油莎豆SRAP-PCR體系優(yōu)化及遺傳多樣性分析
2022-09-19 04:28:48趙琦琦郭玉靜于夢斐王穎高文偉張斌
山東農(nóng)業(yè)科學(xué) 2022年8期
趙琦琦郭玉靜于夢斐王穎高文偉張斌
(1.新疆農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,新疆 烏魯木齊 830000;2.山東省農(nóng)業(yè)科學(xué)院農(nóng)作物種質(zhì)資源研究所,山東 濟南 250100;3.湖南大學(xué)研究生院隆平分院,湖南 長沙 410125;4.曲阜師范大學(xué)生命科學(xué)學(xué)院,山東 曲阜 273165)
油莎豆(Cyperus esculentus)是莎草科莎草屬的禾本科作物,原產(chǎn)于非洲及地中海沿岸的干旱和半干旱地區(qū)[1,2],現(xiàn)廣泛分布于我國的東北、華北及長江流域等中、低緯度地區(qū),性喜溫暖濕潤氣候,具有生長速度快、生物量大、抗逆性強等優(yōu)點[3],耐旱、耐澇、耐貧瘠、耐鹽堿[4]。油莎豆塊莖含油量達27%左右[5],且含有相當(dāng)豐富的膳食纖維、礦物質(zhì)(如鉀、磷、鈣、鎂、鋅、銅)、維生素C和E以及人體必需脂肪酸(如肉豆蔻酸、油酸和亞油酸)等[6]。目前,大多數(shù)油莎豆研究主要集中在種植技術(shù)[7]、油脂提取工藝[8,9]、營養(yǎng)成分[10]及藥用價值分析[11]等方面,對于其種質(zhì)資源遺傳多樣性的研究相對較少。
SRAP又稱相關(guān)序列擴增多態(tài)性(sequencerelated amplified polymorphism,SRAP),由美國加州大學(xué)作物系Li和Quiros于2001年提出[12],是一種無需任何序列信息即可直接PCR擴增的新型分子標(biāo)記技術(shù),具有簡便、高共顯性、易分離條帶及測序等優(yōu)點,已被廣泛應(yīng)用于生物遺傳多樣性分析、遺傳圖譜構(gòu)建、比較基因組及品種鑒定等研究[13,14]。目前,SRAP分子標(biāo)記技術(shù)在紫花苜蓿[15]、馬鈴薯[16]、紫錐菊[17]、草果菊[18]、石榴[19]等多種生物的遺傳多樣性及親緣關(guān)系分析中取得重大進展,但在探究油莎豆種質(zhì)資源遺傳多樣性和親緣關(guān)系上的應(yīng)用卻很少。
