外來入侵植物病毒在我國的發生、危害與管理
2022-08-08 04:38:24陳雅琳潘萌嬌周雪平李方方
植物保護 2022年4期
關鍵詞:癥狀
陳雅琳, 喬 蕊, 潘萌嬌, 周雪平, 李方方
(中國農業科學院植物保護研究所, 植物病蟲害生物學國家重點實驗室, 北京 100193)
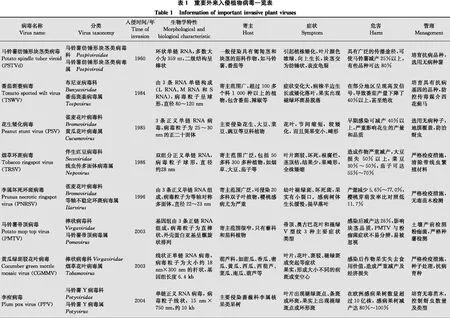
由于全球氣候變暖和國際貿易等行業快速發展的全球經濟一體化,外來生物入侵已成為嚴重的全球性問題,對各國的生態環境、農業發展造成了重大阻礙。目前外來生物入侵也被認作是21世紀五大全球性環境問題之一,開展外來入侵生物的防控已是全球各國政府關注的主要環境問題和工作重心之一[1-2]。在國際環境中,氣候變化和人口壓力正在推動農業實踐和種植制度的迅速改變,這將會導致破壞性的植物病毒病的暴發。由于農業全球化和國際貿易,病毒及其載體傳播到新的地理區域,給糧食生產和自然生態系統帶來意想不到的后果,植物病毒病已成為21世紀全球農業面臨的重大挑戰。根據病毒入侵時間[3],我們總結出我國重要外來入侵植物病毒的種類包括:馬鈴薯紡錘形塊莖類病毒potato spindle tuber viroid (PSTVd)、番茄斑萎病毒tomato spotted wilt virus (TSWV)、花生矮化病毒peanut stunt virus (PSV)、煙草環斑病毒tobacco ringspot virus (TRSV)、李屬壞死環斑病毒prunus necrotic ringspot virus (PNRSV)、馬鈴薯帚頂病毒potato mop-top virus (PMTV)、黃瓜綠斑駁花葉病毒cucumber green mottle mosaic virus (CGMMV)、李痘病毒plum pox virus (PPV)、番茄黃化曲葉病毒tomato yellow leaf curl virus (TYLCV)、木爾坦棉花曲葉病毒cotton leaf curl Multan virus (CLCuMuV)、玉米褪綠斑駁病毒maize chlorotic mottle virus(MCMV)和番茄褐色皺果病毒tomato brown rugose fruit virus (ToBRFV)等。本文將對這12種重要外來入侵植物病毒的相關知識進行梳理,提出適合我國入侵植物病毒的防治建議。
1 重要外來入侵植物病毒
1.1 馬鈴薯紡錘形塊莖類病毒(PSTVd)
馬鈴薯Solanumtuberosum是我國第四大糧食作物,自2015年起, 我國開始逐步推進馬鈴薯主糧化戰略。由于馬鈴薯在生產上主要靠種薯無性繁殖,病毒可通過馬鈴薯塊莖進行世代積累和傳遞,導致馬鈴薯產量和品質大大降低。馬鈴薯紡錘形塊莖類病毒potato spindle tuber viroid (PSTVd)是嚴重影響我國馬鈴薯生產的檢疫性病毒,感病馬鈴薯會出現嚴重的癥狀。PSTVd是侵染植物的最小的植物病原物之一,它是一種環狀、單鏈、無蛋白質包被的RNA。其RNA長度通常為359 nt,由于其堿基高度配對,可折疊成穩定的棒狀二級結構,能夠在寄主體內增殖和移動[4-5]。
1922年在北美發現PSTVd引起的病害,1971年PSTVd被首次分離出來。目前,PSTVd已經在40多個國家被發現,分布于歐洲東部、中亞地區、中美洲、澳大利亞等地區。在我國,PSTVd引起的病害于1960年在黑龍江省首次發現,之后在河北、江蘇、內蒙古、青海等省份相繼發生[6-7]。在田間,除馬鈴薯外,PSTVd還可侵染番茄Lycopersiconesculentum、辣椒Capsicumannuum等多種作物,造成嚴重的經濟損失[8]。
感染PSTVd 后馬鈴薯表現的癥狀與植株生長環境、病原的致病株系以及馬鈴薯的品種有關。一般PSTVd可引起植株矮化、葉片皺縮、塊莖畸形變小、產量降低等,一些易感品種感病后減產達 80%。感病較輕的馬鈴薯不表現癥狀,感病重的馬鈴薯則接近絕產,并且其癥狀可隨感染代數的增加逐年加重。田間生產中,帶毒馬鈴薯種薯傳播PSTVd的效率很高,因此北美等地區主要是通過嚴格控制種薯質量來達到清除田間PSTVd的目的[9]。PSTVd在田間主要以無性繁殖的方式通過農具機械、塊莖切分等接觸的途徑在馬鈴薯間傳播。PSTVd 也可以通過種子傳播,或通過花粉或者卵細胞傳給馬鈴薯實生種子,使其雜交后代攜帶PSTVd,嚴重影響馬鈴薯雜交育種工作。
