工廠化養殖美洲鰣幼魚的生長特性及消化酶活性變化
2022-09-24 03:23:30稅春施永海陸根海徐嘉波
水產科技情報 2022年5期
關鍵詞:生長
稅春 施永海 陸根海 徐嘉波
(上海市水產研究所,上海市水產技術推廣站,上海 200433)
美洲鰣學名美洲西鯡(Alosasapidissima),隸屬于鯡形目(Clupeiformes)、鯡科(Clupeidae)、西鯡屬(Alosa),是典型的溯河產卵魚類。美洲鰣肉質細嫩,且富含必需氨基酸、不飽和脂肪酸、礦物質等營養物質[1],因而深受人們的青睞,具有廣闊的消費市場。因美洲鰣與我國瀕臨滅絕的中國鰣(Tenualosareevesii)在形態、口感、營養價值等方面都極為相似,我國從20世紀90年代開始引進該魚以彌補日漸稀缺的鰣魚資源。經過十幾年的繁育攻關研究,我國科研工作者在美洲鰣的人工繁殖[2]、早期發育[3]、苗種培育[4-6]、攝食特性[7]、營養與生理[8-10]等方面取得了較大成果,我國的美洲鰣繁育養殖技術日益完善和成熟。魚類新陳代謝的生理機能主要體現在對食物的消化吸收方面,機體消化吸收能力的大小與消化酶的活性關系緊密[11]。研究魚類消化酶的特性對研究其代謝機能和食性特征具有積極意義。
魚類的消化酶是由消化腺和消化管分泌的能催化食物分解的酶。蛋白酶、脂肪酶和淀粉酶是魚類消化生理研究中主要的種類,通過它們的催化分解作用使機體能吸收利用食物中的營養物質。魚類消化酶活力的影響因素可以分為兩類:一類為外源性因素,如溫度、鹽度、pH、食物等;另一類為內源性因素,包括魚類自身的食性和各生長階段的變化。在魚類的發育過程中,會經歷由內源性營養向外源性營養的轉變和從仔魚到稚魚的轉變[12],伴隨機體生長發育階段的改變,消化酶活力也將發生變化。本研究通過對工廠化養殖的美洲鰣幼魚的生長過程進行連續跟蹤,檢測其生長性狀和消化酶(蛋白酶、脂肪酶、淀粉酶)活力的變化,探討美洲鰣幼魚在生長發育過程中的消化特性,以期為美洲鰣消化生理和飼料科學的研究提供參考。
1 材料和方法
1.1 試驗材料和設施
試驗用美洲鰣幼魚來自上海市水產研究所(上海市水產技術推廣站)苗種技術中心。美洲鰣受精卵經孵化后獲得初孵仔魚,并于室內育苗池培育至能攝食商品飼料(“明輝”海水魚膨化料:粗蛋白質40%,粗脂肪6%),然后轉入工廠化養殖池。養殖池為方形水泥池,面積約200 m2,水深1.5 m。養殖池底呈一定坡度,西高東低,進水口設置在養殖池西端,沿池壁切線方向進水;排水口則設置于養殖池東端。為更有利于排水,排水口處略低于四周,呈鍋底狀。排水閥置于養殖池外,池內鋪設PVC管控制排水口,并設有溢水口。池內每2~3 m2配備1個充氣石,24 h持續增氧,以保證池水溶解氧高于5.0 mg/L。養殖用水為當地自然海水,鹽度5~15。