本研究利用SRAP分子標(biāo)記技術(shù),采用單因素試驗與正交試驗相結(jié)合的方法,基于引物濃度、混合酶體積、模板DNA濃度三因素,探究了SRAPPCR的最佳擴增體系,并對收集到的16份油莎豆種質(zhì)資源進行遺傳多樣性分析,以探明不同地區(qū)油莎豆種質(zhì)資源的遺傳距離和親緣關(guān)系,以期為油莎豆的種質(zhì)資源鑒定和遺傳育種提供依據(jù)。
1 材料與方法
1.1 試驗材料
供試16份油莎豆種質(zhì)資源材料的來源及特性見表1。于2021年4月收集種子,5月底浸種3~5 d,6月初種植于山東省農(nóng)業(yè)科學(xué)院核心區(qū)溫室大棚內(nèi)底徑26 cm、高23 cm、口徑34 cm的花盆中,盆土為沙質(zhì)土。每盆播種3穴,每穴1粒種子,穴深4~5 cm。播種后及時澆水,保持土壤濕潤,便于種子萌發(fā)。播種后10 d左右出芽,20 d左右采集油莎豆幼嫩葉片,保存于-80℃超低溫冰箱,用于提取DNA。7~12月進行油莎豆SRAPPCR體系優(yōu)化試驗及相關(guān)遺傳多樣性分析。所用引物見表2。

表1 供試16份油莎豆材料的特性及來源
1.2 油莎豆基因組DNA提取與檢測
采用改良的CTAB法提取油莎豆幼嫩葉片基因組DNA,通過微量核酸蛋白濃度測定儀測定DNA濃度。將5μL DNA樣品與1μL 6×Loading buffer混合均勻后點入1%瓊脂糖凝膠,置于100 V電壓下電泳20 min,檢測DNA質(zhì)量。
1.3 SRAP-PCR擴增體系建立與優(yōu)化
選用條帶清晰且多態(tài)性高的Me2+Em2引物對,以2號油莎豆種質(zhì)材料DNA為模板,采用單因素試驗與正交試驗相結(jié)合的方法確定SRAPPCR最佳擴增體系。
