2種稻蝦共作模式對土壤有機氮礦化作用的影響
2022-08-09 08:32:28喻召雄陶先法侯詒然李子萬
南方農業學報 2022年5期
喻召雄,陶先法,賈 睿,侯詒然,李子萬,董 寅,李 冰*,朱 健,,*
(1上海海洋大學水產與生命學院,上海 200120;2南京農業大學無錫漁業學院,江蘇無錫 214000;3中國水產科學研究院淡水漁業研究中心/農業農村部稻漁綜合種養生態重點實驗室,江蘇無錫 214081)
0 引言
【研究意義】稻蝦共作模式是稻漁綜合種養的典型模式之一,能達到“一田多用、一水多收”的效果,在湖北、江蘇、江西、湖南和安徽等長江中下游地區廣泛分布(陳松文等,2020)。土壤中的氮素大部分以有機氮形式存在,經過礦化作用轉變為無機氮后才可被作物吸收利用。土壤有機氮礦化作用是除氮肥外作物獲取氮素營養的主要途徑,是表征土壤供氮潛力的重要指標(郝曉暉,2008;張笑千等,2010)。土壤有機氮礦化作用受多種因素影響,包括土壤有機氮含量、土壤微生物和底棲生物等(王偉等,2016)。因此,研究不同稻蝦共作模式對土壤有機氮礦化作用的影響,明確蝦類對土壤氮素礦化的作用機制,可為實際生產中氮肥的合理施用提供指導。【前人研究進展】稻蝦共作模式下,蝦類的殘餌、糞便可參與稻田的有機質循環,提高土壤有機質含量,進而提升土壤整體肥力(管勤壯等,2019;楊智景等,2020)。稻田環境下,蝦類是最主要的底棲生物,其擾動作用能增加土壤含水率,提升土壤中有機物的降解程度,進而改變土壤理化性質(龔世園等,2007;張雷等,2011;封功成等,2020)。佀國涵等(2020)研究表明,稻蝦共作可顯著提高微生物量碳和顆粒有機碳含量,對土壤中有機氮等有機質的降解有促進作用。而土壤微生物作為有機質最主要的分解者,其群落結構和活性也主要受土壤環境的影響(佀國涵等,2017;佀國涵等,2020)。王蓉等(2019)研究顯示,稻蝦共作顯著增加了氨氧化古菌與氨氧化細菌的群落豐度,改變了群落結構組成,降低了氨氧化古菌群落多樣性;鄭嬌莉等(2021)在長期稻蝦共作養殖環境中篩選出6株芽孢桿菌有機質降解菌,能快速高效降解餌料培養基中的有機質。稻田耕層(0~20 cm)土壤富含氮素和有機質(吳穎琦等,2019;張錫州等,2000),也是生物活動的主要土層,因此關于有機氮礦化作用的研究多集中于該區域。高亞軍等(2000)研究表明,免耕和常耕兩種耕作方式下,10~15 cm土層的礦質態氮、微生物量氮、礦化勢均顯著低于0~5 cm土層;王根林等(2009)研究指出,可礦化氮隨土層深度的增加而下降,在免耕表土層(0~7.5 cm)含量最高;顧春朝和傅民杰(2016)研究表明,稻田單施化肥能促進耕層表層(0~10 cm)土壤有機氮的氨化過程。