1.2 番茄斑萎病毒(TSWV)
番茄斑萎病毒tomato spotted wilt virus (TSWV)是蔬菜作物生產的主要威脅之一,造成全世界超過10億美元的損失,宿主范圍超過1 090種植物,超過100個科,包括番茄、豆類、生菜Lactucasativavar.ramosa、花生Arachishypogaea、辣椒、馬鈴薯和煙草Nicotianatabacum等。番茄斑萎病于1915年在澳大利亞首次被發現,然而,直到1930年才確定該病的病原為番茄斑萎病毒。隨后TSWV逐漸蔓延到歐洲、非洲、北美洲和南美洲,亞洲東亞、中東國家[10]。我國1984年首次在廣州發現TSWV,之后TSWV相繼在甘肅、貴州、青海、云南、四川、寧夏、陜西、山東、臺灣等地大面積發生,造成巨大的經濟損失[11 -12]。
TSWV不僅通過西花薊馬Frankliniellaoccidentalis傳播,也在其體內復制,因此,該昆蟲不僅是傳毒介體,還是一種病毒宿主[12]。西花薊馬在其幼蟲期從受侵染的植株獲得病毒,并在幼蟲期結束時將病毒轉移到健康植株上。由于西花薊馬的大面積發生,TSWV已經在部分地區呈現高發信號。土耳其對感染TSWV的番茄的產量開展了研究,發現 TSWV分別導致番茄產量和市場價值下降了42.1%和95.5%[13]。
TSWV的基因組由3條RNA單鏈,L RNA、M RNA和S RNA構成。L RNA長約8.9 kb,是反義RNA鏈,編碼331 kD的RNA依賴的RNA聚合酶(RdRP);M RNA長約4.8 kb,是雙義RNA鏈,M片段的反義RNA鏈編碼2個糖蛋白G1和G2,正義RNA鏈編碼33.6 kD非結構蛋白NSm;S RNA長約2.9 kb,是雙義RNA鏈,S片段的反義RNA鏈編碼28.8 kD的 CP蛋白,正義RNA鏈編碼52.4 kD 非結構蛋白 NSs[14]。TSWV病毒粒體有包膜,大致呈球形,直徑80~120 nm[14]。
不同發育期的番茄受TSWV侵染后的癥狀有所不同。苗期:生長點和新生葉片顏色變為銅色并上卷,后出現黑環狀斑塊,葉背沿葉脈部位變紫,植株矮化,生長速度變緩,出現萎蔫等病癥;坐果期:果實表面出現淡綠環形斑,表面輕微凸起并有細微輪紋;青果期:果實表面出現褐色壞死斑塊、瘤狀突起,果實極易脫落;果實成熟期:輪紋更加明顯,出現許多紅白或紅黃相間的輪紋斑塊,嚴重時整個果實褐色壞死,且果實臍部癥狀與臍腐病十分類似[15]。
番茄種植者為控制這種致命病毒付出過巨大的努力,然而想要徹底控制TSWV難度很大,主要有幾個因素,包括TSWV分離株具有多樣性、TSWV在傳毒媒介中的增殖、傳播效率高、西花薊馬分布廣泛以及對殺蟲劑易產生耐藥性等。因此,培育具有抗病基因的番茄品種是防治這種病毒的當務之急[11, 15-16]。

1.3 花生矮化病毒(PSV)
1966年花生矮化病毒peanut stunt virus (PSV)在美國被首次報道,迄今已報道過該病毒病的國家包括法國、德國、匈牙利、中國、日本、韓國等。我國于1985年才首次報道了PSV,但從20世紀70年代開始由PSV引起的花生病害就一直存在,其中山東、河北、河南等北方主產區損失相對較大。一些相關調查顯示,該病害1976年-1977年在山東省花生生產區的流行尤其嚴重,病株率20%~30%,重者60%以上,嚴重的地塊幾乎絕產[17]。PSV弱毒株系PSV-Mi于1985年在我國被分離和鑒定,但直到2005年其全長序列才被克隆,進化樹和遺傳多樣性分析表明,來源于我國的PSV-Mi屬于一個新的PSV亞組[18-19]。
PSV的基因組為正單鏈 RNA,含 3條正義單鏈RNA(RNA1,RNA2和RNA3),此外還有亞基因組 RNA4。RNA1全長3 347~3 357 nt,RNA2全長2 942~2 966 nt,RNA3全長2 177~2 188 nt。RNA1編碼1a蛋白,RNA2含兩個開放閱讀框(ORF),編碼2a和2b蛋白,部分序列重疊。1a、2a蛋白與病毒復制相關,2b蛋白與病毒的寄主范圍和癥狀表現相關。RNA3上游編碼3a蛋白,為運動蛋白(MP),與病毒在細胞間移動相關;下游ORF為CP基因,CP蛋白由亞基因組RNA4翻譯。病毒粒子為25~30 nm的正二十面體,PSV的外殼蛋白是分子量為26 kD單體多肽[19 -20]。