SRAP-PCR擴增初始體系(15μL):模板DNA 60 ng,2×PCR Master Mix 7.5μL,正反向引物各0.3μL,ddH2O補足至15μL。SRAP-PCR程序:94℃預(yù)變性5 min;94℃1 min,35℃1 min,72℃1.5 min,10個循環(huán);94℃1 min,50℃1 min,72℃1.5 min,35個循環(huán);72℃延伸10 min。用2%瓊脂糖凝膠檢測PCR產(chǎn)物。
1.3.1 單因素試驗設(shè)計 影響SRAP-PCR擴增體系的主要因素有引物濃度、混合酶體積、模板DNA含量,所以,基于初始擴增體系,對這3個因素分別設(shè)計6個濃度梯度(表2)進行單因素試驗,以確定各因素的基礎(chǔ)濃度范圍。
1.3.2 正交試驗設(shè)計 在單因素所選濃度范圍基礎(chǔ)上設(shè)計L9(33)正交試驗,共9個處理組合,如表3所示。利用陳璨等[20]的方法,對正交試驗結(jié)果進行直觀分析,即:依據(jù)擴增條帶的清晰度和穩(wěn)定性對9個組合的PCR擴增結(jié)果進行評分,條帶數(shù)量最多、清晰度最高的產(chǎn)物記16分,最差的產(chǎn)物記1分;根據(jù)各處理組合評分結(jié)果,計算各因素各水平的得分均值ki,進而計算該因素的極差R=kimax-kimin。

表3 油莎豆SRAP-PCR擴增體系的單因素試驗設(shè)計

表4 油莎豆SRAP-PCR擴增體系的正交試驗設(shè)計
1.4 供試16個油莎豆種質(zhì)材料的SRAP-PCR遺傳多樣性分析
遺傳多樣性分析所用SRAP引物由上海生工生物工程有限公司合成,設(shè)計正向引物6條、反向引物17條(表2),組合成102對引物。利用上述試驗確定的最佳擴增體系對16份油莎豆材料進行擴增,采用6%非變性聚丙烯酰胺凝膠對PCR產(chǎn)物進行電泳,恒壓200 V,電泳1.0~1.5 h,用銀染法顯影,拍照記錄。
利用人工讀帶的方法統(tǒng)計PCR產(chǎn)物中相同位置清晰的條帶數(shù),有條帶的記為“1”,無條帶的記為“0”,轉(zhuǎn)換為“0/1”數(shù)字矩陣。利用NTSYS 2.1計算遺傳一致度和遺傳距離,采用UPGMA法繪制聚類圖,進而分析16份油莎豆材料的親緣關(guān)系。