1.2 幼魚的投放和養殖管理
放魚前1周,用高錳酸鉀溶液(質量濃度4 mg/L)進行全池消毒。于2015年7月30日將體長為(54.04±6.07)mm、體質量為(2.03±0.75)g的4 000尾美洲鰣幼魚放入養殖池中。養殖試驗于2015年10月22日結束,共計84 d。養殖期間每周換水1次,換水量約為全池水量的2/3。每2周進行1次池底和池壁的清潔,清洗操作參照施永海等[13]的方法,以減少對幼魚造成的應激。每天8:00、14:00用商品膨化料進行投喂,投飼量以2 h內攝食完為準。每天早晚用表層水溫計測量水溫并做好記錄。
1.3 樣品采集
設定放養日期(7月30日)為養殖開始日期(0 d)。試驗開始后,每14 d采樣1次,共連續采樣7次,采樣前24 h停止投飼。每次采樣時,隨機從池中取10~20尾魚,并重復取樣3次,分別測定試驗魚的體質量和體長。然后將魚置于冰水中麻醉,再分別采集魚體的胃、幽門盲囊和腸道組織,去除內容物,經生理鹽水清洗和濾紙擦干后放入預冷的離心管中,于-20 ℃冰箱中保存,備測。
1.4 指標檢測
檢測前將組織樣品于4 ℃下解凍。先將樣本組織剪碎,混勻,再稱取0.2 g左右樣本,加入9倍體積的生理鹽水,冰浴勻漿,勻漿液在4 ℃、3 500 r/min下離心10 min,取上清液進行消化酶活力的檢測。蛋白酶、脂肪酶和淀粉酶活力均采用南京建成生物工程研究所的試劑盒,按照試劑盒說明書進行測定。相關計算公式如下。
肥滿度=W/L3×100
(1)
成活率(%)=100×(NT+N)/N0
(2)
飼料系數=消耗的飼料總質量/幼魚總質量增加量
(3)
體長和體質量關系式:W=aLb
(4)
體長與養殖時間的關系式:L=aebt
(5)
體質量與養殖時長的關系式:W=aebt
(6)
式(1)~(6)中:W為試驗魚體質量(g),L為試驗魚體長(cm),NT為結束時幼魚的數量(尾),N為幼魚取樣數量(尾),N0為初始幼魚數量(尾),t為養殖時長(d),a、b為常數。
1.5 數據處理