我國PSV 引起的病害又稱花生普通花葉病,為我國北方花生產區常見的病害,在流行年份其發病率通常大于50%。花生矮化病毒可通過種子傳播,在田間也可由豌豆蚜Acyrthosiphonpisum、桃蚜Myzuspersicae傳播[17, 21]。花生生長早期感染 PSV 后可減產40%以上,感染后的花生表現為莢果變小畸形,花生的產量和品質嚴重下降。在自然條件下,PSV可侵染大豆Glycinemax、豌豆Pisumsativum、菜豆Phaseolusvulgaris等豆科植物,以及芹菜Apiumgraveolens、煙草等,寄主范圍廣泛[21]。
1.4 煙草環斑病毒(TRSV)
煙草環斑病毒tobacco ringspot virus (TRSV)是伴生豇豆病毒科Secoviridae線蟲傳多面體病毒屬Nepovirus的代表種,是我國公布的二類進境檢疫性有害生物。TRSV于1927年首次發現于美國,目前已在50多個國家發生,且廣泛分布于美國中西部和加拿大、歐洲、亞洲、非洲、澳大利亞。我國于1986從1981年采集的樣品中首次鑒定出TRSV[22]。除煙草外,TRSV還能侵染豆科和茄科作物,引起大豆產量損失50%以上, 菜豆減產30%~50%, 茄子減產可達55%~70%[23]。
TRSV的寄主范圍廣泛,能夠侵染馬鈴薯、煙草、大豆等54科300多種的植物,可通過嫁接、帶毒種子以及昆蟲介體傳播,種子和無性繁殖材料是TRSV遠距離傳播的主要方式。其傳播媒介有美洲劍線蟲Xiphinemaamericanum、桃蚜、煙薊馬Thripstabaci等。其病毒粒子球形,基因組為兩條正義單鏈RNA,RNA1編碼RdRP,RNA2編碼MP和CP[24 -25]。
TRSV侵染后的癥狀因寄主不同而有所不同。一般在生長初期,新生幼嫩植株上呈現的癥狀較嚴重,在生長后期不顯著。通常植株感染TRSV后的癥狀表現為葉片上出現環狀或線狀紋、褪綠斑或壞死斑,根腐爛,莖頂枯,結果少,果畸形,節間縮短及矮縮,有些產量降低[26 -29]。
1.5 李屬壞死環斑病毒(PNRSV)
李屬壞死環斑病毒prunus necrotic ringspot virus(PNRSV)是一種在世界范圍內廣泛發生的病毒,屬于雀麥花葉病毒科Bromoviridae等軸不穩定環斑病毒屬Ilarvirus成員。自1941年在桃Prunuspersica和李Prunussalicina樹上首次發現PNRSV以來,該病毒已經在世界多個地區被報道[30]。我國于1996年在陜西省西安市甜櫻桃Prunuspseudocerasus園首次發現并報道了PNRSV[31],目前該病毒在我國東北、華北、華南、西南地區均有過發生和報道。2016年在我國甘肅、河北、河南、遼寧和山東的國產蘋果Maluspumila中發現該病毒,感病蘋果樹葉片均出現花葉和黃脈帶狀癥狀[32]。據統計,在桃樹上PNRSV單獨侵染使植株生長量和產量分別降低12.2%~32.8%和5.6%~77.0%;當其與李矮縮病毒prune dwarf virus (PDV) 混合侵染時,生長量和產量減少49.5%和32.8%,由此失去商品價值;在苗圃內受PNRSV侵染的櫻桃芽萌發率比對照低11.7%[32]。
PNRSV為多分體病毒,基因組由3條正義單鏈RNA組成:RNA1和RNA2分別編碼復制酶蛋白P1、P2,RNA3編碼MP和CP。病毒粒子形態為等軸對稱多面體,直徑22~23 nm[33]。
PNRSV的寄主范圍廣泛,在天然和人工接種的條件下,可侵染的雙子葉植物達21科,其中木本寄主植物中李屬有400余種,包括桃、李和櫻桃,對櫻桃的危害尤其嚴重。其他的木本寄主有西洋梨Pyruscommunisvar.sativa、森林蘋果Malussylvestris、秋海棠Begoniagrandis等[34]。