2 結(jié)果與分析
2.1 DNA提取質(zhì)量檢測結(jié)果
經(jīng)過檢測,提取的DNA濃度范圍在141.5~269.7 ng/μL,OD260/OD280值 在1.8以 上,OD260/OD230值在1.96~2.18之間;DNA條帶清晰完整,無降解和拖尾現(xiàn)象(圖1)。綜上,利用改良CTAB法提取的油莎豆DNA質(zhì)量較好,濃度較高,可用于SRAP-PCR反應(yīng)。

圖1 16份油莎豆材料的DNA電泳檢測圖譜
2.2 SRAP-PCR擴增體系的優(yōu)化
2.2.1 單因素試驗結(jié)果分析 由圖2可知,隨著引物濃度提升,條帶逐漸清晰且多態(tài)性增加,最終根據(jù)條帶的擴增情況選擇0.4、0.5、0.6μmol/L三個引物濃度用于正交試驗。混合酶體積在3.0μL時無擴增條帶,在4.5、6.0μL時擴增條帶較模糊,在7.5、9.0、10.5μL時條帶擴增情況最好,所以選擇7.5、9.0、10.5μL用于正交試驗。DNA模板用量為20 ng時擴增條帶較模糊,其他濃度時擴增情況均較好,選擇60、80、100 ng用于正交試驗。
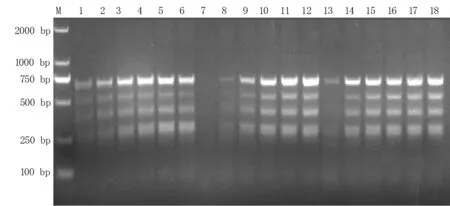
圖2 SRAP-PCR擴增體系單因素試驗結(jié)果
2.2.2 油莎豆SRAP-PCR最佳擴增體系的確立
從圖3可看出,處理組合1、7沒有擴增出條帶,處理組合4、8擴增條帶多態(tài)性較少,其余組合擴增條帶較好但清晰度存在差異。正交試驗直觀分析結(jié)果(表5)顯示,3個因素對SRAP-PCR反應(yīng)體系的影響為混合酶體積>模板DNA含量>引物濃度,引物濃度為0.5μmol/L、混合酶體積為10.5 μmol/L、模板DNA含量為80 ng時擴增效果最佳。以此組合作為最佳反應(yīng)體系,選用不同引物組合,用4號、5號油莎豆材料進行驗證試驗,結(jié)果(圖4)表明,不同引物組合在兩種質(zhì)中均能穩(wěn)定擴增,說明所選體系穩(wěn)定可靠,適用于油莎豆SRAP分子標(biāo)記研究。