【本研究切入點】雖然目前已有較多關于不同耕層有機氮礦化作用的研究報道,但關于耕層下部(10~20 cm)土壤有機氮礦化作用的研究較少,尤其缺乏稻蝦共作模式下土壤有機氮礦化作用的相關研究。【擬解決的關鍵問題】研究2種稻蝦共作模式對稻田耕層下部土壤有機氮礦化作用的影響及相關微生物對礦化作用的響應機制,為稻蝦共生模式中蝦類對土壤氮素肥力的影響機理研究及制定氮肥施用方案提供理論依據。
1 材料與方法
1.1 試驗點概況及試驗材料
試驗點位于江蘇省靖江市(東經120°19′56.8″、北緯32°5′30.9″),年平均氣溫23 ℃,年平均降水量約1055 mm,屬亞熱帶濕潤季風氣候。試驗田為2020年5月末竣工的新建標準化稻田,土壤為油泥土,屬滲育水稻土亞類滲潮粘田土屬,質地均一,砂粘適中,基本理化性質:pH 7.3,全氮0.67g/kg、全磷0.24 g/kg、銨態氮24.62 mg/kg、硝態氮0.91 mg/kg。
試驗用蝦為紅螯螯蝦()和羅氏沼蝦();水稻品種為南粳5055。
1.2 試驗方法
田間試驗開始于2020年7月,采用防水布將試驗田均分為6個小區,單個小區面積約700 m,設稻—紅螯螯蝦(RC)和稻—羅氏沼蝦(RM)2種共作模式,每處理3個平行重復(圖1)。試驗期間,各小區獨立運行,互不影響,水位高度一致。水稻于7月20號移栽,11月12日收獲。移栽采用機插方式進行,株距間隔為10 cm。僅在水稻移栽前1 d施用水稻專用復合肥,施肥量為300 kg/ha,主要成分為N-PO-KO,總養分含量≥40%。蝦苗于8月16日投放,紅螯螯蝦苗種均重約30 g/只,羅氏沼蝦苗種均重約29 g/只;單個小區投放密度為22500尾/ha。蝦苗于11月6日收獲,收獲前進行曬田,緩慢降低各小區水位高度,使紅螯螯蝦離開田面退回到環溝內。2種蝦均為雜食性底棲生物,以水生植物、底棲無脊椎動物、藻類、碎屑喂食。其中,紅螯螯蝦有掘穴行為(黃智偉,2019),羅氏沼蝦無掘穴行為(馮藝,2018)。
試驗于9月1日開始,11月1日結束,為期60 d。養殖過程中每日投餌量為放養總重的2%(每隔20 d通過打樣評估蝦總重),投喂時間為每日下午17:00點,采用無人機對各小區紅螯螯蝦、羅氏沼蝦進行投喂。
1.3 樣品采集
在試驗開始、結束時分別采集初期樣本RC-1、RM-1和末期樣本RC-2、RM-2。單小區選用五點取樣法,用柱狀采泥器采取耕層下部10~20 cm土壤,混合為1個樣品。采集的土樣去除作物根系等雜物,均勻分為3份。1份鮮土用于土壤微生物測定,其余經自然風干后過2 mm篩網,4 ℃保存,用于室內礦化培養試驗和土壤全氮、銨態氮和硝態氮含量測定。