在桃樹上, 被PNRSV侵染的植株, 有些在春季新生葉片上出現褪綠環形斑、壞死斑,但在夏季時癥狀不易被識別;有些品種在春季不會出現明顯的癥狀,夏季果實縫合線處會出現細小裂口。在櫻桃上,尚未完全展開的葉片上會產生淡黃綠色至綠色環形斑或穿孔,穿孔孔洞的邊緣微突。在蘋果樹上,葉片上形成花葉、條斑、鑲邊、環斑、斑駁等不同類型癥狀,感病樹體的生長速度緩慢并提早落葉[35]。
PNRSV傳播途徑包括人為的運輸、嫁接等,也可通過線蟲、菟絲子等媒介傳播[34]。
1.6 馬鈴薯帚頂病毒(PMTV)
馬鈴薯帚頂病毒potato mop-top virus (PMTV)為馬鈴薯帚頂病毒屬Pomovirus成員。1966年在蘇格蘭和北愛爾蘭地區首次發現通過土壤傳播的PMTV。近幾年該病毒分布的范圍逐漸擴大,主要分布在北歐地區以及北美地區。雖然張仲凱等在2003年通過電鏡觀察發現云南省馬鈴薯病樣中存在疑似PMTV的病毒顆粒[36],一些綜述文章也聲稱我國早就存在PMTV,但并未通過血清學與分子生物學鑒定。直到2013年,在我國廣東惠東縣冬季馬鈴薯中發現了具有壞死環/弧的馬鈴薯塊莖,隨后通過核酸檢測和PMTV抗體酶聯免疫吸附試驗正式確認PMTV在我國發生[37]。最近幾年在國內多個地區,頻繁地出現PMTV發生的報道[38],表明PMTV的傳播速度比較迅速且不容易被控制,需要引起高度重視。
PMTV病毒粒子為直棒狀,一般100~150 nm或(250~300)nm×(18~20)nm。其基因組由3條正鏈的RNA組成。感病植株主要呈現出3種癥狀:奧古巴花葉,植株基部葉片出現不規則黃色斑塊、環紋和線狀紋,植株不矮縮;帚頂,主要表現節間縮短,葉片簇擁變小,葉緣波浪狀卷起,植株矮化束生;褪綠V形紋,常發生在植株上部的葉片,由無癥狀到表面產生環斑、裂縫和畸形,發生程度依品種而異[39 -40]。
PMTV寄主范圍較窄,一般為藜科和茄科植物。PMTV可以通過馬鈴薯粉痂菌Spongosporasubterranean和汁液摩擦兩種方式侵染番茄、煙草和龍葵Solanumnigrum,通過人工接種的方式侵染菠菜Spinaciaoleracea、甜菜Betavulgaris和雜草藜Chenopodiumalbum[40]。PMTV除通過土壤傳播外,還可以通過帶毒的馬鈴薯塊莖傳播且傳播迅速。此外粉痂菌是PMTV的主要寄主,并且PMTV的分布也同粉痂菌的分布大致相同,說明PMTV在沒有馬鈴薯塊莖和土壤時也可以通過其寄主增殖和傳播[40]。
1.7 黃瓜綠斑駁花葉病毒(CGMMV)
黃瓜綠斑駁花葉病毒cucumber green mottle mosaic virus(CGMMV)屬于棒狀病毒科Virgaviridae煙草花葉病毒屬Tobamovirus。該病毒粒子呈桿狀,大小約300 nm×18 nm。CGMMV的基因組為正義單鏈RNA,全長約6.4 kb,編碼4個蛋白質,其中186 kD和129 kD蛋白質為病毒復制和轉錄相關蛋白,29 kD蛋白質為MP,17.3 kD蛋白質為CP[41 -42]。
在世界上很多國家和地區,CGMMV都是葫蘆科作物上的重要檢疫性病毒。該病毒可引起植株葉片黃化以及畸形、生長緩慢、結果推遲、果實黃化或變白并可產生墨綠色水皰狀壞死斑塊,產量損失十分嚴重[43-44]。1935年英國首次報道了CGMMV,隨后西班牙、波蘭、烏克蘭等歐洲國家相繼報道了CGMMV。60年代由于西瓜Citrulluslanatus、黃瓜Cucumissativus等作物的引種,CGMMV傳入了亞洲的印度和日本。隨后在沙特阿拉伯、以色列等國家也發現該病毒,目前CGMMV已在世界上30多個國家和地區發生。2003年,在我國遼寧地區的黃瓜和西瓜中報道了一種具有典型CGMMV所致癥狀的病害,經鑒定其病原為CGMMV[41]。值得重視的是,2005年,我國遼寧蓋州由于種子帶毒而導致的大面積西瓜感染了CGMMV,使其失去了食用價值。在西瓜上,2011年-2013年,在我國山東、江蘇、浙江等省份均有報道CGMMV大面積發生[45]。近年來,CGMMV已成為我國許多省份葫蘆科作物上常見的病毒。根據目前在NCBI上登錄的我國CGMMV全長序列分離物的信息以及CGMMV病癥描述的報道,該病毒已經廣泛分布于我國遼寧、河北、山東、河南、安徽、江蘇、浙江、上海、湖北、湖南、廣西、廣東和海南等地。