試驗數據采用EXCEL 2003軟件統計后,用SPSS 20.0軟件進行單因素方差分析(one-way ANOVA),如果差異顯著則進行Duncan’s多重比較,設顯著水平為0.05。用SPSS 20.0軟件對美洲鰣體長與體質量的相關性及其分別與養殖時長的關系進行曲線回歸分析和最佳函數關系模擬。利用EXCEL 2003軟件繪圖。描述性統計值采用“平均值±標準差”的方式表示。
2 結果
2.1 美洲鰣幼魚的生長性能
養殖期間水體水溫變化情況如圖1。經過84 d的養殖,美洲鰣幼魚養殖成活率為86%,飼料系數為1.84;幼魚平均體長由(54.04±6.07)mm增至(115.49±7.29)mm,平均體質量由(2.03±0.75)g增至(22.72±4.20)g,平均體長和體質量分別增長了1.14倍和10.19倍。經曲線回歸分析得到體長和體質量的冪函數相關關系式:W=7×10-6L3.158 2,R2=0.991,P<0.01(見圖2)。幼魚體質量(W)和體長(L)均與養殖時長(t)呈指數相關:W=2.463 3e0.027 4t,R2=0.968,P<0.01;L=56.568e0.008 8t,R2=0.981,P<0.01。養殖14 d后,美洲鰣幼魚的肥滿度較初始時顯著提高(P<0.05);養殖28 d時,幼魚肥滿度有所下降,但與14 d時無顯著差異(P>0.05);之后,美洲鰣幼魚的肥滿度顯著下降,養殖42 d時肥滿度與養殖14 d和28 d時相比顯著降低(P<0.05),但隨后幼魚的肥滿度保持著較快的增長速度,養殖56 d、70 d和84 d時的肥滿度均顯著高于養殖42 d時和初始的水平(P<0.05);肥滿度最大值出現在第84天,即試驗結束時(見圖3)。
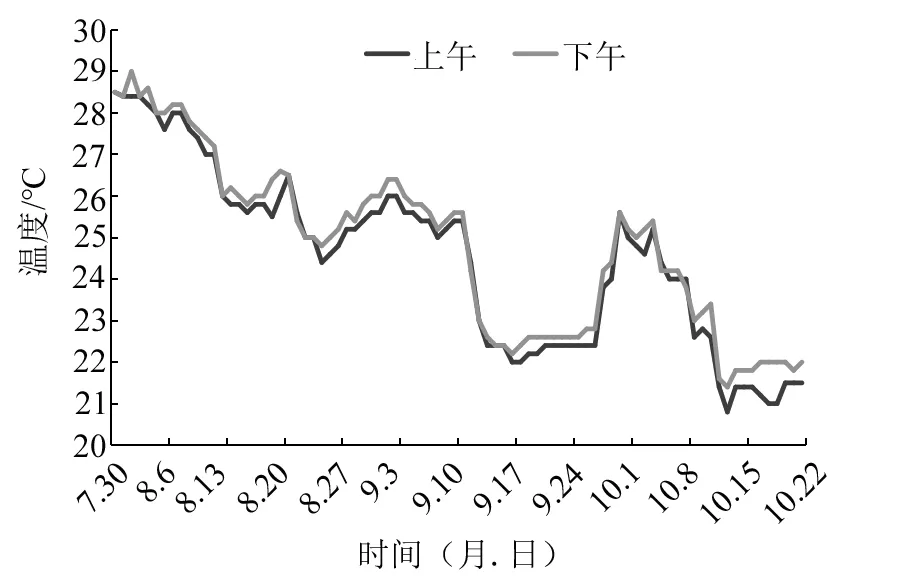
圖1 養殖期間水體溫度變化情況
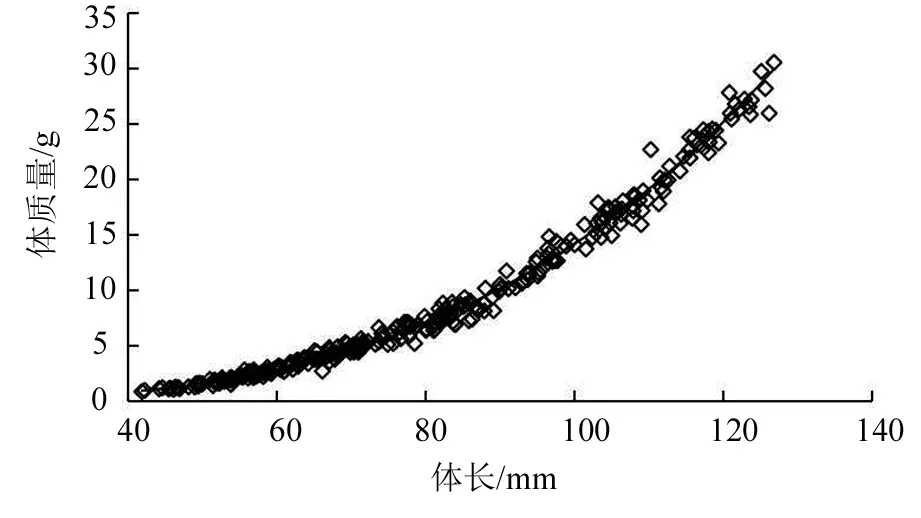
圖2 美洲鰣幼魚體長與體質量的關系

注:圖中有相同字母表示組間差異不顯著(P>0.05)。圖3 美洲鰣幼魚肥滿度隨養殖時長的變化
2.2 美洲鰣幼魚消化酶活力的變化情況
2.2.1 胃蛋白酶
在美洲鰣幼魚生長過程中,其胃蛋白酶活力隨著機體的生長而逐漸升高(見圖4)。養殖14 d時,幼魚胃蛋白酶活力較最初時有顯著上升(P<0.05);從養殖14 d到42 d,胃蛋白酶活力處于相對穩定狀態,沒有明顯的升高趨勢(P>0.05);到養殖56 d時,試驗魚的胃蛋白酶活力又有了顯著提高(P<0.05),之后一直保持著上升的趨勢,但養殖70 d時幼魚胃蛋白酶的活力與56 d時相比沒有顯著提高(P>0.05);試驗期間胃蛋白酶活力的最大值出現在第84天。