1.4 測定項目及方法
土壤全氮含量采用過硫酸鉀消解—紫外分光光度法測定,土壤銨態氮含量采用氯化鉀浸提—納氏試劑分光光度法測定,土壤硝態氮含量采用氯化鉀浸提—紫外分光光度法測定。
土壤樣品經室內礦化培養后計算土壤有機氮的單周礦化量、累積礦化量、礦化勢、一級反應速率和可礦化氮比例,培養方法采用改進的長期淹水密閉—間歇淋洗培養法(李文軍等,2019)。其中,土壤氮素礦化速率采用一階動力學模型計算(李慧琳等,2008):

式中,為在時間內土壤氮素凈礦化量(mg/kg),累積礦化量為各時期礦化產物NH-N含量之和,表示培養時間(d),為氮礦化勢(mg/kg),k為氮素礦化一級反應速率[mg/(kg·d)]。礦化勢指土壤潛在最大可礦化氮量;對同一土壤而言,其數值固定,不隨土壤培養條件的變化而變化。數值按如下方法確定:另取各采樣時期的土壤樣本,于30 ℃下進行室內礦化培養,培養時間為120 d,將各時期測得NH
-N相加即為。
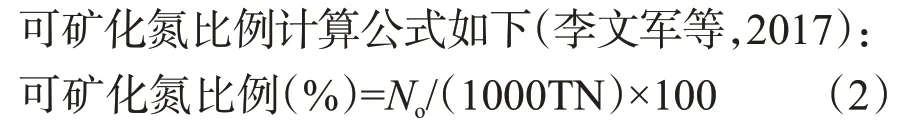
式中,TN為土壤全氮含量(mg/kg)。
土壤微生物群落組成測定:采用Fast DNA SPIN Kit(MP Biomedical,France)試劑盒提取土壤DNA,并用1%瓊脂糖凝膠電泳對其進行檢測;按指定測序區域,合成帶有barcode的特異引物,采用TransGen AP221-02:TransStart Fastpfu DNA Poly‐merase方法對基因組DNA進行PCR擴增,采用2%瓊脂糖凝膠電泳檢測PCR產物,使用AxyPrepDNA凝膠回收試劑盒(AXYGEN公司)切膠回收PCR產物,并用Tris HCl洗脫;將擴增產物委托上海凌恩生物科技有限公司完成Illumina PE250測序流程;將微生物序列信息根據overlap關系進行拼接,同時對序列質量進行質控和過濾。
1.5 數據處理及統計分析
利用Usearch(vsesion 10)將優質序列聚類成操作分類單元(Operational Taxonomic Units,OTU),按照97%相似性對非重復序列(不含單序列)進行OTU聚類,得到OTU的代表序列;采用RDP classifier貝葉斯算法對97%相似水平的OTU代表序列進行分類學分析,使用Mothur 1.30.1在相似水平97%上進行微生物多樣性指數評估;采用上海凌恩生物科技有限公司網上平臺完成微生物主坐標分析(PCoA分析)。
