用Cutadapt 1.9.1 軟件進行引物序列的識別與去除,得到不包含引物序列的高質量Reads,再拼接序列即Clean Reads,使用UCHIME v4.2 軟件,鑒定并去除嵌合體序列,得到最終有效數據。使用QIIME 2 軟件,對樣品Alpha 多樣性指數進行評估。
1.3 數據統計
微生物數量的計數轉換為log10cfu·g-1FM。采用SPSS 19.0 進行獨立樣本T檢驗,分析晾曬對青貯前原材料的化學特性和微生物數量的影響,用雙因素方差分析和Duncan 法對晾曬、添加物和溫度處理下象草青貯后發酵品質進行多重比較,P<0.05 為差異顯著。
2 結果與分析
2.1 象草青貯前的特性
象草原料特性如表1 所示。象草原材料的DM 含量約17%,晾曬后增加至約25%,晾曬處理顯著(P<0.05)增加了象草的pH、緩沖能和除梭菌外的其他微生物數量,顯著降低了WSC 含量。
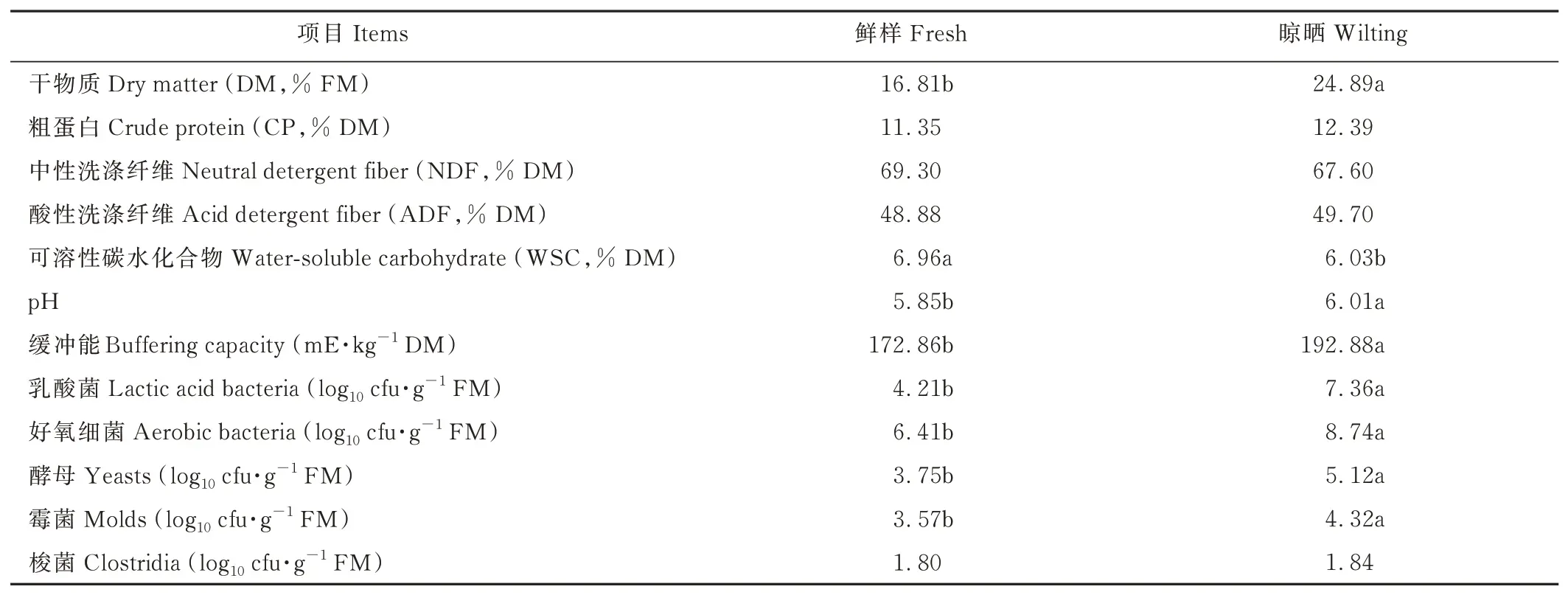
表1 象草晾曬前后的化學特性和微生物數量Table 1 The characteristic and microbial composition of fresh and wilting napier grass
2.2 象草青貯的發酵品質
由表2 可知,青貯60 d 后,對照在30 ℃下的青貯飼料的DM 和乳酸含量顯著高于20 ℃(P<0.05),且40 ℃下的乙酸含量最高,溫度對其pH 無顯著影響(P>0.05)。原材料晾曬后,所有溫度下青貯料的pH 提高,乳酸含量降低,尤在30 ℃,發酵品質最差,表現為pH、乙酸、丁酸和NH3-N 含量最高,乳酸含量最低(P<0.05)。20 和40 ℃條件下,雖晾曬后pH 高于對照,但降低了丁酸和NH3-N 含量。添加10%玉米粉后,與對照相比,每個溫度下的pH 均降低,30 ℃的pH 下降最顯著(P<0.05),但有機酸含量均有所降低,同時,20 和40 ℃下的NH3-N 含量降低,但30 ℃下增加。添加2%蔗糖后的pH 顯著低于對照和除40 ℃的添加玉米粉處理(P<0.05),同時,乳酸含量也較高,乙酸、丁酸和NH3-N 含量降低。方差分析結果表明,溫度除對DM 無顯著影響,NH3-N 顯著影響(P<0.05)外,對其他指標均有極顯著影響(P<0.01),晾曬和添加物處理對所有指標均有極顯著影響(P<0.01),二者的交互作用對pH、乳酸、乙酸和丁酸含量均有極顯著影響(P<0.01)。