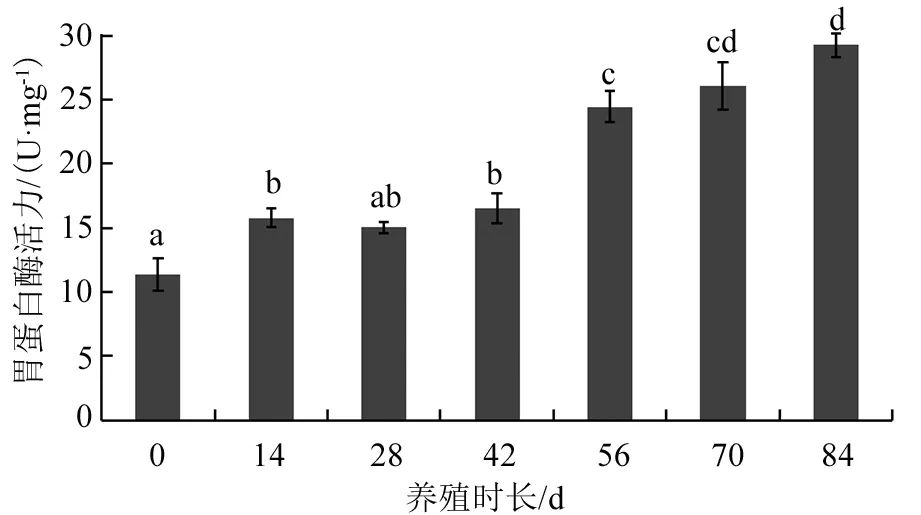
注:圖中有相同字母表示組間差異不顯著(P>0.05)。圖4 美洲鰣幼魚胃蛋白酶的變化
2.2.2 幽門盲囊消化酶
美洲鰣幼魚的幽門盲囊消化酶活力隨養殖時長的變化情況見表1。總體上,淀粉酶活力隨著美洲鰣幼魚的生長呈下降趨勢(P<0.05);從試驗開始至第14天,淀粉酶活力保持著較高水平,第28天時顯著降低(P<0.05);之后保持著相對穩定的狀態,養殖56 d,幼魚淀粉酶活力顯著下降(P<0.05),第70天、第84天時的淀粉酶活力顯著低于初始水平(P<0.05)。蛋白酶活力在第28、42天時顯著低于第84天時(P<0.05),但與其他組無顯著差異(P>0.05)。除第28天和第42天外,其余各組間的蛋白酶活力均無顯著性差異(P>0.05)。養殖開始14 d內幼魚的脂肪酶活力沒有顯著變化(P>0.05),至第28天時顯著下降(P<0.05),第42天與第28天時沒有顯著差異(P>0.05),第56天時的脂肪酶活力較第42天時的顯著提高(P<0.05),并在之后保持顯著上升的趨勢(P<0.05)。
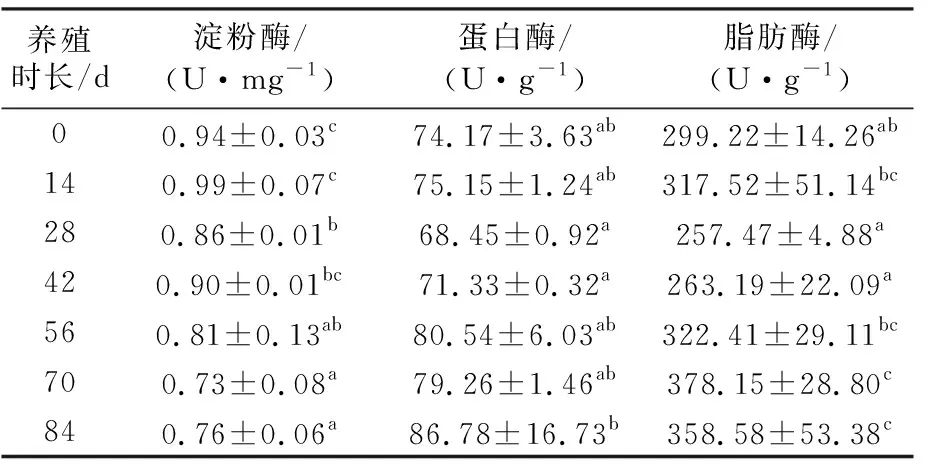
表1 美洲鰣幼魚幽門盲囊消化酶活力隨養殖時長的變化
2.2.3 腸道消化酶
試驗期間美洲鰣幼魚腸道消化酶活力隨時間的變化情況見表2。淀粉酶活力在前56 d內無顯著變化(P>0.05),第70天時腸道淀粉酶的活力較第56天時有所降低,但下降不顯著(P>0.05),直至第84天,即試驗結束時,淀粉酶活力顯著降低(P<0.05)。蛋白酶活力的最大值出現在試驗第84天時,且顯著高于0 d、28 d和56 d的水平(P<0.05),但與70 d、42 d、14 d時的活力水平差異不顯著(P>0.05)。脂肪酶活力的最大值也出現在84 d時,但與42 d、56 d、70 d及初始時的酶活力無顯著差異(P>0.05);試驗第14天和第28天脂肪酶的活力較初始時顯著降低(P<0.05);美洲鰣幼魚的脂肪酶活力在養殖14 d至70 d期間無顯著差異(P>0.05)。