利用SPSS 25.0對測定數據進行統計分析,利用GraphPad Prism 8.0.2繪圖。
2 結果與分析
2.1 2種稻蝦共作模式對耕層10~20 cm土層土壤氮素含量的影響
各時期土壤氮素含量變化如圖2,在試驗初期,2種模式間土壤全氮含量無顯著差異(>0.05,下同),但RC模式的銨態氮和硝態氮含量顯著高于RM模式(<0.05,下同)。試驗末期,RC模式土壤全氮含量顯著低于RM模式,銨態氮含量顯著高于RM模式,硝態氮含量無顯著差異。
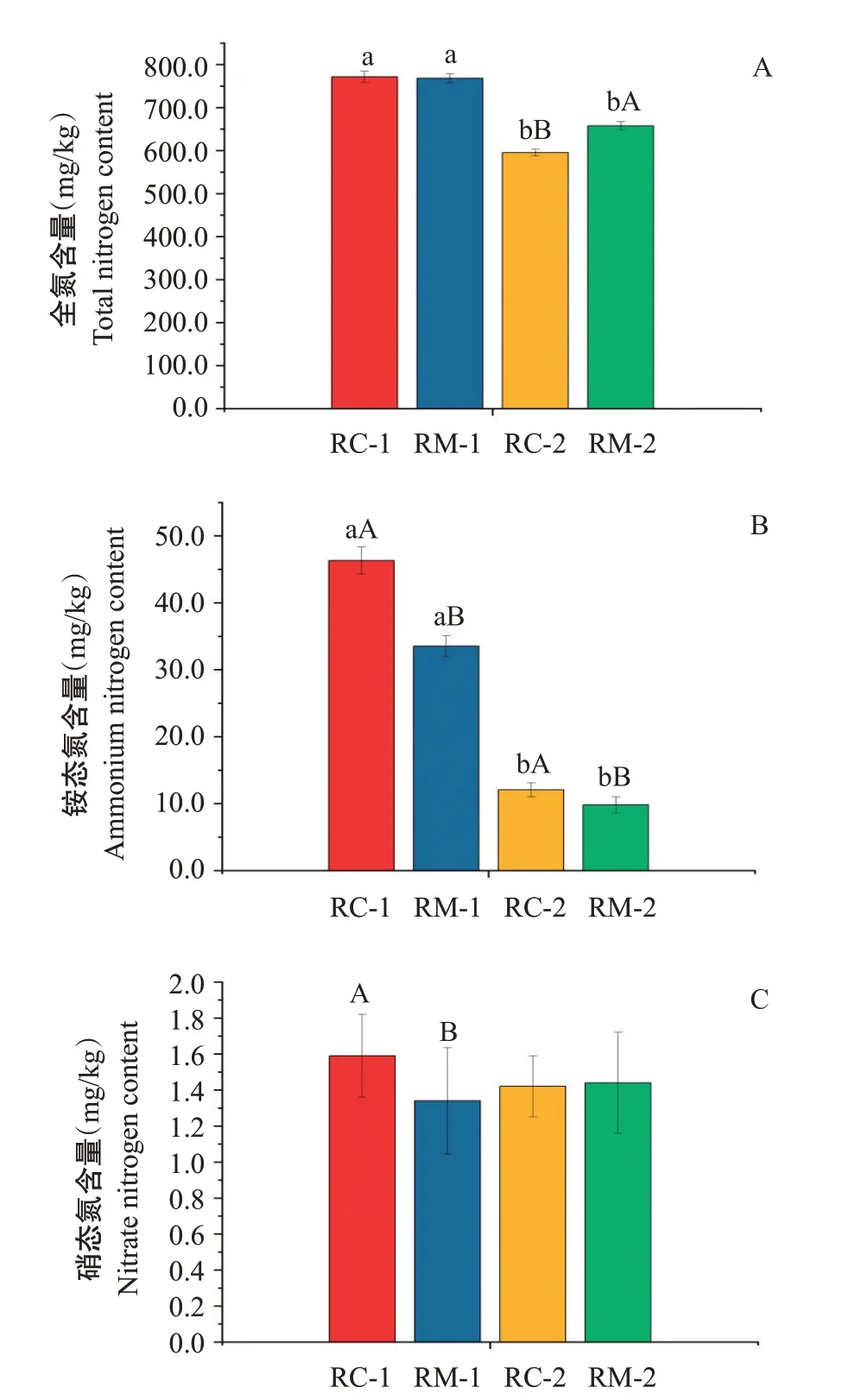
與試驗初期相比,試驗末期時RC和RM模式的土壤總氮、銨態氮含量均明顯下降,而硝態氮含量變化較小,且RC模式的土壤全氮含量下降幅度大于RM模式。
2.2 2種稻蝦共作模式對耕層10~20 cm土層土壤有機氮礦化參數的影響
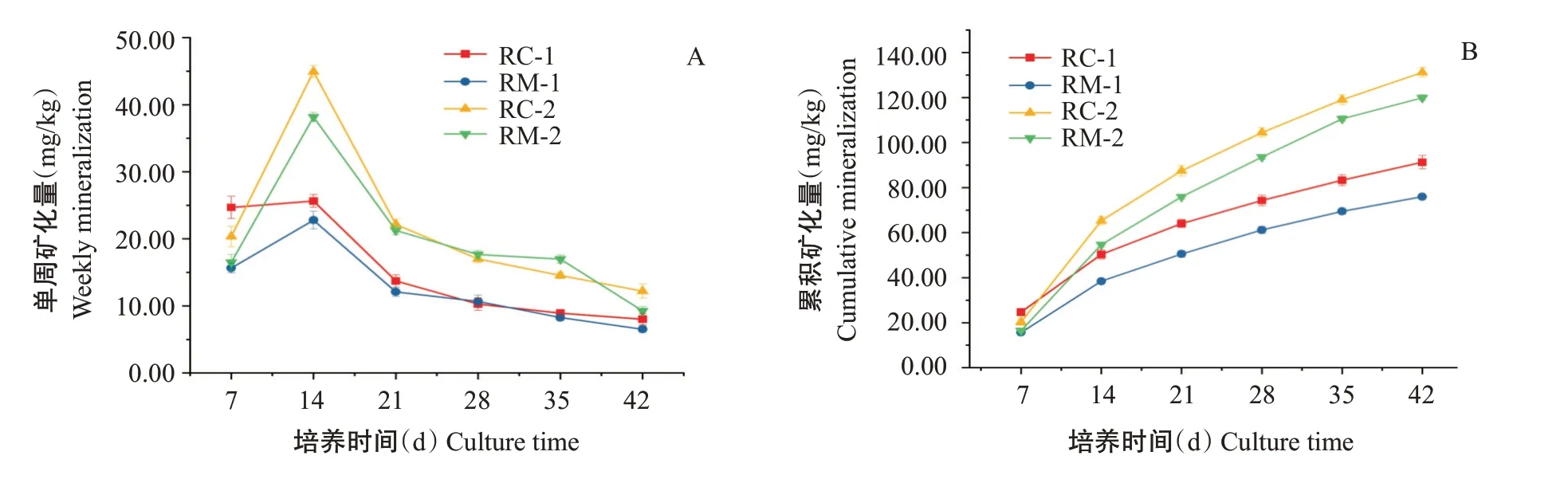
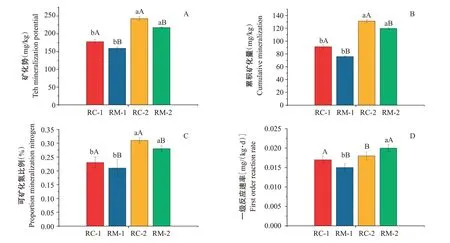
單周礦化量和累積礦化量變化曲線(圖3)顯示,連續淹水培養初期(0~14 d),RC-1、RM-1、RC-2和RM-2的單周礦化量均隨培養時間的延長呈增加趨勢,在第14 d達最大值,而后逐漸減小。20 ℃室內培養條件下,培養初、末期2種模式的累積氮礦化量表現為RC-2(131.21±2.12 mg/kg)>RM-2(119.85±1.36 mg/kg)>RC-1(91.27±2.96 mg/kg)>RM-1(76.00±1.16 mg/kg)。初、末期RC模式土壤累積氮礦化量均高于RM模式,且2種模式下試驗末期土壤的累積氮礦化量較試驗初期均明顯上升。圖4顯示,初、末期RC模式礦化勢、累積氮礦化量、可礦化氮比例均顯著高于RM模式。末期土壤中的礦化勢、累積氮礦化量、可礦化氮比例較初期均明顯上升,一級反應速率升高,但僅RM模式達顯著差異水平。
2.3 2種稻蝦共作模式對耕層10~20 cm土層土壤微生物群落結構特征的影響
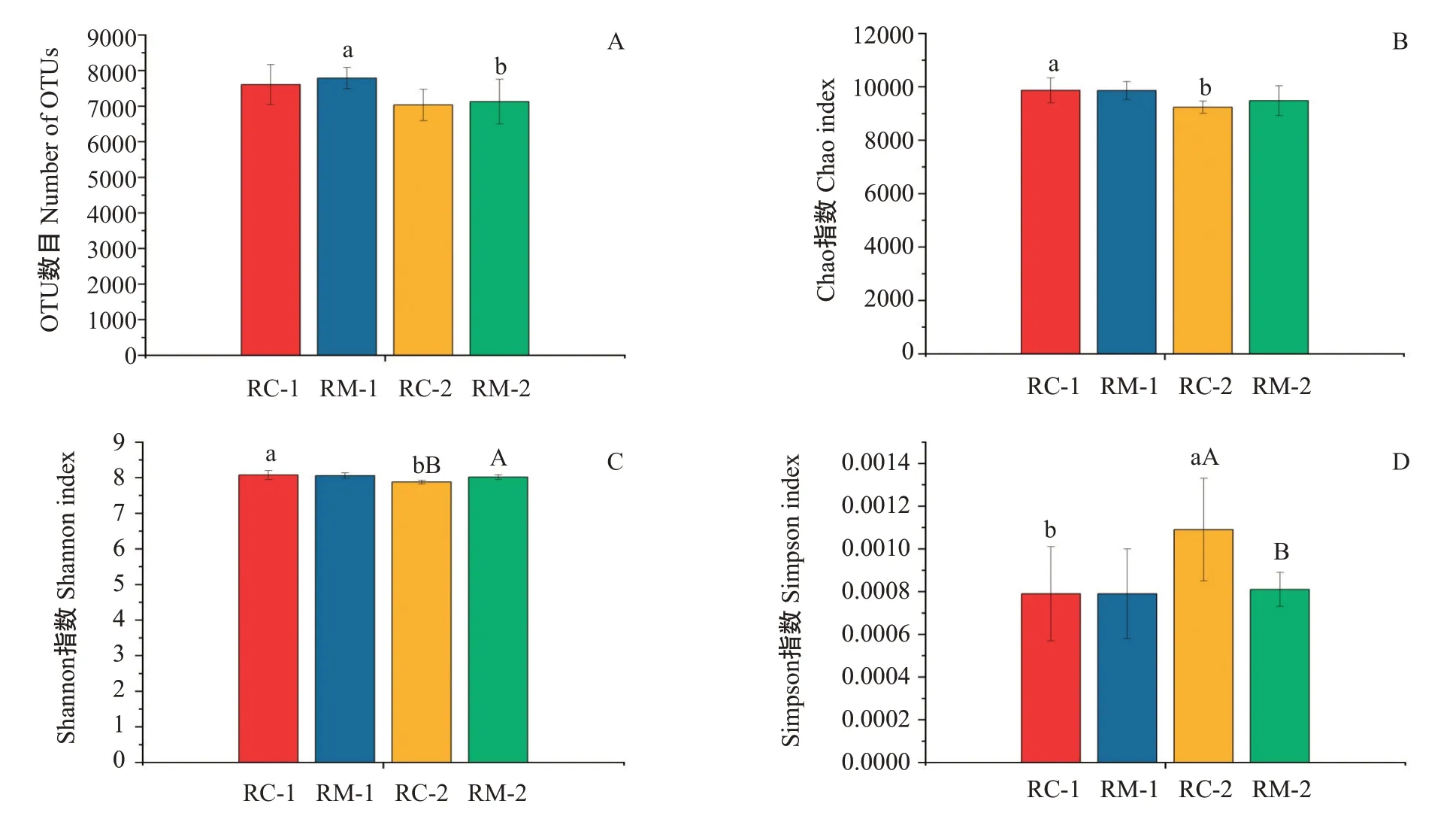
2.3.1 對稻田土壤微生物群落多樣性的影響 Al‐pha多樣性分析結果(圖5)顯示,RC-1與RM-1間OTU數目、Chao指數、Shannon指數、Simpson指數均無顯著差異;RC-2與RM-2間Shannon指數和Simpson指數存在顯著差異,Shannon指數RC-2低于RM-2,Simp‐son指數RC-2高于RM-2。結果表明,在試驗初期,RC模式與RM模式優勢菌群豐度、菌群多樣性均無明顯差異;而試驗末期菌群多樣性存在顯著差異,RC模式顯著低于RM模式。