表2 不同處理下青貯象草的發酵品質比較Table 2 The fermentation quality comparison of napier grass silage at different treatments
所有在20 ℃下發酵的青貯飼料中,LAB 和好氧細菌數量最多,且隨溫度升高而下降,晾曬處理在每個溫度下均增加了LAB 和好氧細菌數量,而添加玉米粉和蔗糖則降低了其數量,尤其在40 ℃時,效果最顯著(P<0.05)。所有青貯飼料的酵母和霉菌均低于2 log10cfu·g-1FM。方差分析結果表明,溫度、晾曬和添加物處理及兩者交互作用對LAB 數量均有極顯著影響(P<0.001,表3)。
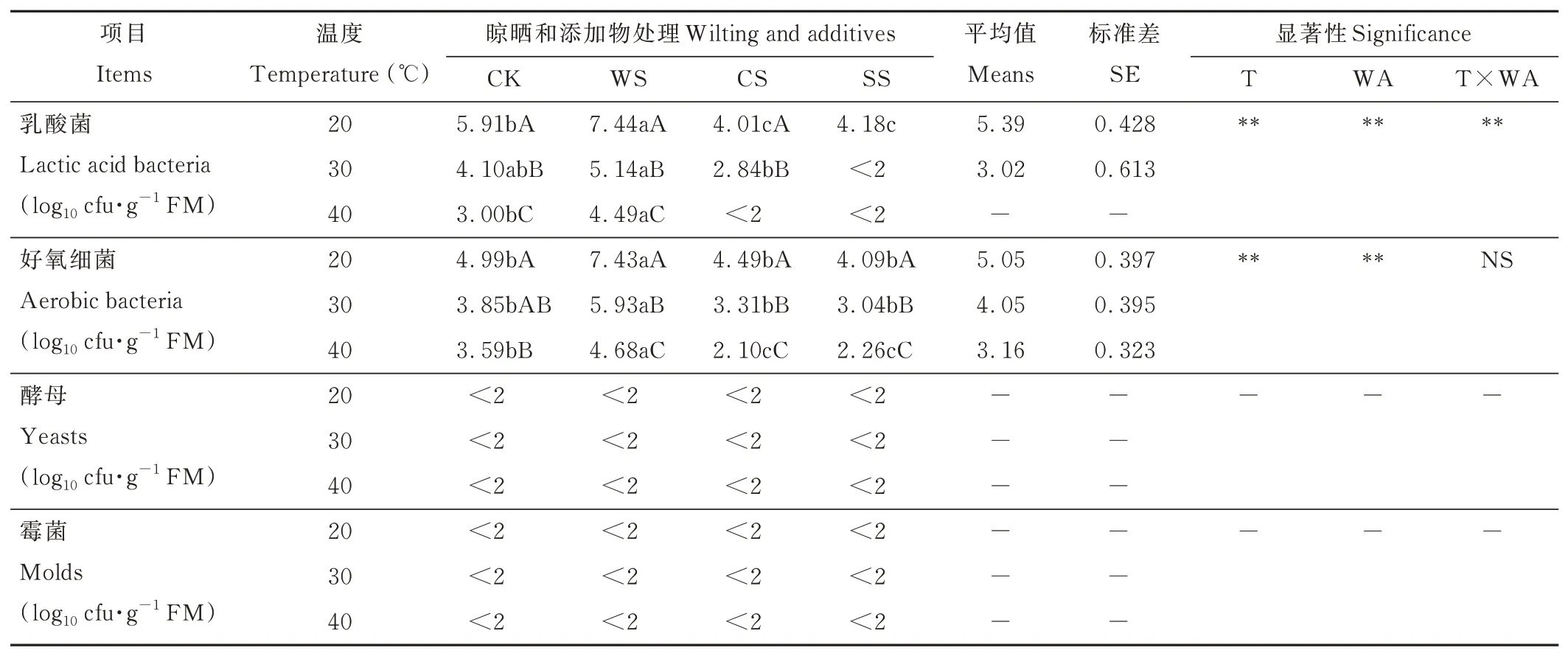
表3 晾曬、添加物和貯藏溫度對青貯象草微生物數量的影響Table 3 Effects of wilting,additives and storage temperature on microbial composition of napier grass silage in 60 days
由圖1 可知,對照、添加玉米粉和蔗糖的青貯飼料在20 ℃下發酵的產氣量均高于30 和40 ℃,但除對照在20 ℃顯著高于40 ℃外,其余差異不顯著,而晾曬處理后,20 和30 ℃條件下的青貯飼料的產氣量相當,均顯著高于40 ℃和其他處理(P<0.05)。40 ℃下各處理間青貯飼料的產氣量差異不顯著(P>0.05)。

圖1 象草青貯的產氣量Fig.1 Gas production of napier grass silage in 60 days
2.3 象草青貯的細菌群落
通過16S rRNA 基因擴增子的高通量測序,共獲得2239912 個細菌序列(每個樣品約158282 個有效序列)。經過生物信息學分析,如表4 所示,這些序列共聚類7582 個可操作分類單元(operational taxonomic units,OTUs)。每個樣本的OTUs 從334 到771 不等,其中,新鮮象草的最多(771)。用來評估細菌多樣性的Chao 1 指 數,Simpson 和Shannon 指 數 顯 示 出 與OTUs 相似的趨勢,均是新鮮象草的細菌豐度高,晾曬后降低。所有處理在40 ℃條件下發酵的青貯飼料的Chao 1、Simpson 和Shannon 指數基本上均高于20 和30 ℃。所有樣品的覆蓋度均為0.99,表明大多數細菌群落被檢測出。

