信號肽及發(fā)酵條件優(yōu)化促進(jìn)膠原蛋白在谷氨酸棒桿菌中分泌表達(dá)
2022-08-18 04:17:46齊靜靜范炳森張萌許菲
食品與發(fā)酵工業(yè) 2022年15期
關(guān)鍵詞:生長
齊靜靜,范炳森,張萌,許菲
(江南大學(xué) 生物工程學(xué)院,工業(yè)生物技術(shù)教育部重點(diǎn)實驗室,江蘇 無錫,214122)
膠原蛋白是哺乳動物體內(nèi)含量最多的一類蛋白質(zhì),占蛋白質(zhì)總量的25%~30%,是動物結(jié)締組織和細(xì)胞外基質(zhì)的重要組成成分。膠原蛋白由3條左旋多脯氨酸α鏈以相錯一個殘基的形式相互纏繞形成右手三股螺旋[1],典型的三股螺旋結(jié)構(gòu)決定了其具有良好的生物力學(xué)性能和生物相容性[2],在醫(yī)藥組織工程、化妝品、食品等領(lǐng)域[3]均有廣泛的應(yīng)用,2019年全球膠原蛋白市場規(guī)模達(dá)153.56億美元,并保持穩(wěn)步增長的趨勢。目前市面上的膠原蛋白主要通過動物提取獲得,具有潛在的免疫原性,有一定的安全隱患;此外,也可通過轉(zhuǎn)基因動植物表達(dá)和化學(xué)合成獲得,但往往價格昂貴、產(chǎn)量低,難以大批量生產(chǎn)。而微生物發(fā)酵表達(dá)重組膠原蛋白由于具有安全性高、成本低、產(chǎn)量高、序列設(shè)計靈活等優(yōu)點(diǎn)[4],近年來受到了越來越廣泛的關(guān)注,市場潛力大。
目前重組膠原蛋白常用的表達(dá)系統(tǒng)有大腸桿菌(Escherichiacoli)和畢赤酵母(Pichiapastoris),大腸桿菌具有發(fā)酵周期短、表達(dá)量高、便于遺傳操作等優(yōu)點(diǎn),是蛋白質(zhì)異源表達(dá)最廣泛的宿主菌。利用大腸桿菌表達(dá)來源于化膿性鏈球菌(Streptococcuspyogenes)的膠原蛋白Scl2[5],即使缺乏脯氨酸羥化酶修飾,仍能形成穩(wěn)定的三股螺旋結(jié)構(gòu),且在接近、甚至略高于37 ℃穩(wěn)定性較好,在搖瓶水平膠原蛋白V-CL的最優(yōu)產(chǎn)量為300 mg/L,補(bǔ)料分批發(fā)酵最優(yōu)產(chǎn)量為19 g/L[6-7];通過共表達(dá)伴侶蛋白、響應(yīng)面優(yōu)化等策略可以促進(jìn)單鏈類人源膠原蛋白在大腸桿菌中的異源表達(dá)[8-9],其中類人源II型膠原蛋白產(chǎn)量達(dá)到14.2 g/L[10]。然而由于大腸桿菌外膜的屏障作用,外源蛋白多為胞內(nèi)表達(dá),且易形成包涵體,此外,大量內(nèi)毒素的存在也為后期純化帶來壓力。酵母作為真核表達(dá)系統(tǒng),可進(jìn)行翻譯后修飾,并且無致病性、易進(jìn)行高密度發(fā)酵。利用畢赤酵母分泌表達(dá)單鏈重組人源膠原蛋白的研究比較成熟,人源Ш型膠原蛋白α1鏈的膠原域片段經(jīng)親水性改造及補(bǔ)料分批發(fā)酵條件優(yōu)化,產(chǎn)量提高至19.49 g/L[11];其膠原域全長在畢赤酵母GS115的胞外分泌產(chǎn)量最高達(dá)3.36 g/L[12]。然而由于缺少促進(jìn)三股螺旋折疊的C-端前導(dǎo)肽,重組膠原蛋白只能以單鏈的形式分泌到胞外。通過表達(dá)帶有C-端前導(dǎo)肽的膠原蛋白序列和共表達(dá)羥化酶可以促進(jìn)人源膠原蛋白在畢赤酵母中折疊形成三股螺旋[13],但是會導(dǎo)致其表達(dá)量下降和在內(nèi)質(zhì)網(wǎng)中的積累[14];即使僅截取9 kDa的膠原域片段并將C-端前導(dǎo)肽替換為分子量低的折疊域仍無法在畢赤酵母分泌具有三股螺旋結(jié)構(gòu)的膠原蛋白[15]。