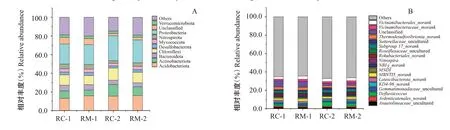
2.3.2 對稻田土壤微生物群落組成的影響 門分類水平上,2種共作模式稻田土壤微生物群落組成較相似,但不同采樣時期微生物組成有所差異。相對豐度≥1%的門類有16個,約占各樣品序列總數的93.0%。由圖6-A可看出,微生物門分類水平上相對豐度排前10的分別為變形菌門(Proteobacteria)、疣微菌門(Verrucomicrobiota)、酸桿菌門(Acidobacter‐iota)、粘菌門(Myxococcota)、脫硫桿菌門(Desulfo‐bacterota)、綠彎菌門(Chloroflexi)、硝化螺旋菌門(Nitrospirota)、擬桿菌門(Bacteroidota)、放線菌門(Actinobacteriota)和Unclassifild。其中,變形菌門為土壤中的最優勢菌群,相對豐度為21.4%~26.7%;其次為酸桿菌門、放線菌門、綠彎菌門和擬桿菌門,相對豐度分別為13.0%~16.0%、6.1%~12.4%、9.7%~13.0%、4.3%~6.1%。微生物屬分類水平上(圖6-B),相對豐度≥1.0%的微生物菌屬共有18個,約占樣品豐度的35.0%,其中屬于綠彎菌門的共有4個菌屬,屬于變形菌門的共有3個菌屬,屬于酸桿菌門的共有3個菌屬,屬于硝化螺旋菌門的共有2個菌屬,剩余6個則屬于其他門類。其中,為最優勢菌屬,相對豐度為2.4%~3.5%,其次為、、_及,相對豐度分別為1.0%~4.7%、2.1%~3.0%、1.5%~2.7%和1.8%~2.5%。
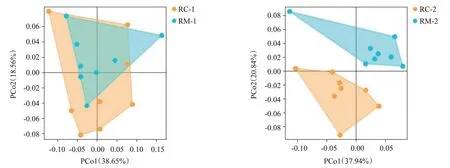
微生物群落結構主坐標分析結果(圖7)顯示,初期PCo1為38.65%,PCo2為18.56%,二者之和大于50%;末期PCo1為37.94%,PCo2為20.84%,二者之和大于50%;兩個時期均具有代表性。在初期,RC模式與RM模式各樣本位點在PCoA分析圖上距離較接近,各位點之間連線所構成的區域有重疊部分,表明2種養殖模式間土壤微生物菌群結構較相似;而在末期,2種養殖模式各樣本位點距離較遠,無明顯重疊區域,表明2種養殖模式間土壤微生物菌群組成結構明顯不同。
在門分類水平和屬分類水平上對相對豐度排前10的物種(Unclassified除外)的相對豐度數值進行-test檢驗,結果顯示(圖8-A),門水平上,試驗初期RC模式酸桿菌門相對豐度顯著低于RM模式,綠彎菌門相對豐度顯著高于RM模式;末期RC模式脫硫桿菌門、硝化螺旋菌門相對豐度顯著低于RM模式,而綠彎菌門、放線菌門相對豐度顯著高于RM模式。在RC模式中,末期黏菌門、脫硫桿菌門、硝化螺旋菌門、擬桿菌門相對豐度均顯著低于試驗初期,而變形菌門、放線菌門相對豐度均顯著高于初期;在RM模式中,硝化螺旋菌門、擬桿菌門相對豐度均顯著低于初期,而變形菌門、綠彎菌門、放線菌門相對豐度均顯著高于初期。