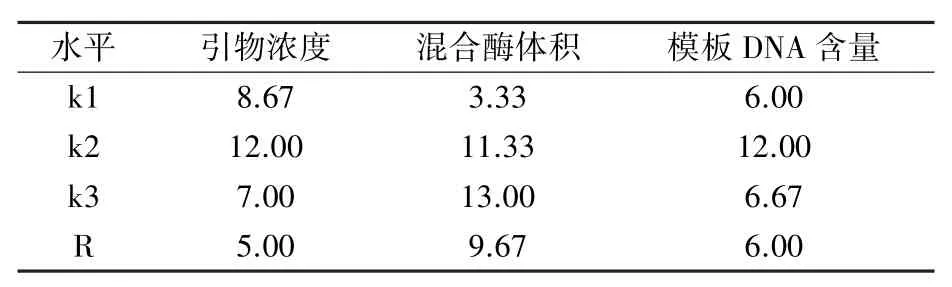
表5 正交試驗直觀分析結(jié)果

圖3 SRAP-PCR擴增體系的正交試驗結(jié)果
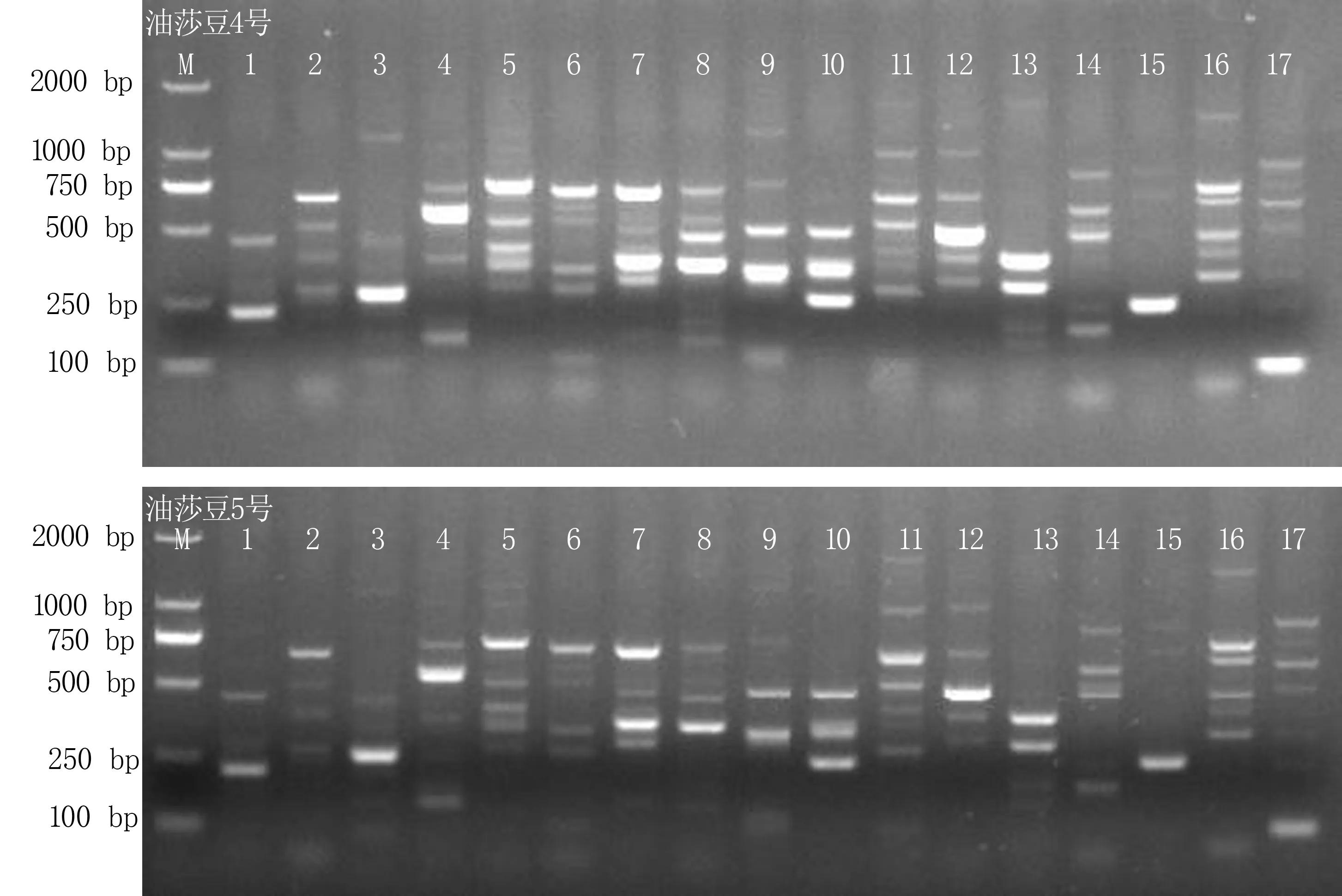
圖4 SRAP-PCR最佳擴增體系在不同品種不同引物穩(wěn)定擴增結(jié)果
2.3 16份油莎豆種質(zhì)材料的遺傳多樣性分析
2.3.1 SRAP-PCR多態(tài)性分析 利用6%非變性聚丙烯酰胺凝膠電泳從102對引物組合中篩選出30對條帶清晰且多態(tài)性較好的引物組合(表6),用這30對引物組合共擴增出312個位點,其中多態(tài)性位點289個,平均多態(tài)性比率達92.82%;每對引物擴增出的總位點數(shù)在6~17個之間,平均10.40個位點;多態(tài)性位點有5~16個,平均9.63個位點。表明篩選出的30對SRAP分子標(biāo)記在16份油莎豆材料中所擴增出的條帶都具有較好的多態(tài)性,可用于遺傳多樣性分析。圖5是引物Me1+Em7和Me2+Em1對16份油莎豆材料的SRAP擴增結(jié)果。
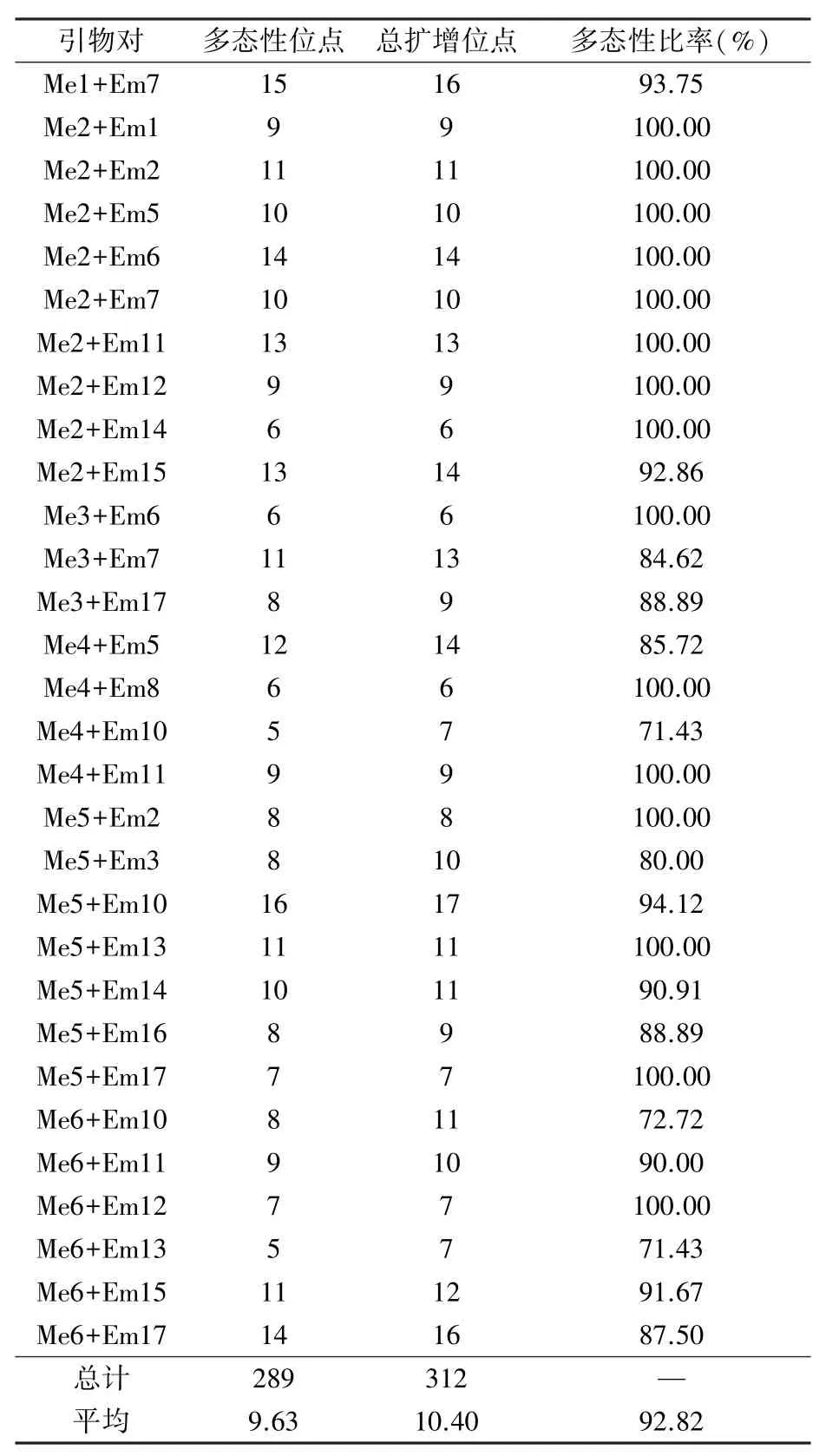
表6 多態(tài)性引物組合擴增結(jié)果

圖5 Mel+Em7和Me2+Eml對16份油莎豆種質(zhì)材料的擴增結(jié)果
2.3.2 16份油莎豆種質(zhì)材料的SRAP-PCR遺傳距離與遺傳一致度分析 由表7可知,遺傳一致度范圍為0.60~0.93,遺傳距離范圍為0.07~0.40。8號與11號種質(zhì)的遺傳一致度最低,親緣關(guān)系最遠,遺傳距離最大;來自新疆維吾爾自治區(qū)的14號與來自河北省的15號材料遺傳一致度最高,親緣關(guān)系最近,遺傳距離最小。由于14、15號材料的品種特性都為黃色圓粒,結(jié)合遺傳一致度結(jié)果,推測它們可能為同一品種。綜合分析油莎豆材料的遺傳距離和遺傳一致度,16份油莎豆種質(zhì)材料間遺傳一致度較高,遺傳背景較狹窄,遺傳差異較小,這可能是因為我國的油莎豆種質(zhì)是由國外引種而來,引種后地域間種質(zhì)交流較頻繁,從而造成了油莎豆種質(zhì)資源具有較高的遺傳一致度。