谷氨酸棒桿菌(Corynebacteriumglutamicum)是公認(rèn)的符合食品安全標(biāo)準(zhǔn)的宿主,不含內(nèi)毒素,可應(yīng)用于氨基酸、有機(jī)酸、藥用蛋白等的生產(chǎn),其含有豐富的信號肽序列和單層膜結(jié)構(gòu),且胞外很少檢測到蛋白酶活性,有利于異源蛋白的分泌表達(dá)。目前研究表明其具有2條主要分泌途徑,通過不同信號肽使蛋白質(zhì)穿過細(xì)胞膜,分別是分泌未折疊蛋白的Sec依賴型和轉(zhuǎn)運(yùn)折疊蛋白的Tat依賴型。對C.glutamicumATCC 13032的胞外蛋白組進(jìn)行研究,發(fā)現(xiàn)在搖瓶水平,分泌型蛋白Cg2052的含量最高[16];對該菌株在高密度培養(yǎng)條件下的分泌組分析,發(fā)現(xiàn)胞外蛋白Cg1514占比最高[17],以Cg1514的信號肽介導(dǎo)分泌淀粉酶的產(chǎn)量達(dá)786.2 mg/L;PorB是谷氨酸棒桿菌中4種孔道蛋白之一,對陰離子選擇型細(xì)胞壁通道的形成起重要作用,可以有效地作為錨定蛋白用于細(xì)胞表面展示,而且其信號肽介導(dǎo)的Sec依賴性分泌途徑可以高效分泌外源蛋白,介導(dǎo)M18 scFv的分泌量達(dá)68 mg/L[18];CspB作為幾種谷氨酸棒桿菌菌株的主要分泌蛋白和表面蛋白,其信號肽作為Sec類型的強(qiáng)信號肽用于多種外源蛋白的分泌表達(dá)[19]。對C.glutamicumR基因組分析發(fā)現(xiàn),由CgR0949介導(dǎo)的Tat依賴型分泌途徑分泌的α-淀粉酶活性最高[20];后續(xù)與Ptac啟動子結(jié)合CgR0949進(jìn)行(green fluorescent protein,GFP)分泌表達(dá),產(chǎn)量可達(dá)2.8 g/L;此外,有研究報導(dǎo)來源于大腸桿菌的TorA信號肽可以有效地介導(dǎo)GFP在C.glutamicumATCC 13869中經(jīng)Tat依賴型途徑的分泌表達(dá)[21],過表達(dá)Tat途徑的TatABC蛋白的同時用TorA信號肽介導(dǎo)分泌的pro-TG蛋白的量大于經(jīng)Sec途徑的產(chǎn)量[22]。
為了搭建谷氨酸棒桿菌分泌表達(dá)重組膠原蛋白的平臺,本研究選用來自Streptococcuspyogenes膠原蛋白Scl2中穩(wěn)定性較高的B片段(約占CL膠原域的1/3)[6]為研究對象,在B前端插入輔助膠原折疊形成三股螺旋結(jié)構(gòu)的V-domain和胰蛋白酶切位點(diǎn)LVPRGS,通過在V-B前端引入不同信號肽,并進(jìn)行發(fā)酵條件優(yōu)化,對目的蛋白進(jìn)行分離純化和表征。本研究為重組膠原蛋白異源表達(dá)新宿主的選擇提供基礎(chǔ),也為谷氨酸棒桿菌分泌表達(dá)信號肽的選擇提供參考。
1 材料與方法
1.1 材料
1.1.1 菌株與質(zhì)粒
大腸桿菌E.coliJM109用于載體構(gòu)建和擴(kuò)增,谷氨酸棒狀桿菌C.glutamicumATCC 13032用于目的基因表達(dá);大腸桿菌-谷氨酸棒桿菌穿梭表達(dá)質(zhì)粒pXMJ19、克隆有V-B基因的質(zhì)粒pCold-III-V-B,本實驗室保藏。擴(kuò)增不同信號肽構(gòu)建重組質(zhì)粒pXMJ19-SPs-V-B所用的引物(表1)蘇州金唯智生物科技有限公司。

表1 本研究使用的引物Table 1 Primers used in this study
1.1.2 主要試劑和培養(yǎng)基
蛋白Marker、Prime STAR?HS(Premix)、Blunting Kination Ligation (BKL) Kit、T4 DNA Ligase,TaKaRa公司;異丙基-β-D硫代半乳糖苷(isopropyl-β-D-thiogalactopyranoside,IPTG)、氯霉素,上海生工生物工程股份有限公司;腦心浸出液肉湯(brain heart infusion,BHI),青島海博生物;其他試劑均為國產(chǎn)分析純。
LB培養(yǎng)基(g/L):酵母粉5,蛋白胨10,NaCl 10,固體培養(yǎng)基含瓊脂糖15;氯霉素作為大腸桿菌抗性篩選時,終質(zhì)量濃度為30 μg/mL。
種子培養(yǎng)基:BHI培養(yǎng)基。