在屬水平上(圖8-B),試驗初期RC模式uncultured、、、的相對豐度均顯著低于RM模式,的相對豐度顯著高于RM模式。試驗末期RC模式uncultured、的相對豐度均顯著低于RM模式,而的相對豐度顯著高于RM模式。RC模式中,試驗末期的相對豐度顯著低于試驗初期,而、的相對豐度顯著高于試驗初期;RM模式中,試驗末期的相對豐度顯著低于試驗初期,而、的相對豐度顯著高于試驗初期。
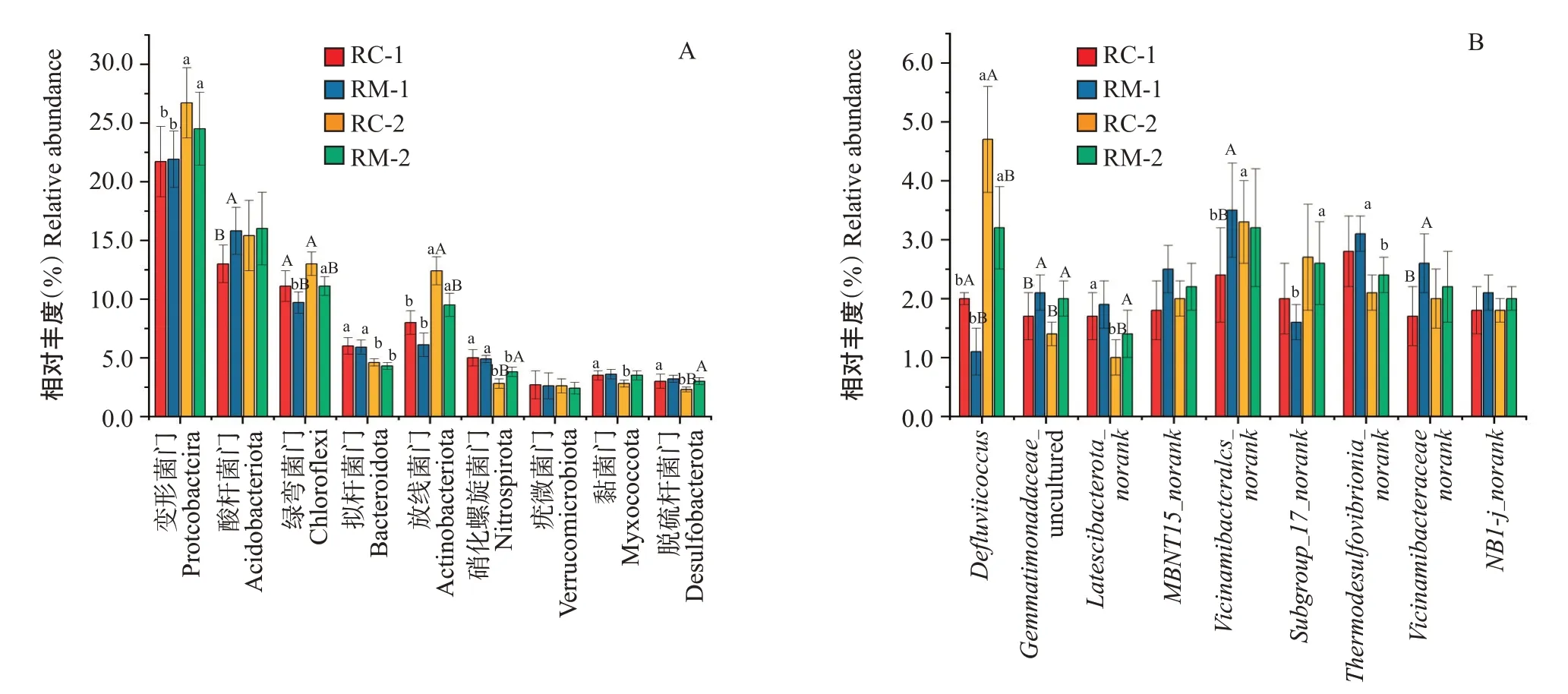
門分類水平上,同一采樣時期,2種模式之間相對豐度差異顯著的微生物門類由初始期的2門(酸桿菌門和綠彎菌門)增加到末期的4門(放線菌門、脫硫桿菌們、粘菌門和硝化螺旋菌門)。屬分類水平的變化與門分類水平類似,表現為相同采樣時期2模式間有差異的菌屬所對應的菌門由初期的4個門減少至末期的3個門。
2.3.3 土壤微生物類群與土壤氮礦化參數、土壤氮素的相關分析 對土壤微生物門分類水平(相對豐度前10)物種的相對豐度與土壤全氮、銨態氮、硝態氮、累積氮礦化量、礦化勢、可礦化氮比例及一級反應速率進行相關分析(圖9-A)。氮素含量相關性結果顯示,全氮含量與擬桿菌門、脫硫桿菌門、粘菌門、硝化螺旋菌門呈極顯著正相關(<0.01,下同),與放線菌門、綠彎菌門、變形菌門呈極顯著負相關;銨態氮含量與擬桿菌門、硝化螺旋菌門呈極顯著正相關,與脫硫桿菌門、粘菌門呈顯著正相關,與放線菌門、變形菌門呈極顯著負相關;硝態氮含量與各菌門均無顯著相關性。礦化參數相關性結果顯示,除一級反應速率外,累積氮礦化量、礦化勢和可礦化氮比例與放線菌門、綠彎菌門、變形菌門呈極顯著正相關,與擬桿菌門、脫硫桿菌門、粘菌門、硝化螺旋菌門呈極顯著負相關。表明除疣微菌門外,土壤全氮和銨態氮含量會影響其余門類土壤微生物的相對豐度,且相關門類微生物均能顯著影響土壤有機氮的礦化參數。
在屬分類水平中(圖9-B),、、與累積氮礦化量、礦化勢、可礦化氮比例和一級反應速率均為極顯著正相關;、與累積氮礦化量、礦化勢、可礦化氮比例和一級反應速率均為極顯著負相關;與累積氮礦化量、礦化勢和可礦化氮比例為顯著負相關,與一級反應速率無顯著相關性;屬僅與礦化勢存在顯著相關性;其余屬類微生物與土壤有機氮礦化參數均無顯著相關性。

3 討論