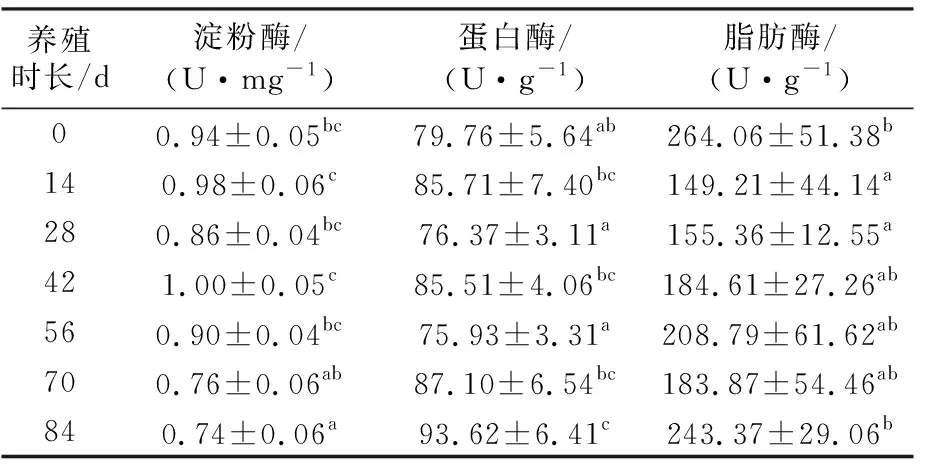
表2 美洲鰣幼魚腸道消化酶活力隨養殖時長的變化
3 討論
3.1 美洲鰣幼魚的生長類型
經過在復雜環境長期的生存選擇,每種魚類都形成了其獨特的、可以遺傳的生長規律。對魚類生長規律最直觀的反映就是體長與體質量的關系。比較生長速度、冪函數方程被廣泛應用于描述魚類的生長特點。冪函數方程通常表述為:W=bLa,參數a表示魚類生長發育的不均勻性,這是由體質量和體長增長的不均勻造成的,所以可通過生長冪指數a來判斷魚類是否處于等速生長:當a=3即為等速生長,a≠3即為異速生長[14]。本研究中,美洲鰣的生長冪函數方程為:W=7×10-6L3.158 2,其中a≈3,表明美洲鰣幼魚的生長為等速生長,且體質量與體長的回歸分析結果為顯著相關關系,其相關系數為0.991。據此推測,目前的養殖條件對于美洲鰣的生長是適宜的,具有積極的影響[15]。
3.2 工廠化養殖方式對美洲鰣幼魚生長的影響
美洲鰣幼魚在工廠化養殖條件下的生長性能除了由其自身遺傳因子所決定外,還會受到溫度、食物等諸多外部環境因素的影響。朱雅珠等[16]用不同蛋白質和脂肪水平的膨化飼料飼喂美洲鰣,結果顯示,當飼料的蛋白質水平在40%~45%,脂肪含量為6%~8%時養殖效果最佳;楊坤等[17]的研究認為,當飼料中的蛋白質含量為40%時養殖效果最佳。參考以上的研究結果,本試驗使用粗蛋白質質量分數為40%的商品膨化飼料,以保證美洲鰣幼魚的正常生長。
肥滿度作為魚類的生長性狀之一,被廣泛用于衡量魚類的營養狀況和豐滿程度[18]。本研究中,美洲鰣幼魚在前28 d內肥滿度顯著增加,但第42天又降至初始水平,之后一直保持著上升趨勢。這樣的變化可能與美洲鰣幼魚在前42 d內淀粉酶活力的逐漸下降及蛋白酶活力的波動有關。從飼料系數看,工廠化養殖美洲鰣幼魚的飼料系數為1.84,較水庫網箱養殖模式(2.5)更低[19],這可能與養殖模式及試驗用魚規格不同有關。
3.3 美洲鰣幼魚生長過程中消化酶活力變化特點
魚類體內淀粉酶的活力與魚類食性相關。研究表明,草食性和雜食性魚類的淀粉酶活力高于肉食性魚類[20]。淀粉酶存在于魚類的各消化器官中,但酶的活力會有所差異。本研究中,幽門盲囊和腸道中的淀粉酶活力整體都不高,且隨著美洲鰣的生長發育呈現顯著降低的趨勢,這是由美洲鰣的動物食性決定的[21]。淀粉酶活力的前后變化屬機體自身調控的結果。劉金蘭等[22]對美洲鰣消化酶的研究發現,肝胰臟是淀粉酶分泌及存儲的主要場所,但真正發揮作用的是幽門盲囊和腸道。鑒于本研究中美洲鰣幽門盲囊和腸道被檢測出的淀粉酶活力較低,建議在養殖過程中可以適當調整飼料中淀粉的含量或添加生物酶制劑,以提高美洲鰣對飼料中淀粉類物質吸收利用的效率。