發(fā)酵培養(yǎng)基1(g/L)[23]:(NH4)2SO420,玉米漿干粉20,K2HPO41,KH2PO41,MgSO40.25,3-(N-嗎啉基)丙磺酸鈉鹽42,葡萄糖20,pH 7.0。
發(fā)酵培養(yǎng)基2:BHI培養(yǎng)基。
發(fā)酵培養(yǎng)基3(g/L):葡萄糖150,(NH4)2SO440,KH2PO41,MgSO40.5,玉米漿15,酵母膏1,蛋白胨0.5,MnSO4·H2O 0.01,F(xiàn)eSO4·7H2O 0.01,焦磷酸硫銨素0.001,生物素0.001,輕質(zhì)碳酸鈣20,pH 7.2。
氯霉素在谷氨酸棒桿菌中作為抗性篩選時,終質(zhì)量濃度為15 μg/mL。
1.2 實驗方法
1.2.1 表達(dá)載體的構(gòu)建
pXMJ19-V-B的構(gòu)建:將質(zhì)粒pCold-III-V-B和空載pXMJ19均用Hind Ⅲ和EcoR I雙酶切,分別回收目的產(chǎn)物,經(jīng)純化后,用T4 DNA連接酶于16 ℃過夜連接,并轉(zhuǎn)化至E.coliJM109中,然后涂布于含30 μg/mL氯霉素的LB平板,挑取轉(zhuǎn)化子于37 ℃培養(yǎng),提取質(zhì)粒,將測序驗證正確的重組質(zhì)粒命名為pXMJ19-V-B。
pXMJ19-SPs-V-B的構(gòu)建:以質(zhì)粒pXMJ19-V-B為模板,設(shè)計引物CspB-F/R、PorB-F/R、Cg1514-F/R、CgR0949-F/R、Cg2052-F/R、TorA-F/R,通過PCR獲得帶有6種不同信號肽的重組質(zhì)粒。PCR體系為:Prime STAR HS (Premix) 25 μL、上下游引物1 μL、模板1 μL,加水至50 μL。PCR擴(kuò)增條件為:98 ℃ 2 min;98 ℃ 10 s,55~68 ℃ 10 s,72 ℃ 8 min,30個循環(huán);72 ℃ 10 min。產(chǎn)物回收后做磷酸化處理,16 ℃過夜連接,轉(zhuǎn)化至E.coliJM109,然后涂布于含30 μg/mL氯霉素的LB平板,培養(yǎng)12~14 h挑取單菌落培養(yǎng)送基因測序。測序驗證正確后,將質(zhì)粒命名為pXMJ19-SPs-V-B。
1.2.2 谷氨酸棒桿菌的轉(zhuǎn)化
將測序正確的重組質(zhì)粒pXMJ19-SPs-V-B電擊轉(zhuǎn)化進(jìn)C.glutamicumATCC 13032感受態(tài)細(xì)胞,通過抗性篩選得到陽性轉(zhuǎn)化子,并命名為C.glutamicumATCC 13032/pXMJ19-SPs-V-B。為了能夠更好地反映信號肽的分泌表達(dá)情況,同時構(gòu)建胞內(nèi)表達(dá)C.glutamicumATCC 13032/pXMJ19-V-B作為對照菌株。
1.2.3 重組菌誘導(dǎo)表達(dá)的初始條件
將保存重組菌的甘油管在BHI平板上劃線,30 ℃,培養(yǎng)24~32 h,然后挑取單菌落至5 mL的BHI中,30 ℃,200 r/min過夜培養(yǎng);取一定量的種子液轉(zhuǎn)接至發(fā)酵培養(yǎng)基[23],使初始OD600為0.1,28 ℃,200 r/min培養(yǎng)3 h后添加IPTG至終濃度為0.8 mmol/L進(jìn)行誘導(dǎo),培養(yǎng)48 h。
1.2.4 重組菌發(fā)酵條件的優(yōu)化
(1)信號肽的優(yōu)化:將C.glutamicumATCC 13032/pXMJ19-SPs-V-B和C.glutamicumATCC 13032/pXMJ19-V-B分別誘導(dǎo)表達(dá),每隔8 h取樣靜置15 min測OD600,并繪制生長曲線,誘導(dǎo)結(jié)束后,收集發(fā)酵上清液,濃縮20倍進(jìn)行SDS-PAGE分析,比較目的條帶的粗細(xì),確定最優(yōu)信號肽。
(2)發(fā)酵培養(yǎng)基的優(yōu)化:選擇最優(yōu)信號肽對應(yīng)菌株,分別用發(fā)酵培養(yǎng)基1、2、3進(jìn)行誘導(dǎo)表達(dá),誘導(dǎo)結(jié)束后取上清液濃縮后進(jìn)行SDS-PAGE分析,確定最佳發(fā)酵培養(yǎng)基。