水稻主要吸收土壤中的無機氮供自身生長發育需要,而有機氮需經微生物的礦化作用分解為無機氮后,才能被水稻吸收利用(武際等,2012)。研究表明,土壤中的有機氮經銨化作用和硝化作用后最終生成銨態氮和硝態氮(王艷杰等,2005),因此,土壤銨態氮和硝態氮含量在一定程度上可反映有機氮的礦化作用強度。本研究結果表明,RC模式下土壤銨態氮含量在初期、末期均顯著高于RM模式,RC模式中土壤銨化作用更強。同時,初期2種模式間全氮含量無顯著差異,而末期RC模式顯著低于RM模式,表明紅螯螯蝦較羅氏沼蝦更能促進土壤有機氮的礦化。而硝態氮主要靠銨態氮的硝化作用生成,但試驗稻田長期處于淹水環境下,土壤硝化作用受到抑制,故初期、末期RC模式與RM模式間硝態氮含量無顯著差異,且同一模式不同時期硝態氮含量也無顯著差異,與李成芳等(2008)的研究結果一致。在水稻生長期間,其氮素來源于土壤現存氮素和外源氮素輸入(人工施肥等)(尹冬,2011;朱杰等,2018)。而本研究在蝦類養殖期間也未做施肥處理,水稻的氮素養分依靠有機氮礦化作用產生的銨態氮提供,因而至試驗末期土壤全氮含量和銨態氮含量顯著下降。
已有研究表明,底棲生物會影響土壤有機氮的降解。馬紅波(2001)研究發現,底棲生物通過影響沉積物環境而影響氮循環,加速沉積物礦化作用的進行,影響NH-N的生成速率;在Hulth等(2004)的研究結果中,底棲生物的生理活動會在沉積物中形成孔洞,使有氧界面在沉積物中延伸,進而影響有機氮的礦化;Hou等(2020)進一步研究發現,日本沼蝦生物擾動可改變沉積物氧化還原電位,促進有機物的降解,加速有機氮的礦化。本研究室內礦化培養試驗結果顯示,2種模式下末期土壤累積氮礦化量、礦化勢、可礦化氮比例均顯著高于試驗初期,且同一時期RC模式的累積氮礦化量、礦化勢、可礦化氮比例也均顯著高于RM模式,表明稻蝦共作末期土壤有機氮礦化作用較前期有所增強,RC模式中土壤有機氮礦化作用強度大于RM模式。在本研究中,蝦類作為稻田中最主要的底棲生物,除攝食投喂的餌料外,還會攝食植物殘體、有機碎屑等,可加速土壤中有機質的降解(Ferris et al.,1998),其攝食行為也會對土壤造成一定的攪動,降低土壤團聚體大小,進一步增強有機氮的礦化作用(王根林等,2009)。同時,較羅氏沼蝦而言,紅螯螯蝦自身還有掘穴行為,可連通表層和中層土壤,改善中層土壤的通氣狀況,其水氣條件等能更好地滿足好氧微生物生長(Kravchenko et al.,2017),因而更能提高有機氮的礦化作用。另外,底棲生物的攝食、爬行等生命活動也會對土壤微生物群落的結構、活性和多樣性造成影響(Laverock et al.,2010;Rugenski et al.,2012;Delmont et al.,2014;宋宇等,2020)。稻蝦共作期間,紅螯螯蝦、羅氏沼蝦的不同生活習性為土壤微生物營造了不同的生存環境,使得末期2種模式間微生物群落結構和多樣性產生明顯區別。
