醉馬草種子萌發對增溫和降雨變化響應的研究進展
2022-09-29 11:05:38田偉濤張耀丹李超鋒李慶偉徐井潤董乙強吳淑娟袁曉波
草地學報 2022年9期
關鍵詞:研究
田偉濤, 張耀丹, 夏 江, 李超鋒, 李慶偉, 徐井潤, 董乙強, 吳淑娟, 袁曉波*
(1. 新疆昌吉州草原站, 新疆 昌吉 831100; 2. 蘭州大學草地農業科技學院, 甘肅 蘭州 730020; 3. 新疆瑞昶設計院有限公司, 新疆 烏魯木齊 830000; 4. 新疆農業大學草業學院, 新疆 烏魯木齊 830052)
近年來,全球氣候變化已成為各地政府和科研工作者所關注的重點問題之一[1-2]。聯合國政府間氣候變化專門委員會(Intergovernmental Panel on Climate Change,IPCC)第五次氣候變化評估報告中強調,溫度升高和降水變化是當前全球氣候變化最為顯著的驅動因子[3]。據統計,自工業革命以來,人類活動加劇導致全球平均氣溫升高了約1.5℃[4]。與之相比,我國氣候變暖形勢更為嚴峻,過去50年,我國地表溫度增加幅度較大,同期增溫幅度明顯高于全球平均水平,且表現出在未來仍會持續增加的趨勢[5-6]。相比于全球平均氣溫不斷升高的趨勢,降水變化具有較大的不確定性[7]。Folland等[8]研究發現,過去一個世紀全球總降水量呈現出增加的趨勢,并且暴雨和干旱等極端氣候事件的發生頻率亦有所增加。胡婷等[9]通過模型模擬的研究表明全球降水變化總體表現為“濕者愈濕、干者愈干”。總之,以溫度升高和降水變化為主要特征的全球氣候變化趨勢愈發明顯。
種子萌發是植物幼苗生長和群落建植等不可或缺的階段,受溫度和水分等非生物因子調控[10-11]。溫度對于植物種子萌發具有至關重要的作用,其影響植物種子萌發進程的可能原因有以下兩點:一方面,溫度會影響種子內部細胞器和生物膜系統等,進而影響細胞內部正常生命活動的有序進行[12];另一方面,溫度的高低亦影響植物發芽過程中對各種生理生化反應起關鍵作用酶的活性,當溫度過低時,較低的酶活性不足以催化各種代謝反應的有效進行,而溫度過高會導致酶變性失活。因此,溫度對植物種子萌發的影響取決于溫度的高低,并且不同的植物具有適宜其種子萌發的最佳溫度和耐受限度[13-14]。水分也是調控植物種子萌發的重要非生物因子[15-16]。通常,植物種子成熟后會極度脫水,吸漲是種子萌發的第一步,只有吸漲后種子才能啟動萌發過程,而不同植物啟動其種子萌發時對外界水分含量的需求或高或低,這可能是植物躲避惡劣氣候的一種生態對策[17-18]。水分脅迫會促使植物種子內活性氧的產生,高濃度的活性氧可通過氧化蛋白質干擾種子內部正常的細胞新陳代謝活動,從而對植物種子萌發進程產生一定影響[19]。溫度和水分通過影響植物種子萌發特性,進而對田間出苗等農牧業生產活動產生深遠影響,因此研究種子萌發如何響應溫度和水分等非生物因子的變化對于農牧業生產具有重要實踐意義[11,16,20-21]。
醉馬草(Achnatheruminebrians)為多年生冷季型草本植物,屬于禾本科(Gramineae)芨芨草屬(Achnatherum),分布海拔為1 200~4 300 m,尤其以亞高山和高山草原分布較為廣泛,是我國西北地區天然草原常見的一種多年生有毒草本植物[22-25]。眾多研究發現醉馬草種子中內生真菌帶菌率可達100%[26-28],內生真菌侵染致使家畜采食醉馬草后表現出食欲不振、神情呆鈍、蹣跚如醉等中毒癥狀,對畜牧業生產造成巨大經濟損失[25,29]。另外,內生真菌與醉馬草之間可形成互利共生的關系[30],醉馬草可為內生真菌提供生活場所,而內生真菌不僅能夠提高醉馬草對干旱、高低溫、鹽堿、重金屬等逆境的耐受程度,亦能促進醉馬草在逆境條件下種子的萌發[31-35]。目前,關于增溫和降水變化對醉馬草種子萌發影響的基礎性研究已有眾多報道,但對于種子萌發響應增溫和降水變化的作用機理和分子機制尚不明晰,關于禾草內生真菌在調控種子萌發對增溫和降水變化響應過程中扮演何種角色仍具有較大不確定性。鑒于此,本研究針對該領域國內外相關研究進展進行梳理總結并展望,以期為進一步深入開展醉馬草防控相關研究提供基礎資料和理論依據。