(3)起始誘導(dǎo)OD600的優(yōu)化:將種子液轉(zhuǎn)至最佳發(fā)酵培養(yǎng)基,在OD600分別為2、6、10、14時,加入IPTG誘導(dǎo),48 h后取上清液濃縮,通過SDS-PAGE確定最佳誘導(dǎo)時機(jī)。
(4)誘導(dǎo)時長的優(yōu)化:在最佳起始誘導(dǎo)OD600,加入IPTG分別誘導(dǎo)24、32、40、48、56、64 h,探究誘導(dǎo)時長對pXMJ19-SP-V-B表達(dá)量的影響。
(5)IPTG濃度的優(yōu)化:在最佳起始誘導(dǎo)OD600加入終濃度分別為0.2、0.5、0.8、1.1 mmol/L的IPTG,28 ℃誘導(dǎo)至最佳時長,研究IPTG濃度對pXMJ19-SP-V-B表達(dá)的影響。
(6)溶氧條件的優(yōu)化:在最優(yōu)條件基礎(chǔ)上,將誘導(dǎo)表達(dá)培養(yǎng)基的裝液量改為50 mL/500 mL(帶擋板搖瓶)、50 mL/500 mL、100 mL/500 mL(帶擋板搖瓶)、100 mL/500 mL,研究不同溶氧條件對表達(dá)的影響。
1.2.5 膠原蛋白的純化
在最優(yōu)發(fā)酵條件下,收集菌液,4 ℃、12 000 r/min離心25 min,取上清液,經(jīng)0.22 μm水系濾膜過濾。然后用His TrapTMHP 5 mL親和純化,先用5倍柱體積結(jié)合緩沖液A(20 mmol/L Na2HPO4、20 mmol/L NaH2PO4、500 mmol/L NaCl、10 mmol/L咪唑、pH 7.4)平衡,再以5 mL/min的流速上樣。上樣結(jié)束后,用洗脫緩沖液B(20 mmol/L Na2HPO4、20 mmol/L NaH2PO4、500 mmol/L NaCl、500 mmol/L咪唑、pH 7.4)進(jìn)行梯度洗脫獲得目的蛋白,并利用SDS-PAGE分析純化情況。
1.2.6 胰蛋白酶酶切
將純化后的膠原蛋白與胰蛋白酶以物質(zhì)的量比250∶1的比例混合,16 ℃過夜酶切后,用HiTrap Desalting做脫鹽處理,以超純水為流動相,流速為5 mL/min,經(jīng)SDS-PAGE驗證后進(jìn)行真空冷凍干燥。
1.2.7 圓二色譜檢測
將凍干后的樣品用10 mmol/L、pH 7.4的PB緩沖液溶解成1 mg/mL的溶液,4 ℃平衡48 h后進(jìn)行圓二色譜鑒定。全波長是在4 ℃下以1 nm為間隔測定190~260 nm的圓二色譜光譜,平均掃描時間為5 s。熱變曲線是通過監(jiān)測225 nm處的圓二色譜信號,以10 ℃/h的升溫速率從10 ℃升高到70 ℃獲得的,每個溫度下平衡8 s,熔融溫度(Tm)通過擬合熱變曲線的一階導(dǎo)數(shù)求得。
2 結(jié)果與分析
2.1 不同信號肽對重組菌表達(dá)的影響
信號肽種類不同,介導(dǎo)分泌外源蛋白的能力也有所差別。為了研究不同信號肽對重組膠原蛋白表達(dá)的影響,在目的基因前端分別引入信號肽CspB、PorB、Cg1514、CgR0949、Cg2052、TorA,構(gòu)建重組菌進(jìn)行誘導(dǎo)表達(dá)。如圖1-a所示,引入信號肽的菌株整體生長趨勢相近,在誘導(dǎo)期的前24 h,均保持較快的生長速度,積累較多菌體;誘導(dǎo)24~48 h,菌體生長速度明顯變緩,OD600維持在25左右;對照菌株V-B在誘導(dǎo)期的前24 h生長趨勢與其他菌株相似,但誘導(dǎo)24~40 h仍保持較高的生長速度,誘導(dǎo)40~48 h趨于平穩(wěn),OD600約為28。誘導(dǎo)結(jié)束后,取發(fā)酵上清液進(jìn)行SDS-PAGE分析,如圖1-b、圖1-c所示,與未插入信號肽的對照菌株相比,所有引入信號肽的菌株發(fā)酵上清液中均出現(xiàn)分子質(zhì)量約為23 kDa的條帶,高于目的蛋白的理論分子質(zhì)量17.82 kDa,這可能是由于膠原域含有大量的脯氨酸,導(dǎo)致膠原蛋白在SDS-PAGE中遷移速度較慢,表觀分子質(zhì)量約為理論分子質(zhì)量的1.4倍[24]。其中,Cg1514、CspB、PorB介導(dǎo)分泌蛋白的效果較好,尤其是PorB對應(yīng)目的蛋白條帶最粗,因此選擇PorB作為介導(dǎo)膠原蛋白V-B分泌的信號肽。