除底棲生物外,土壤微生物群落的也是有機氮礦化作用的重要影響因子,微生物的種類、結構、功能與有機氮的分解、礦化密切相關(李輝信等,2004;張俊清等,2004)。本研究中,放線菌門、綠彎菌門、變形菌門均與礦化作用相關參數呈極顯著正相關,而擬桿菌門、脫硫桿菌門、硝化螺旋菌門與礦化參數呈極顯著負相關。前人研究表明,變形菌門是一種r策略細菌(富營養菌),能加速土壤有機質分解,在土壤氮素循環中發揮重要作用(徐凡迪等,2021);放線菌門部分屬類可參與碳、氮循環,降解木質素,加速土壤植物殘體的分解,降低土壤木質素總量和氮總量的比值,促進有機氮的礦化(謝長校等,2015;Hua et al.,2021);綠彎菌門部分屬類以有機質為碳源,可加速有機質降解(Davis et al.,2011),對土壤有機氮的礦化有促進作用;擬桿菌門與土壤有機碳和氮素含量有較好關聯性,可反映有機質的降解程度,高有機質水平時相對豐度較高(Fierer et al.,2012;趙雅姣等,2020);而脫硫桿菌門部分屬類可將有機質完全氧化成CO或部分氧化為乙酸鹽,降低其余微生物的碳源利用能力,從而抑制有機氮的礦化;硝化螺旋菌門部分屬類微生物為化能無機自養,通過亞硝酸鹽氧化還原酶介導的亞硝酸鹽氧化生成硝酸鹽來獲取能量,抑制土壤有機氮的礦化過程(Liang et al.,2021)。在試驗末期,2種模式土壤中放線菌門、綠彎菌門、變形菌門相對豐度均上升,擬桿菌門、脫硫桿菌門、硝化螺旋菌門相對豐度均下降。由此可知,稻蝦共作模式能提升利于有機氮礦化的微生物相對豐度,進而提高土壤有機氮礦化作用的強度。進一步對末期2種模式間以上菌門的相對豐度進行比較分析,結果顯示RC模式放線菌門、綠彎菌門相對豐度顯著高于RM模式,而脫硫桿菌門、硝化螺旋菌門顯著高于RM模式。結合微生物相對豐度與礦化參數的相關分析結果可知,與有機氮礦化作用相關的微生物在不同模式中的相對豐度不同,是導致2種模式間有機氮礦化參數產生顯著差異的直接原因。
4 結論
蝦類的生活習性不同造就了不同的土壤環境,進而影響土壤微生物的群落結構組成,導致2種稻蝦共作模式間耕層下部10~20 cm土層土壤有機氮的礦化作用產生差異。在江蘇省靖江市,與稻—羅氏沼蝦共作模式相比,稻—紅鰲螯蝦共作模式下的稻田土壤有機氮的礦化作用更強,礦化勢等相關參數更高,具有較高的供氮潛力,更有利于水稻的生長。