CGMMV侵染的大多數植物都屬葫蘆科。這些包括很多瓜類水果和蔬菜,如甜瓜Cucumismelo、西瓜、黃瓜、南瓜Cucurbitamoschata、筍瓜Cucurbitamaxima、冬瓜Benincasahispida、葫蘆Lagenariasiceraria、廣東絲瓜Luffaacutangula、苦瓜Momordicacharantia等。通常寄主被CGMMV侵染后葉片主要的表現為花葉、斑駁、褪綠、突起等癥狀,果實上會形成大小不一的病斑或空心,發病嚴重時可導致無法結果。黃瓜受侵染后最開始在新生葉片上出現黃色小點,后擴展為花葉并伴隨有深綠色瘤狀突起;在侵染初期果實出現淡黃色圓形斑,后發展為綠色突起并產生畸形[42, 46]。CGMMV寄主范圍雖然較為狹窄,但是其分布廣泛,一旦傳入后,擴散的可能性非常大。CGMMV 既可通過整枝、打杈和嫁接等農事操作傳播,也可通過土壤、灌溉水、花粉、種子和菟絲子傳播[46]。可見該病毒可通過多種渠道傳播,預防和治理非常困難。
1.8 李痘病毒(PPV)
李痘病毒plum pox virus (PPV)為馬鈴薯Y病毒科Potyviridae馬鈴薯Y病毒屬Potyvirus成員,引起李痘病,是核果作物中最具毀滅性的病毒病害。該病毒于1932年在歐洲東部保加利亞被發現并迅速傳播至地中海沿岸及歐洲大部分地區,之后又在南美以及北美等地區被發現[47]。2004年在我國湖南發現其危害杏樹Prunusarmeniaca[48],并于2007年被列入《中華人民共和國進境植物檢疫性有害生物名錄》。我國作為世界上最大的核果生產國,其核果病毒的發生頻率和分布仍不清楚。 2008年-2018年對我國核果類果樹進行了系統調查[49],結果發現PPV主要分布在杏和梅Prunusmume上,后者的平均發病率大于30%;在北京、南京、上海、武漢、無錫和運城采集的樣本中, 96% (67/70) 的樣品表現病毒癥狀,77% (85/110) 樣品可以通過免疫層析條帶試驗和RT-PCR檢測到PPV;對4個分離株 (3個來自梅,1個來自杏) 的全基因組序列測定和系統發育分析表明,盡管基因組大小略有差異,但4個分離株均屬于PPV-D分支。 這些發現不僅揭示了PPV在我國的廣泛發生和分布,而且為我國PPV分離株的基因組特征和進化位置提供了詳細的信息。
PPV含有1條正義單鏈RNA,其病毒粒子線狀,長750 nm,寬15 nm,基因組長度約10 kb,包被在由2 000個亞基構成的外殼蛋白中。與其他馬鈴薯Y病毒屬的病毒一致,該病毒的蛋白翻譯策略為:先翻譯一個特殊的長的開放閱讀框,后通過蛋白酶加工產生成熟的病毒蛋白[50]。
PPV主要侵染薔薇科李屬果樹,包括桃、杏、李、日本李P.salicina等及其雜交種。處于不同生長期的寄主均可被PPV侵染,并且在葉片、樹枝、樹皮、花瓣、果實及果核等不同組織都可以表現出病毒癥狀。春季感病的桃樹葉片上產生的癥狀尤其明顯,如產生褪綠斑點、條斑或環斑甚至畸形等癥狀,并且有些桃樹的品種還出現了斷花等癥狀[51]。PPV主要的傳播介體是蚜蟲,有翅蚜在感病后的植株上取食后可遷飛到其他健康植株上繼續為害。因此, 感染PPV的植株數量一般與有翅蚜的種群密度相關[52]。此外,PPV還可以通過嫁接或出芽傳播,感染病毒的樹是無法治愈的,病毒癥狀會隨著時間惡化。現階段抗PPV的種質資源十分稀缺,目前還沒有優良的自然抗性被整合到李屬的優良品種中。
1.9 番茄黃化曲葉病毒(TYLCV)
番茄黃化曲葉病有“番茄癌癥”之稱,其主要病原番茄黃化曲葉病毒tomato yellow leaf curl virus (TYLCV)是一種毀滅性的蔬菜病毒。TYLCV屬雙生病毒科Geminiviridae菜豆金色花葉病毒屬Begomovirus成員。TYLCV是具有孿生顆粒形態的單鏈DNA病毒,病毒粒子為雙聯體結構,大小約為18 nm×30 nm,無包膜[53]。該病毒于1939年在以色列被發現,1959年在約旦大面積暴發,2006年傳入我國[54]。該病毒病害在中東、東亞、東南亞、非洲等多個國家以及地區發生,給番茄造成嚴重的損失。隨全球氣候變化、農耕制度改變、國際貿易活動加強以及介體昆蟲煙粉虱Bemisiatabaci在世界的擴展,TYLCV有蔓延的趨勢,其已經成為影響全世界番茄生產減產的主要因素之一[53 -54]。