a-不同重組菌株的生長曲線;b-引入信號肽菌株的發(fā)酵上清液的SDS-PAGE圖;c-對照菌株(未插入信號肽)發(fā)酵上清液的SDS-PAGE圖(1~6分別對應(yīng)引入信號肽CspB、PorB、Cg1514、CgR0949、Cg2052、TorA的菌株;M: premixed protein marker)圖1 信號肽對菌體生長和蛋白表達(dá)的影響Fig.1 Effects of different signal peptides on bacterial growth and protein expression
谷氨酸棒桿菌ATCC 13032在搖瓶水平胞外含量最高的蛋白是Cg2052[16],而在高密度培養(yǎng)條件下胞外的Cg1514蛋白含量最高,本研究中信號肽Cg1514介導(dǎo)分泌膠原蛋白的量明顯高于信號肽Cg2052,這種差異可能是由于搖瓶培養(yǎng)條件的區(qū)別[17],也可能是由于宿主菌胞外蛋白含量的高低并不能完全決定其信號肽分泌外源蛋白能力的強(qiáng)弱。YIM等[17]對比了搖瓶水平信號肽Cg1514和PorB介導(dǎo)分泌內(nèi)切木聚糖酶的能力,發(fā)現(xiàn)Cg1514介導(dǎo)分泌目的蛋白的產(chǎn)量高;而在本實驗中PorB作為信號肽分泌V-B的量約為Cg1514的1.8倍,這可能和蛋白自身性質(zhì)有關(guān),如蛋白結(jié)構(gòu)和形態(tài)等[25],或者與目的蛋白和信號肽的適配性有關(guān)[26]。此外,從分泌途徑來說,作為Sec依賴型信號肽的Cg1514、CspB、PorB介導(dǎo)的膠原蛋白產(chǎn)量高于Tat依賴型的信號肽CgR0949和TorA,在最新的研究中利用谷氨酸棒桿菌分泌表達(dá)蛛絲蛋白,其主要分泌途徑為Sec依賴型[27],因此推測Sec依賴型信號肽介導(dǎo)的分泌途徑有利于膠原蛋白、蛛絲蛋白等纖維狀蛋白的分泌表達(dá)。
2.2 發(fā)酵培養(yǎng)基對C.glutamicum ATCC 13032/pXMJ19-PorB-V-B表達(dá)的影響
在微生物發(fā)酵培養(yǎng)的過程中,合適的營養(yǎng)供給是微生物快速生長和目的產(chǎn)物大量合成的基礎(chǔ)。為了獲得適合C.glutamicumATCC 13032/pXMJ19-PorB-V-B表達(dá)的發(fā)酵培養(yǎng)基,選取了3種谷氨酸棒桿菌常用培養(yǎng)基1、2、3進(jìn)行重組菌的發(fā)酵。如圖2-a所示,重組菌在3種發(fā)酵培養(yǎng)基中的生長狀況有明顯差異:在培養(yǎng)基2中,誘導(dǎo)16 h菌濃度達(dá)到最高,直到誘導(dǎo)結(jié)束一直維持在10左右,菌體生長受限,積累菌量最少;在培養(yǎng)基1中,菌體前期生長迅速,后期生長趨勢變緩,誘導(dǎo)24~48 h,OD600逐漸增加到25左右;發(fā)酵培養(yǎng)基3中菌體前期生長速度雖低于培養(yǎng)基1,但誘導(dǎo)40 h內(nèi)一直維持較高的增長速度,誘導(dǎo)48 h時OD600與培養(yǎng)基1相當(dāng)。

a-不同培養(yǎng)基下菌株的生長曲線;b-不同培養(yǎng)基下發(fā)酵上清液的SDS-PAGE圖(泳道1~3分別對應(yīng)培養(yǎng)基1、2、3)圖2 培養(yǎng)基條件對菌體生長和蛋白表達(dá)的影響Fig.2 Effects of medium conditions on bacterial growth and protein expression
取發(fā)酵上清液做SDS-PAGE鑒定,如圖2-b所示,培養(yǎng)基1中目的蛋白表達(dá)量最高,條帶最粗;培養(yǎng)基3次之;培養(yǎng)基2產(chǎn)量最低,幾乎看不到條帶。分析膠原蛋白在培養(yǎng)基1表達(dá)量高的原因可能包括:
(1)培養(yǎng)基1中特有的3-(N-嗎啉基)丙磺酸鈉鹽可提供pH在6.5~7.9范圍的緩沖環(huán)境,有利于菌體的快速生長和外源蛋白的表達(dá);(2)培養(yǎng)基1和培養(yǎng)基3雖然誘導(dǎo)結(jié)束菌濃度相近,但培養(yǎng)基1中的菌體前期生長速度快,積累了較多菌量,在發(fā)酵后期較培養(yǎng)基3更有利于目的蛋白的表達(dá)和積累。因此,選擇培養(yǎng)基1作為重組膠原蛋白V-B分泌表達(dá)的發(fā)酵培養(yǎng)基。
2.3 起始誘導(dǎo)OD600對C.glutamicum ATCC 13032/pXMJ19-PorB-V-B表達(dá)的影響
起始誘導(dǎo)OD600對重組菌合成目的蛋白有很大影響,當(dāng)菌體處于對數(shù)生長期,活力最強(qiáng),能很快適應(yīng)添加誘導(dǎo)劑后的環(huán)境,迅速進(jìn)入蛋白合成期。為了探究起始誘導(dǎo)OD600對C.glutamicumATCC 13032/pXMJ19-PorB-V-B表達(dá)的影響。如圖3-a所示,培養(yǎng)前4 h,菌種處于延滯期,生長緩慢,OD600約為2;培養(yǎng)4 h后開始進(jìn)入對數(shù)期,4~14 h為菌種的對數(shù)生長期,OD600從5提高至18;之后生長速度明顯變緩。因此,在種子液轉(zhuǎn)接至發(fā)酵培養(yǎng)基后,控制OD600分別達(dá)到2、6、10、14時,添加IPTG至終濃度為0.8 mmol/L進(jìn)行誘導(dǎo),28 ℃培養(yǎng)48 h,每隔8 h測定OD600并繪制生長曲線。如圖3-b所示,受起始OD600的影響,菌株在誘導(dǎo)表達(dá)前8 h,OD600仍維持較大差距。誘導(dǎo)16 h,差距明顯縮小,OD600達(dá)到22左右;誘導(dǎo)24 h后,菌體生長速度變緩,OD600以較低速度增長,誘導(dǎo)48 h,所有條件的最終OD600差別不大,都達(dá)到28左右。由圖3-c可知,誘導(dǎo)結(jié)束后,起始OD600為2和6的菌株對應(yīng)目的條帶較粗,蛋白表達(dá)量高。可能是由于起始OD600為10和14的菌株誘導(dǎo)劑添加時間較晚,前期培養(yǎng)基中營養(yǎng)物質(zhì)被大量消耗,影響后期目的蛋白合成。而OD600為2時,可觀察到菌體處于延滯期,故選取處于對數(shù)生長期的OD600=6為最佳起始誘導(dǎo)菌濃度。

a-未加誘導(dǎo)劑菌株的生長曲線;b-不同起始誘導(dǎo)OD600下菌株的生長曲線;c-不同起始誘導(dǎo)OD600下發(fā)酵上清液的SDS-PAGE圖(泳道1~4分別對應(yīng)起始誘導(dǎo)OD600為2、6、10、14;M-premixed protein marker)圖3 起始誘導(dǎo)OD600對菌體生長和蛋白表達(dá)的影響Fig.3 Effect of initial induction of OD600 on bacterial growth and protein expression
2.4 誘導(dǎo)時長對C.glutamicum ATCC13032/pXMJ19-PorB-V-B表達(dá)的影響
由圖3-b誘導(dǎo)期菌濃度變化情況發(fā)現(xiàn),誘導(dǎo)48 h,OD600仍有緩慢上升的趨勢,表明菌體仍在積累,誘導(dǎo)時間會影響菌量和目的蛋白的表達(dá)量。為了確定最佳誘導(dǎo)時長,在OD600=6時加入IPTG至終濃度為0.8 mmol/L進(jìn)行誘導(dǎo)。由圖4-a可看出,在誘導(dǎo)前16 h,菌體保持較快的生長速度,誘導(dǎo)16~48 h,菌體生長速度放緩,誘導(dǎo)48~64 h,OD600趨于穩(wěn)定,維持在28左右。從圖4-b可觀察到,在24~40 h,目的條帶亮度增加,目的蛋白表達(dá)量隨誘導(dǎo)時間的延長而增加;誘導(dǎo)時間超過40 h后,目的蛋白條帶沒有明顯的變化,蛋白未出現(xiàn)降解的跡象。考慮到隨著培養(yǎng)時間的延長,體系中營養(yǎng)物質(zhì)有限,會積累較多代謝產(chǎn)物,導(dǎo)致菌體衰退和老化,以及獲得更高的時空產(chǎn)量,故選取40 h為最佳誘導(dǎo)時長。