表4 新鮮象草和青貯飼料的細菌群落多樣性統計Table 4 Diversity statistics of bacterial community of fresh napier grass and its silage
圖2顯示了象草青貯前后在科屬水平上的細菌群落的差異。新鮮象草中主要以變形菌門(Proteobacteria)和厚壁菌門(Firmicutes)為主,分別占40% 和29%,晾曬后變形菌門增加至90%,主要是腸桿菌科(Enterobacteriaceae)的腸桿菌屬(Enterobacter)和泛菌屬(Pantoea),分別占27%和47%。與LAB 相關的乳球菌屬(Lactococcus)、乳桿菌屬(Lactobacillus)和魏斯氏菌屬(Weissella)的相對豐度下降。青貯后,飼料中主要由腸桿菌科的腸桿菌屬和鏈球菌科(Streptococcaceae)的乳球菌屬細菌代替。其中,20 ℃下發酵的所有青貯飼料中,腸桿菌屬和乳球菌屬豐度均高于30 和40 ℃,且除晾曬處理30 ℃下腸桿菌屬豐度最低外,對照和添加物處理均是40 ℃下的豐度最低。在晾曬后20 和30 ℃青貯時,腸球菌屬豐度約15%,高于40 ℃。腸桿菌屬豐度在20 ℃下添加玉米粉處理發酵時豐度最高,達到近55%,乳球菌屬豐度在20 ℃下添加蔗糖時最高,達到近38%,而乳桿菌屬豐度在對照30 ℃和添加蔗糖20 ℃條件下較高,約為4.1%,稍高于青貯前的新鮮象草。

圖2 象草青貯前和青貯后的科(a)和屬(b)水平的細菌多樣性Fig.2 Family-(a)and genus-(b)level microbiota analysis of fresh napier grass and its silage
3 討論
3.1 象草青貯前的化學特性
本試驗中新鮮象草的CP 含量較高,為11.35% DM,高于Yin 等[19]和Tao 等[20]報道的10.41% DM 和6.42%DM,低于Gulfam 等[14]報道的12.7% DM。WSC 含量(6.96% DM)高于Yin 等[19]和Gulfam 等[14]的結果,滿足優質青貯飼料制作所需的6%~7% DM[21]。原材料營養成分含量的不同,可能是由于不同地理位置以及生長收獲期等因素影響[22]。晾曬處理顯著提高了象草的DM,降低了WSC 含量,對CP、NDF 和ADF 含量無顯著影響,這一結果與Yin 等[19]報道的一致。新鮮象草中的LAB 數量(4.21 log10cfu·g-1FM)沒達到5 log10cfu·g-1FM 優良青貯飼料的最低要求[3],而晾曬處理顯著增加了LAB 數量(7.36 log10cfu·g-1FM),但好氧細菌、酵母和霉菌數量也顯著增加,好氧細菌數量高達8.74 log10cfu·g-1FM,可能會影響象草的青貯發酵品質。