TYLCV主要通過煙粉虱傳播,煙粉虱獲毒后,可以終生傳毒但不經卵傳播,一些研究表明TYLCV也可通過嫁接傳播,但不能通過摩擦或種子傳播。據報道,目前該病毒可以侵染茄科、豆科等20多種植物,在這其中,易感病的寄主為:本氏煙Nicotianabenthamiana、曼陀羅Daturastramonium、煙草、心葉煙Nicotianaglutinosa、苦苣菜Sonchusoleraceus、菜豆等。受侵染的植株發病癥狀與病毒株系、寄主的遺傳背景、生長階段以及寄主所處的環境條件有關。發病植株的典型癥狀為頂部葉片變小、皺縮卷曲黃化,節間縮短以及植株明顯矮化,開花延遲、花朵減少,坐果少而小,成熟期果實不能正常由綠轉紅,且成熟狀況不均勻[55]。番茄各生長階段均可感染該病,若在開花前感病,果實產量和商品價值均會大幅下降,嚴重時可造成100%的產量損失[55-56]。值得注意的是,由于該病毒的傳毒介體B型和Q型煙粉虱的大規模暴發,該病毒自從2006年在我國上海發現后,目前已經快速傳播和分布到我國絕大多數省市[54]。
1.10 木爾坦棉花曲葉病毒(CLCuMuV)
在棉花生產上,棉花曲葉病是世界范圍內最具毀滅性的病毒病害, 木爾坦棉花曲葉病毒 cotton leaf curl Multan virus(CLCuMuV)是造成棉花曲葉病的主要病原之一。CLCuMuMV也是雙生病毒科菜豆金色花葉病毒屬成員[57-58]。CLCuMuV的病毒粒子大小、形態以及基因組結構與TYLCV類似。在自然條件下CLCuMuV由煙粉虱傳播,也可以通過嫁接方式傳播,但無法通過摩擦接種傳播。棉花曲葉病最早報道于尼日利亞,隨后在蘇丹(1924年)發現該病害。1967年在巴基斯坦發現棉花曲葉病,1988年后該病在巴基斯坦棉區開始大流行,給棉花生產造成巨大的經濟損失。另外,印度、埃及和南非等國也報道了該病害的嚴重發生與為害。在我國,2006年首次發現由CLCuMuV侵染引起的朱槿曲葉病[57]。隨后,陸續在廣東、廣西等多個省市發現CLCuMuV侵染棉花Gossypiumhirsutum、垂花懸鈴花Malvaviscusarboreus等多種植物。
通過測定和分析34個地理上和/或時間上不同的CLCuMuV分離株和其他6個在我國公開的CLCuMuV基因組的全序列,40株CLCuMuV分離株具有99%的同源性。CLCuMuV和CLCuMuV伴隨的衛星CLCuMuB在我國具有較高的遺傳同源性,表明它們是由單一的毒株進化的[59]。此外,來自不同地區CLCuMuV的衛星群體表現出較高的突變率。雖然在進化上有聯系,但這些種群在不同地區獨立進化[60]。這些研究結果將有助于全面了解世界主要產棉區CLCuMuV群體的遺傳變異現狀和進化模式。
受該病毒侵染后的植株表現的癥狀為全株的葉片上卷、葉脈腫大、產生葉耳、開花少甚至不開花等[58]。在我國各地,該病毒病的傳播媒介煙粉虱已普遍發生,該病害很可能隨煙粉虱快速擴散傳播[61-62]。因此,CLCuMuV對我國棉花等作物的威脅日益增強, 相關部門應采取有效的防控措施進行管控,防止其進一步擴散。
1.11 玉米褪綠斑駁病毒(MCMV)
玉米褪綠斑駁病毒maize chlorotic mottle virus (MCMV) 是我國重要的檢疫性病毒,屬于番茄叢矮病毒科Tombusviridae玉米褪綠斑駁病毒屬Machlomovirus,并且是該屬的唯一成員。MCMV于1971年首次在秘魯發現,目前在巴西、美國等美洲國家,泰國、中國等亞洲國家以及剛果、肯尼亞等非洲國家都有廣泛的分布。2009年我國首次在進境玉米種子中檢測到MCMV[63-64]。MCMV單獨侵染玉米時產生輕微的病毒癥狀,主要可使玉米葉片產生褪綠斑駁的癥狀以及使植株生長緩慢等,可致使玉米的產量比對照降低10%~15%,但當與玉米矮花葉病毒 maize dwarf mosaic virus (MDMV)、小麥線條花葉病毒 wheat streak mosaic virus (WSMV) 或甘蔗花葉病毒sugarcane mosaic virus (SCMV) 等馬鈴薯Y病毒科的病毒復合侵染時可發生協生作用,引起非常嚴重的玉米病毒病害—玉米致死性壞死病 (maize lethal necrosis disease, MLND),可導致植株系統性壞死,使玉米產量損失達到90%[63-65]。