a-不同誘導(dǎo)時長菌株的生長曲線;b-不同誘導(dǎo)時長下發(fā)酵上清液的SDS-PAGE圖(泳道1~6分別對應(yīng)發(fā)酵時長24、32、40、48、56、64 h)圖4 誘導(dǎo)時長對菌體生長和蛋白表達(dá)的影響Fig.4 Effect of induction time on bacterial growth and protein expression
2.5 IPTG濃度對C.glutamicum ATCC 13032/pXMJ19-PorB-V-B表達(dá)的影響
IPTG作為半乳糖的結(jié)構(gòu)類似物,能夠誘導(dǎo)乳糖操縱子下游基因的表達(dá),其添加量對菌株生長及蛋白產(chǎn)量可能有一定影響。為了確定誘導(dǎo)劑最適添加濃度,在OD600=6時加入IPTG。從圖5-a可看出,IPTG濃度對菌體整體生長趨勢無太大影響。由圖5-b可知,IPTG終濃度為0.5 mmol/L時,目的蛋白條帶最粗,0.2 mmol/L次之;終濃度為0.8和1.1 mmol/L的目的條帶較淺,可能是由于較高的誘導(dǎo)劑濃度會造成菌體代謝壓力大而導(dǎo)致蛋白產(chǎn)量下降[28]。因此在本實驗中誘導(dǎo)劑IPTG的最佳工作濃度為0.5 mmol/L。
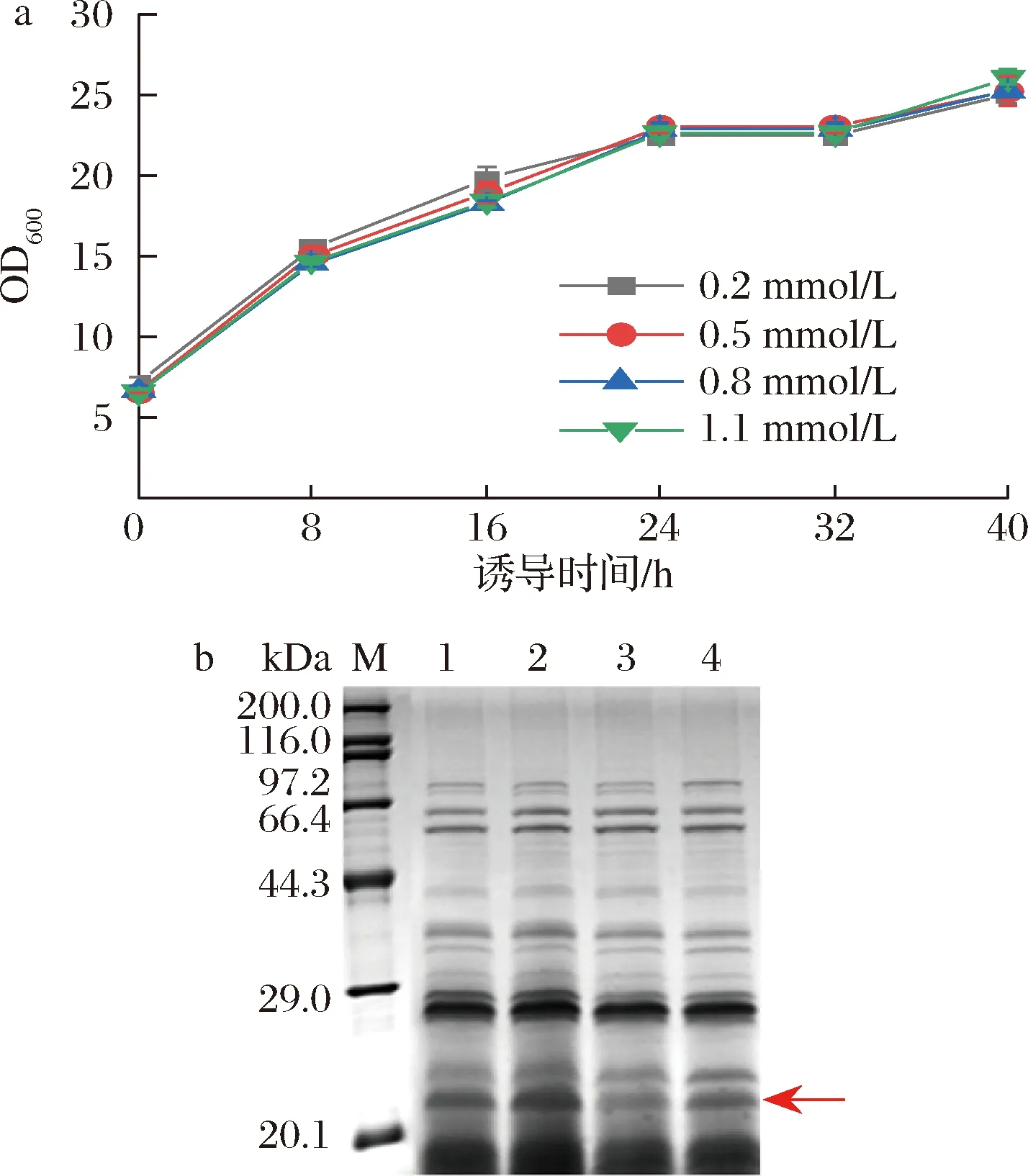
a-不同IPTG濃度下菌株的生長曲線;b-不同IPTG濃度下發(fā)酵上清液的SDS-PAGE圖(泳道1~4分別對應(yīng)IPTG濃度為0.2、0.5、0.8、1.1 mmol/L)圖5 IPTG濃度對菌體生長和蛋白表達(dá)的影響Fig.5 Effects of IPTG concentration on bacterial growth and protein expression
2.6 溶氧對C.glutamicum ATCC 13032/pXMJ19-PorB-V-B表達(dá)的影響
谷氨酸棒桿菌是嚴(yán)格的好氧型細(xì)菌,其生長代謝和產(chǎn)物合成均需要氧氣,因此氧氣濃度至關(guān)重要。由圖6-a可看出,在誘導(dǎo)相同時長下,帶擋板的搖瓶OD600比不帶擋板的高,且相同類型搖瓶,裝液量低的OD600較高,說明溶氧對菌體的生長狀況影響較大。誘導(dǎo)0~8 h,帶擋板搖瓶的菌體生長速度很快,約為相同裝液量不帶擋板搖瓶的2倍,其中裝液量為50 mL和100 mL的帶擋板搖瓶在誘導(dǎo)8 h的OD600分別超過50和43,之后OD600無太大變化,進(jìn)入穩(wěn)定期;不帶擋板搖瓶條件下的菌體OD600在誘導(dǎo)0~24 h穩(wěn)定增長,誘導(dǎo)24 h后,OD600保持平穩(wěn),誘導(dǎo)40 h后,裝液量50 mL和100 mL的搖瓶OD600分別達(dá)到40和28。如圖6-b所示,裝液量50 mL帶擋板的搖瓶目的條帶最粗。表明溶氧對菌體生長和蛋白表達(dá)有顯著影響,高的溶氧條件有利于菌體生長和外源蛋白表達(dá)。YANG等[29]在谷氨酸棒桿菌中表達(dá)腈水合酶,搖瓶培養(yǎng)最高OD600為15.6,酶活性為143.1 U/mL,在10 L發(fā)酵罐中的高密度發(fā)酵,OD600達(dá)到200,總酶活性達(dá)到1 432 U/mL,表明在發(fā)酵罐水平對溶氧的嚴(yán)格控制可以顯著提高谷氨酸棒桿菌中外源蛋白的表達(dá),對進(jìn)一步高密度培養(yǎng)有指導(dǎo)意義。
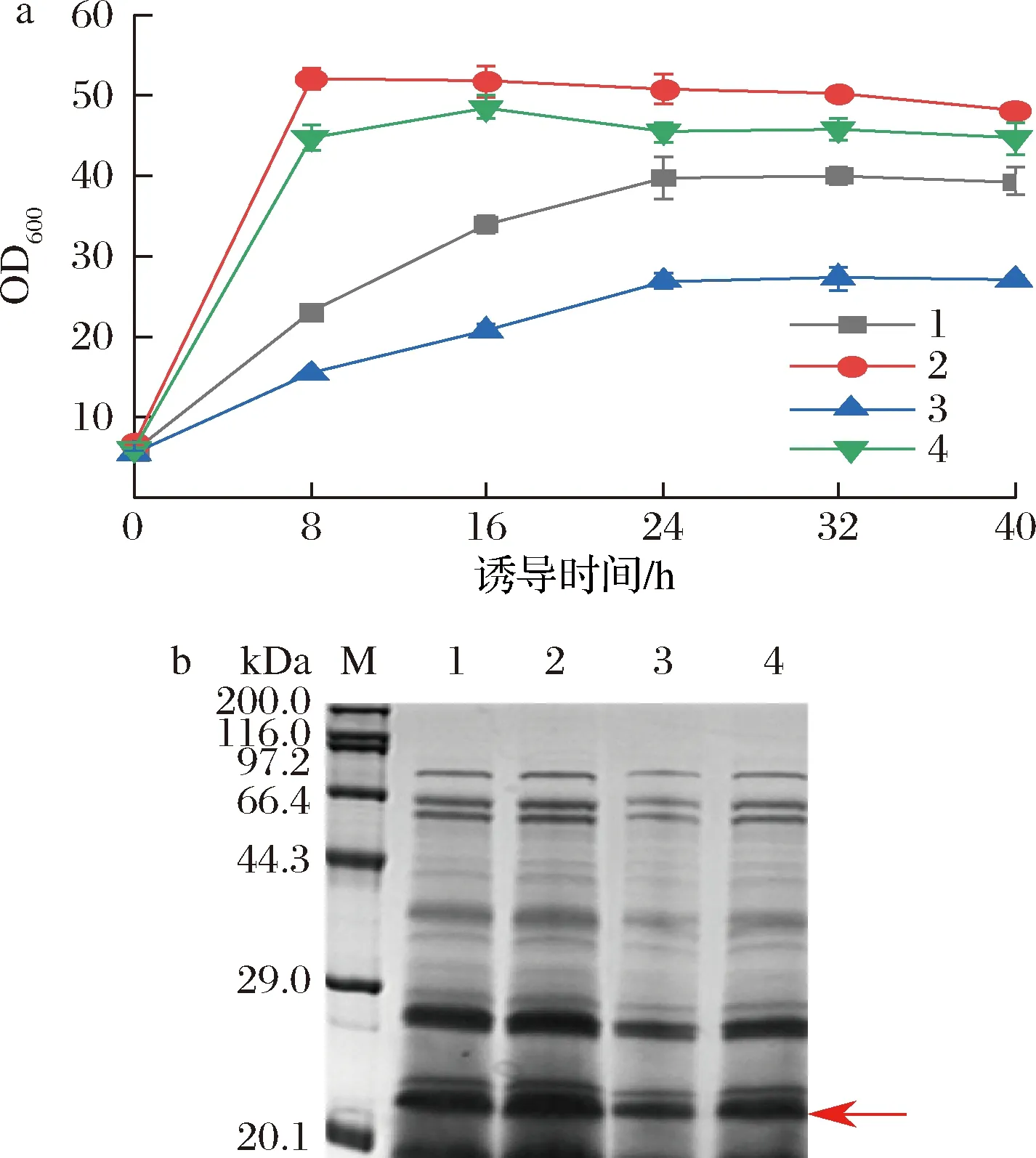
a-不同溶氧條件下的生長曲線;b-不同溶氧條件下發(fā)酵上清液的SDS-PAGE圖(1~4分別對應(yīng)50 mL/500 mL、50 mL/500 mL帶擋板搖瓶、100 mL/500 mL、100 mL/500 mL帶擋板搖瓶)圖6 溶氧對菌體生長和蛋白表達(dá)的影響Fig.6 Effects of dissolved oxygen on bacterial growth and protein expression