3.2 象草青貯的發酵品質和微生物多樣性
青貯飼料的pH、有機酸和NH3-N 是衡量青貯飼料發酵品質的重要指標。當青貯飼料pH 低于4.2 時,往往青貯發酵效果較好,而pH 高于4.2 表明發酵品質較差。本試驗中晾曬處理在3 個溫度下的pH 均高于5.0,對照和添加玉米粉處理的pH 均是在30 ℃條件下最低(4.28 和4.14)、20 ℃條件下最高(4.37 和4.24),添加蔗糖處理的青貯飼料pH 值均低于4.2,由此可見添加蔗糖對其青貯品質效果較好。
環境溫度影響青貯料的發酵品質,有研究報道大麥(Hordeum vulgare)秸稈[23]和柱花草(Stylosanthes guianensias)[24]在30 ℃條件下的青貯飼料比20 ℃青貯產生較少的乳酸,較多的乙酸。苜蓿[25]、小麥(Triticum aestivum)和玉米(Zea mays)等[26]在約40 ℃條件下發酵比低溫下青貯產生較少的有機酸,較高的pH 和NH3-N 含量。然而本研究發現,對照和添加玉米粉的象草在不同溫度下青貯對pH 的影響不顯著。添加蔗糖后,20 ℃保存的青貯料pH 顯著低于40 ℃的青貯飼料,且均與30 ℃青貯飼料的pH 無顯著差異,而晾曬后,30 ℃保存的青貯飼料pH 達到5.85,20 和40 ℃條件下pH 為5.5 以下,表明晾曬后的象草在30 ℃青貯時發酵較差,而對照和添加物處理在30 ℃的發酵與20 和40 ℃差異不顯著。原因可能是晾曬時空氣濕度大,原材料的水分含量高,下降緩慢,好氧細菌等有害微生物增多,消耗了較多的WSC,本試驗中晾曬只降低了10 個百分點的水分,未將水分降低至65%以下,因此,晾曬后象草的發酵品質未能改善,反而變差,且30 ℃時發酵最差。因為與20 和40 ℃相比,30 ℃條件下青貯需要更高含量的WSC 來限制丁酸發酵[27],且Yang 等[28]曾報道只有當葡萄糖添加率超過7.0% DM時,LAB 在30 ℃的小麥秸稈青貯飼料中才能占主導地位。與對照相比,添加玉米粉后的青貯飼料的pH 和NH3-N含量降低,乳酸含量升高,但效果不顯著,而添加蔗糖的效果顯著(P<0.05)。