MCMV含有一條長約4.4 kb的基因組RNA,并且能在寄主細胞中合成兩條亞基因組RNA (subgenomic RNA, sg RNA),分別為1.47 kb的sg RNA1、0.34 kb的sg RNA2。病毒粒子的直徑約30 nm,由CP蛋白及其包裹的單鏈正義RNA構成,病毒粒子呈二十面體結構[66]。玉米Zeamays和甘蔗Saccharumofficinarum等是MCMV的天然寄主。MCMV不能侵染雙子葉植物,但能侵染多種單子葉植物。大麥Hordeumvulgare、小麥Triticumaestivum、高粱Sorghumbicolor、黍Panicummiliaceum等均可被MCMV侵染,是MCMV的實驗寄主[66]。
1.12 番茄褐色皺果病毒(ToBRFV)
近年來,一種新的煙草花葉病毒屬Tobamovirus病毒番茄褐色皺果病毒tomato brown rugose fruit virus (ToBRFV) 在歐洲、美洲和亞洲等多個國家和地區的茄科作物產區發生為害,嚴重影響了茄科作物的產品質量以及產量。2014年,在約旦溫室種植的番茄上發現了輕微的花葉以及葉片畸形等病毒病的癥狀,并且其果實上出現了嚴重的著色不均、褐色皺紋等癥狀,使番茄的商品質量以及產量受到影響。在全球范圍,該病毒已快速擴展至歐洲、美洲多地,ToBRFV已經被歐洲及地中海植物保護組織(European and Mediterranean Plant Protection Organization, EPPO)添加到了警戒名單中。我國山東于2019年首次報道該病毒侵染番茄,鑒于其傳播速度快,在我國茄科產區急需加強該病毒的監測[67]。2019年,山東禹城地區3個大棚的番茄,ToBRFV發病率約為50%[67]。山東省是我國最大的蔬菜生產和苗木調度中心,也是我國最重要的設施蔬菜種植區。因此,相關部門應該警惕ToBRFV在茄科作物上的暴發性危害以及在我國的快速擴展[68]。
ToBRFV為正義單鏈RNA病毒,病毒粒子呈桿狀,寬約18 nm,長約300 nm,與典型的煙草花葉病毒屬病毒粒子形態特征類似[69]。
目前報道 ToBRFV的自然寄主主要有番茄和辣椒。感染病毒的番茄葉片的主要癥狀為花葉、深綠色突起、葉片變狹窄、葉脈黃化等,嚴重時葉片壞死,花和果實數量減少,果實上出現黃色或褐色的斑塊,果實變小,出現皺紋,嚴重時會導致果梗的壞死;在辣椒上的癥狀主要表現為植株發育緩慢,葉片上有褶皺和黃色的斑駁狀,嚴重時果實畸形[70]。ToBRFV 通過機械接觸傳播,這意味著該病毒的傳播媒介極其廣泛,只要能與其接觸的都有一定幾率會成為ToBRFV傳播的媒介。對于此類病毒的防控,主要集中在切斷傳播途徑這一方向上[71]。
2 檢疫性植物病毒管理對策
根據我國農業部門的初步調查總結,目前我國外來物種入侵的嚴峻形勢,主要存在3個特點:入侵物種發生種類多、入侵物種分布范圍廣、入侵生物危害嚴重[72]。為減少入侵植物病毒在我國的為害與擴散,提出以下建議:
首先,為了防止破壞性的植物病毒大流行,必須實施檢疫限制,還要在陸地邊境、海港和機場防止病毒傳入[73]。當阻止病毒進入的嘗試失敗時,需要立即應用旨在根除或遏制入侵的檢疫規定,以防止進一步的傳播。應盡量減少任何入侵成功的可能性,這對于防止破壞性植物病毒病害向更多的地方傳播至關重要。值得注意的是,當通過種子傳播的病毒通過國際貿易在不知情的情況下遠距離傳播時,為了避免這種無意的引入,不僅要在收獲前監測種子作物的病毒癥狀,采集其葉片進行病毒檢測,還需要在收獲后對具有代表性的種子樣本進行病毒檢測[73]。此外,在這種檢測不夠嚴格的情況下,作為預防措施,進口國仍需要在每個進口種子發布前檢測具有代表性的樣品。