表7 16份油莎豆材料遺傳距離與遺傳一致度
2.3.3 16份油莎豆種質(zhì)材料的SRAP-PCR聚類分析 利用UPGMA法對16份油莎豆種質(zhì)材料進行聚類分析,得到樹狀聚類圖見圖6,可見聚類結(jié)果與遺傳一致度和遺傳距離的結(jié)果一致。在遺傳相似系數(shù)0.824處,可將16份油莎豆種質(zhì)材料劃分為4個類群,第一、第二、第四類群均包含1份油莎豆種質(zhì)材料,第三類群包含13份種質(zhì)材料。在遺傳相似系數(shù)0.872處,可進一步將第三類群劃分為5個亞類,其中第1亞類包含河南省的3份油莎豆材料,大部分為圓粒;第2亞類包含吉林省的3份油莎豆材料,均為長粒;第5亞類包含其他4個地區(qū)的5份油莎豆材料,絕大部分為圓粒;來自吉林省的長粒油莎豆7號和來自云南省的長粒油莎豆11號材料單獨歸于一個亞類。表明,油莎豆種質(zhì)資源的地域來源和品種特性均對聚類結(jié)果產(chǎn)生影響。
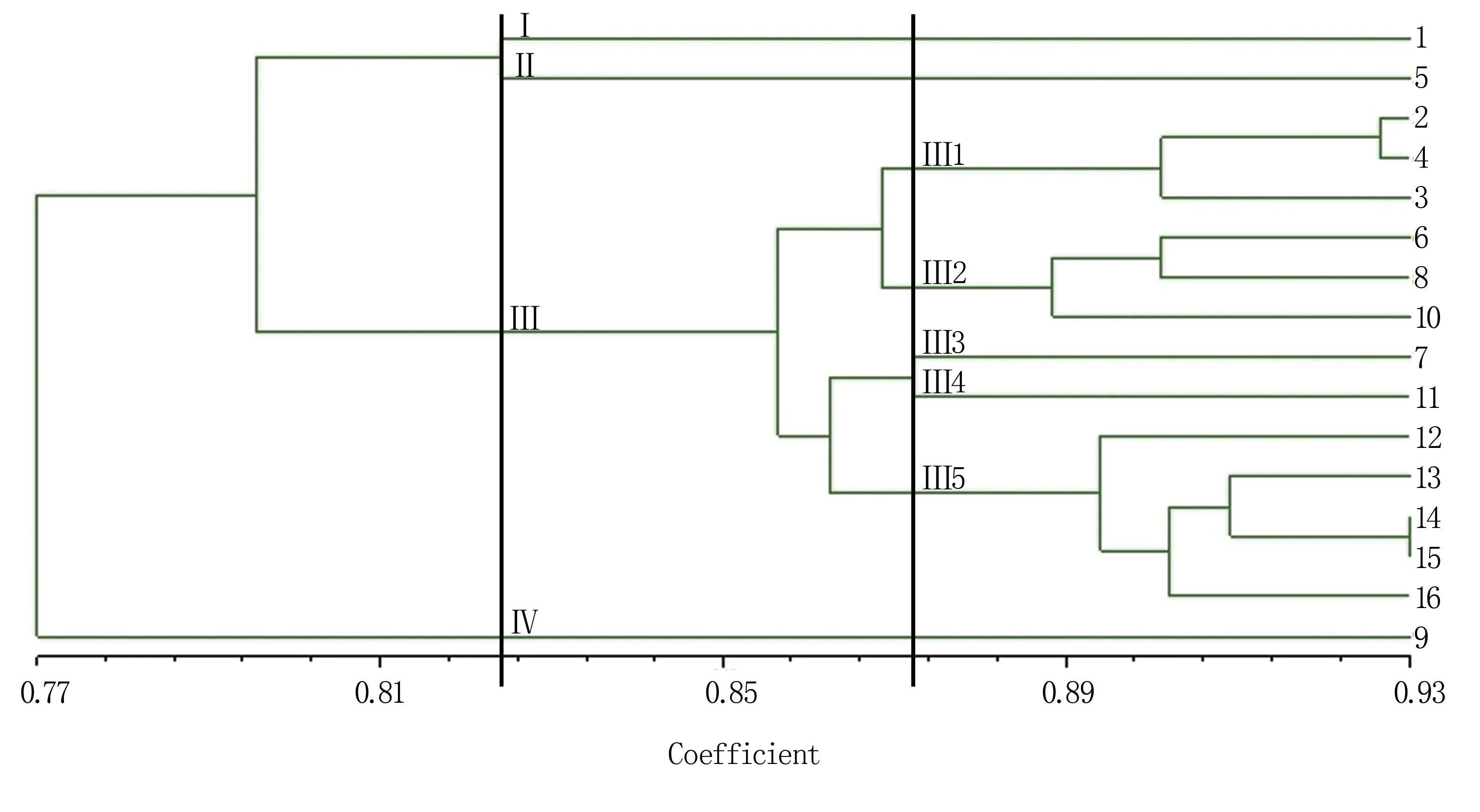
圖6 16份油莎豆種質(zhì)材料的UPGMA聚類結(jié)果
3 討論
3.1 油莎豆SRAP-PCR體系的優(yōu)化
利用PCR擴增體系獲得條帶清晰、多態(tài)性高的引物組合是研究SRAP分子標(biāo)記的基礎(chǔ)。本研究利用單因素試驗和正交試驗確定了適合油莎豆的SRAP-PCR最佳擴增體系,即引物為0.5μmol/L,混合酶體積為10.5μL,模板DNA含量為80ng,用ddH2O補足15μL。這與趙永國等[21]確定的油莎豆SRAP優(yōu)化體系(15μL擴增體系包括DNA模板25 ng、Mg2+1.5 mmol/L、引物濃度1 μmol/L、dNTPs 0.3 mmol/L和Taq酶1 U)不一致,可能與DNA提取方法不同有關(guān),另外,不同廠家的試劑、不同的實驗操作方法也會對SRAPPCR的擴增結(jié)果產(chǎn)生不同的影響。