原因可能是加糖處理可為LAB 提供充足的底物,相對較低的pH 抑制了有害微生物的生長,使蛋白組分得到了較好的保存,故NH3-N 含量較低。有研究報道高溫(約40 ℃)通常會促進蛋白水解,從而對發酵產生不利影響,加劇NH3-N 和丁酸的產生。而本研究結果表明高溫對NH3-N 沒有顯著影響,只是晾曬處理的青貯飼料的丁酸含量有增加趨勢,原因可能是高溫下乙酸含量的增加抑制了一些蛋白水解微生物[26]。LAB 和好氧細菌數量在添加物處理后的每個溫度下均出現下降,且隨溫度的增加而降低,Li 等[7]也報道了高溫(40 ℃)會顯著抑制LAB 的生長。
Li 等[7]報道乙酸是熱帶地區牧草青貯產生的主要有機酸且隨青貯時間延長乙酸水平上升。而在本研究中除晾曬處理在30 ℃青貯時,乳酸與乙酸含量類似外,象草均以乳酸發酵為主,可能是因為象草表面原附著的菌群和環境條件影響到青貯飼料發酵所涉及的微生物生理和代謝[27]。除晾曬外的其他青貯飼料隨青貯溫度的增加,乙酸含量增加。這與Wang 等[25]的研究結果相一致,可能歸因于高溫抑制了產乳酸細菌(如乳桿菌屬)的代謝[13],而低溫未抑制腸桿菌屬和異型發酵LAB(如乳球菌屬)等的生長。本研究的細菌多樣性也表明腸桿菌屬和乳球菌屬的相對豐度在20 ℃青貯時最高,且晾曬后在20 和30 ℃保存的青貯飼料的腸球菌屬豐度顯著高于40 ℃。Chen等[29]的研究結果也表明青貯過程中產生的CO2與乳球菌屬、明串珠菌屬(Leuconostoc)等呈正相關,與沙雷氏菌屬(Serratia)、鞘氨醇桿菌屬(Sphingobacterium)和片球菌屬(Pediococcus)呈負相關(P<0.05)。因此,本研究中腸桿菌、乳球菌和腸球菌屬可能是導致20 ℃下青貯飼料的產氣量高于30 和40 ℃以及晾曬處理在20 和30 ℃青貯時產氣量均較高的原因。
4 結論
在本研究中,新鮮象草的對照、添加玉米粉或蔗糖處理發酵品質均在30 ℃條件下較好,乳酸含量較高,其中添加蔗糖效果最好;隨青貯溫度升高,乙酸的含量趨于增加。晾曬雖增加了象草的DM 含量,但顯著降低其WSC含量,導致乳酸含量下降、青貯發酵品質變差,特別是30 ℃條件下青貯發酵品質最差。晾曬后在20 和30 ℃時青貯,產氣量顯著高于其他處理。象草青貯前的細菌多樣性較豐富,青貯后主要是腸桿菌屬和乳球菌屬,尤其在20 ℃青貯時的相對豐度較高,晾曬后在20 和30 ℃青貯時,腸球菌屬的相對豐度高于40 ℃。因此,在華南地區潮濕環境下,相比于晾曬,添加玉米粉或蔗糖對其發酵品質效果更好。