當植物病毒已經入侵時,需要制定綜合的病毒病管理計劃,以確保有效地控制。設計這些方法需要了解致病病毒的生物學特性以及流行病學。通過結合適當的植物檢疫、栽培、化學農藥和寄主抗性等防控措施來優化病毒病害控制的有效性,進行綜合病害管理,這些措施的作用各不相同,并針對病毒病害周期的不同部分。目前我國也正在制定包括植物檢疫和栽培控制措施在內的更全面的方法,以管理由入侵性植物病毒引起的病害[72]。
為了遏制病毒入侵的源頭,從其他國家引進新品種以及在國家或地區之間進行育種材料交流、保存植物種質時必須生產脫毒植株。體外技術是根除病毒最成功的方法,離體培養技術是生產脫毒植株最成功的策略。到目前為止,已經建立了多種根除植物病毒的方法,包括莖尖培養(也稱分生組織培養)、微嫁接、化療、熱療和莖尖冷凍治療[74]。積累的數據證明,多種方法結合比單獨使用更可有效地去除病毒。例如,莖尖冷凍療法完全不能根除樹莓叢矮病毒raspberry bushy dwarf virus (RBDV)[75]和蘋果莖溝病毒apple stem grooving virus (ASGV)[76],而熱療法與莖尖冷凍療法相結合,分別有33%和100%的植株脫除了RBDV[77]和ASGV[75]。
使用抗病毒作物品種是減少病毒感染造成損失的一種傳統而有效的方法。然而,傳統的抗病毒育種策略,即使加上現代分子技術,如數量性狀位點定位、標記輔助選擇和基于全基因組序列的方法,都是緩慢和費力的,并且需要對多個世代的大型作物群體進行監測[78]。因此,為了優化外來入侵植物病毒控制的管理措施,還應采用新技術,例如利用目前高效的新工具技術NGS(next generation sequencing,二代測序)和生物信息學方法來研究病毒多樣性和宿主在轉錄組上對病毒侵染的反應[79],可以加快入侵植物病毒的研究進程。通過利用基于核酸的生物防治技術,如RNAi(RNA interference, RNA干擾)和CRISPR/Cas9技術[79],可以大大縮短抗病毒作物育種的周期。現階段圍繞RNAi和CRISPR應用的商業預期正在上升,農業生物技術領域的巨頭,如拜耳作物科學,孟山都和先正達,以及許多其他新開發的公司[80]對基于RNA的噴灑技術有著極大的興趣。這一趨勢得到了RNAi相關物質的簡單化生產和成本降低的支持[81]。目前研究人員正在開展大量工作,以擴展和優化基于RNAi的工具(dsRNAs、amiRNAs、tasiRNAs等),用于局部施用以增強植物對病毒的抗性[82]。CRISPR/Cas9系統具有操作簡便、突變效率高和成本低等優勢,能夠通過尋找和編輯外源的核酸或者內源的隱性抗病基因產生抗病毒的植物材料。然而,類似其他技術,CRISPR/Cas9在傳遞病毒的抗性方面也有一些缺陷,比如容易脫靶以及切割后產生新病毒的風險。因此CRISPR基于CRISP/Cas免疫系統設計抗病毒的策略在大規模應用田間之前應該合理、謹慎設計靶點序列。此外,還包括結合精準農業部署遙感等在內的高效新工具技術[73],可分析預測:1)通過衛星監控某區域范圍內病毒病害的發生和流行;2)通過輕型無人飛行器在小范圍內監察外來入侵植物病毒;3)制定防控措施的具體目標。最后,為了有效地防止外來入侵植物病毒在我國的發生和流行,需要組織多學科之間的協作研究。這需要我們國家的植保研究人員,包括植物病毒學者、昆蟲學者和作物育種學者,以及生物信息學專家、統計學專家和社會經濟學專家之間的緊密聯系。
猜你喜歡
初中生學習指導·提升版(2023年8期)2023-09-12 10:26:19
保健醫苑(2022年1期)2022-08-30 08:39:40
中老年保健(2021年12期)2021-08-24 03:30:44
今日農業(2020年17期)2020-10-27 03:10:52
今日農業(2020年16期)2020-09-25 03:05:08
家庭醫學(下半月)(2020年2期)2020-05-11 02:07:18
基層中醫藥(2020年10期)2020-02-13 15:45:52
吉林蔬菜(2017年10期)2017-11-01 07:47:04
獸醫導刊(2016年6期)2016-05-17 03:50:35
中國醫學影像學雜志(2015年9期)2015-12-15 11:03:26