2.7 重組膠原蛋白V-B的表征
重組膠原蛋白V-B包含V domain和膠原域B,純化后經(jīng)胰蛋白酶消化,V domain被切成多個小肽段,而膠原域B如果正確折疊形成了具有剛性的三股螺旋結(jié)構(gòu),則不會被胰蛋白酶消化。由圖7-a可知,酶切前V-B分子質(zhì)量約為23 kDa,酶切后有明顯的單一條帶,分子質(zhì)量約為12 kDa,與膠原域B分子質(zhì)量的1.4倍相符,可以推測分泌表達(dá)出的膠原蛋白具有三股螺旋結(jié)構(gòu)。對膠原域B進(jìn)行除鹽凍干處理,稱重估算產(chǎn)量約為30 mg/L,是優(yōu)化前的3倍。為了進(jìn)一步確定膠原域B的二級結(jié)構(gòu),進(jìn)行了圓二色譜鑒定,如圖7-b所示,膠原域B在225 nm處有膠原蛋白的特征正峰,表明其正確折疊形成三股螺旋結(jié)構(gòu)。又利用圓二色譜在10~70 ℃掃描其熱變曲線,如圖7-c所示,進(jìn)行擬合后顯示膠原域B的Tm值為25.82 ℃,與文獻(xiàn)報道相近[30]。因此,通過信號肽的優(yōu)化,構(gòu)建的谷氨酸棒桿菌表達(dá)體系可以直接分泌表達(dá)具有三股螺旋結(jié)構(gòu)的膠原蛋白。
PENG等[7]在大腸桿菌中表達(dá)來源于化膿性鏈球菌的全長Scl2膠原蛋白V-CL,在搖瓶水平最高產(chǎn)量為300 mg/L,發(fā)酵罐水平最高產(chǎn)量達(dá)19 g/L。本研究中膠原域B的表達(dá)量約為30 mg/L,忽略酶切、除鹽、濃縮的損失以及序列長度對蛋白表達(dá)的影響,根據(jù)分子質(zhì)量估算酶切前重組膠原蛋白V-B的保守產(chǎn)量約為67.3 mg/L,而V-B的分子質(zhì)量約為膠原蛋白V-CL的1/2,由此推算V-B在谷氨酸棒桿菌中的產(chǎn)量約為大腸桿菌中的1/2,且本研究中目的基因尚未進(jìn)行針對宿主的密碼子優(yōu)化,因此推測谷氨酸棒桿菌在分泌表達(dá)膠原蛋白上仍有較大潛力。
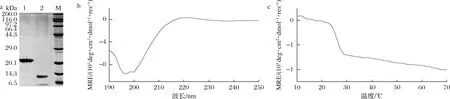
a-SDS-PAGE圖(泳道1為純化后的V-B,泳道2為酶切后樣品);b-膠原域全波長圖譜;c-膠原域熱變曲線圖7 重組膠原蛋白V-B純化、酶切后的SDS-PAGE圖及膠原域B的圓二色譜圖Fig.7 SDS-PAGE after purification and digestion of V-B and circular dichroism of B domain
3 結(jié)論
本研究以食品安全級菌株谷氨酸棒桿菌為底盤細(xì)胞,通過引入不同種類信號肽介導(dǎo)膠原蛋白的分泌表達(dá),結(jié)果顯示Sec依賴型信號肽PorB介導(dǎo)分泌膠原蛋白的產(chǎn)量最高,經(jīng)發(fā)酵條件優(yōu)化,膠原蛋白的產(chǎn)量提高到優(yōu)化前的3倍,經(jīng)圓二色譜表征,證明其膠原域正確折疊形成三股螺旋結(jié)構(gòu),同時發(fā)現(xiàn)溶氧是搖瓶水平影響膠原蛋白產(chǎn)量的關(guān)鍵因素,為工業(yè)生產(chǎn)水平的進(jìn)一步放大試驗提供了指導(dǎo)。本研究是膠原蛋白在谷氨酸棒桿菌中表達(dá)的首次嘗試,后續(xù)可通過密碼子優(yōu)化、表達(dá)元件和載體結(jié)構(gòu)等方面的進(jìn)一步優(yōu)化提高膠原蛋白的表達(dá)量,并且,溶氧作為好氧發(fā)酵過程中的重要參數(shù),通過在發(fā)酵罐水平對溶氧的嚴(yán)格控制有望大幅度提高膠原蛋白在谷氨酸棒桿菌中的產(chǎn)量。本研究中谷氨酸棒桿菌分泌表達(dá)膠原蛋白體系的建立,為膠原蛋白的重組表達(dá)提供了新的宿主選擇,也為微生物難以分泌具有三股螺旋結(jié)構(gòu)膠原蛋白的困境提供了解決思路和理論依據(jù)。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