本研究確定了3個因素對油莎豆SRAP-PCR擴增體系的影響大小為混合酶體積>模板DNA含量>引物濃度。但不同植物中SRAP-PCR擴增體系各因素產(chǎn)生的影響不同:戚華沙等[22]在海南油茶SRAP-PCR擴增體系優(yōu)化中發(fā)現(xiàn),各因素影響大小為引物濃度>dNTPs濃度>TaqDNA聚合酶用量>模板DNA含量;嚴莉等[23]對山桐子SRAP-PCR擴增體系的優(yōu)化結(jié)果表明各因素的影響大小為TaqDNA聚合酶用量>Mg2+濃度>引物濃度>模板DNA濃度>dNTPs濃度。
3.2 SRAP分子標(biāo)記探究油莎豆遺傳多樣性
SRAP分子標(biāo)記使用通用引物進行擴增,常被用于分析遺傳多樣性和親緣關(guān)系,如沈秀芬等[24]利用SRAP標(biāo)記分析草菇遺傳多樣性,發(fā)現(xiàn)草菇種質(zhì)資源在一定程度上出現(xiàn)了地域基因分化;Deng等[25]利用SRAP標(biāo)記首次對中國特有的10種莪術(shù)藥材進行了親緣關(guān)系分析。這些研究表明,SRAP分子標(biāo)記技術(shù)可有效區(qū)分種質(zhì)資源間的遺傳差異,明確其親緣關(guān)系。本研究對收集到的16份種質(zhì)資源進行遺傳多樣性分析,篩選到30對多態(tài)性較好的引物組合,并用其擴增出312個位點,平均多態(tài)性比率為92.82%,說明16份油莎豆種質(zhì)材料具有豐富的多態(tài)性。聚類分析結(jié)果表明,16份油莎豆種質(zhì)材料的遺傳差異與品種特性及來源均有關(guān)。
4 結(jié)論
本研究基于引物濃度、混合酶體積和DNA模板含量3個因素,通過單因素試驗和正交試驗,確立了油莎豆SRAP-PCR的最佳擴增體系為:引物0.5μmol/L,混合酶體積10.5μL,模板DNA含量80 ng,用ddH2O補足15μL。利用該體系從102對引物組合中篩選出30對多態(tài)性引物,并用于對16份油莎豆種質(zhì)材料的遺傳多樣性進行分析,發(fā)現(xiàn)油莎豆種質(zhì)材料間的遺傳多樣性受品種特性和地域來源的雙重影響。本研究結(jié)果可為油莎豆種質(zhì)資源的品種鑒定和選育等提供一定的參考依據(jù)。