美洲鰣的胃發達且呈“Y”型,攝入的食物首先到達胃部,在胃蛋白酶的作用下對食物中的蛋白質進行消化。但本研究發現,美洲鰣幼魚胃蛋白酶活力顯著低于幽門盲囊和腸道中蛋白酶的活力。分析認為,美洲鰣對蛋白質的消化吸收場所主要在幽門盲囊和腸道中,其中胃的主要功能可能是短暫容納食物,使胃蛋白酶完成對食物中蛋白質的初步消化,為幽門盲囊和腸道的進一步消化吸收作準備,同時提高對食物蛋白質的吸收。這在對鱖魚消化酶的研究中也得到證實[23]。美洲鰣具有十分發達的幽門盲囊,為食物的充分消化和吸收提供了保障。研究認為,幽門盲囊是腸道的延伸,其在腸道長度和厚度不變的條件下可以起到增加腸道表面積的作用,這是魚類的一種適應性表現[24]。本研究結果顯示,養殖美洲鰣幼魚幽門盲囊蛋白酶始終保持著較高的活力,且隨著機體的生長,蛋白酶活力不斷升高。與幽門盲囊相連的是腸道,美洲鰣的腸道較短,不及其體長的一半[21],因此吸收功能不及幽門盲囊強。但試驗結果顯示,腸道蛋白酶也具有較高的活力,說明美洲鰣腸道對蛋白質依然保持著較高的消化能力,可以繼續消化幽門盲囊未消化的物質,對提高機體的吸收利用率起到了積極的作用。
魚類的脂肪酶主要由肝胰臟合成[25]。研究認為,腸道是魚類脂肪消化吸收的主要場所[26-28]。通過檢測美洲鰣幼魚幽門盲囊和腸道的脂肪酶活力發現,兩處消化器官中的脂肪酶都保持著較高的活力,說明美洲鰣有較強的脂肪消化能力。此外,試驗過程中美洲鰣幼魚腸道和幽門盲囊脂肪酶活力均呈先下降后上升的變化趨勢,這就要求在實際養殖工作中應適時調整飼料的營養成分,或通過人為輔助手段(如添加生物酶制劑),及時滿足美洲鰣的營養需求,提高飼料的利用率。
綜上所述,美洲鰣幼魚在工廠化養殖條件下呈等速生長,胃蛋白酶活力隨著機體生長而提高;幽門盲囊和腸道中的淀粉酶呈顯著降低趨勢,蛋白酶和脂肪酶具有先降低后升高的變化特點。研究結果提示,在養殖過程中,需注意飼料中營養成分的含量,適時降低飼料中的碳水化合物含量,或采取人為輔助措施以提高飼料的利用效率,從而提高養殖經濟效益。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