1 醉馬草的特性以及國內分布概況
醉馬草是我國天然草地常見的一種有毒草本植物,分布范圍廣泛,主要集中于海拔為1 200~4 300 m的地帶,包括新疆、青海、內蒙古、寧夏、甘肅以及西藏部分地區(表1)[26]。相關研究表明,新疆地區醉馬草的分布面積在1987年為40.0萬公頃,1992年分布面積達到了53.3萬公頃,5年期間分布面積增加了33.25%[36-37],局部地區醉馬草的覆蓋率甚至高達85%[38]。近年來,醉馬草分布面積和覆蓋率在不同地區呈擴增趨勢[39]。例如,近期有研究通過建立生態位模型預測了醉馬草在青海省的潛在擴散區域,結果表明,醉馬草在青海省的潛在可擴散生存區域達到青海省土地面積的50%[40]。
醉馬草分布面積的增加主要與其自身較強的適應性和抗逆性密切相關。首先,醉馬草具有株體高(平均高度約100 cm),葉片質地偏硬,圓錐花序緊密呈穗狀且分蘗數多(約100)等特點[39],其對光能和資源的獲取能力強,具有更強的生態位優勢;其次,醉馬草種子產量高,且種子成熟后易脫落,可附著于放牧家畜表皮并進行傳播,有利于其在草地中定植[34,38,41];此外,眾多研究發現醉馬草種子中內生真菌帶菌率可達100%[26-28],且后續研究已發現2種與醉馬草共生的內生真菌,包括甘肅內生真菌(Epichloёgansuensis)和醉馬草內生真菌(E.inebrians)[27],其可提高醉馬草對干旱、高低溫、鹽堿、重金屬等逆境的耐受程度[23,31-35]。
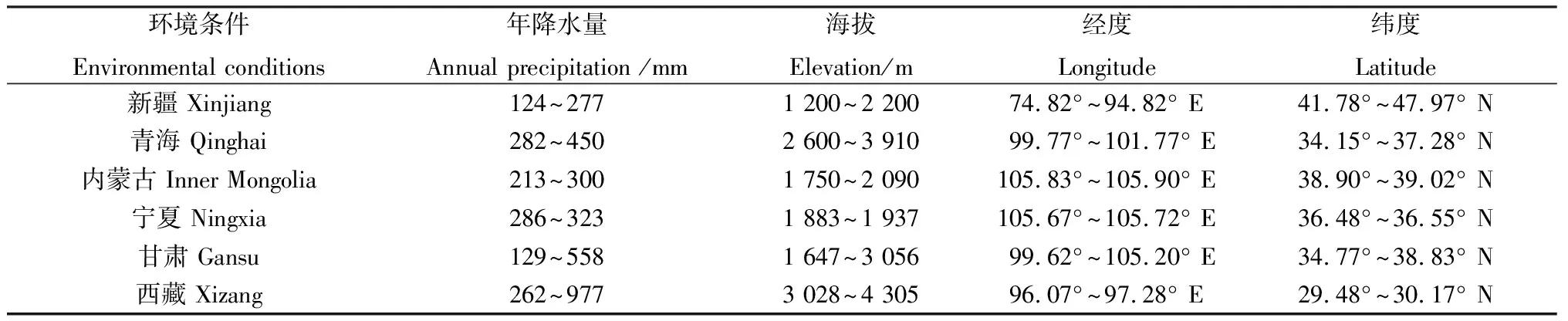
表1 醉馬草在我國的主要分布區及環境條件[26]Table 1 The main distribution areas and environmental conditions of Achnatherum inebrians in China[26]
家畜采食醉馬草后通常表現出食欲不振、神情呆鈍和蹣跚如醉等中毒癥狀[25]。相關研究認為家畜采食醉馬草后表現中毒癥狀與其芒刺刺傷家畜有關[42-43]。然而,有研究表明醉馬草芒刺刺傷家畜多表現為傷口處形成小膿腫或刺傷眼角膜使得家畜失明,因此,物理性芒刺刺傷并非醉馬草致毒的真正原因[44]。此外,相關學者通過旋轉蒸發等方法從醉馬草水提液中分離得到麥角新堿、麥角酰胺及異麥角新堿等多種生物堿,并確定醉馬草大量產堿的原因是香柱菌屬內生真菌的侵染,但這些研究并未進一步深入開展毒性試驗,從而得到生物堿造成家畜中毒的機理[45-48]。后續有研究表明,麥角新堿及異麥角新堿這兩種生物堿顯著抑制了動物體內平滑肌細胞的正常生長,并且明確了麥角新堿及異麥角新堿對平滑肌細胞的半致死濃度[29]。至此,明確了家畜采食醉馬草致其中毒的真正原因是醉馬草與內生真菌共生后產生的生物堿對家畜的直接毒害作用[48-50]。
2 醉馬草種子萌發對增溫的響應
溫度是植物種子萌發的重要調控因子,種子萌發初期需將種胚內所儲存的蛋白質等大分子物質分解為氨基酸等可溶性小分子物質,以提供種子萌發時所需的能量物質[51]。這一過程需要各種酶類參與,酶的種類及含量受外界溫度影響,進而直接影響種子的代謝狀態和能量水平,從而調控種子的萌發[52]。因此,溫度過低或過高均會對酶促反應如蛋白酶將貯藏蛋白質降解成氨基酸的過程產生不利影響,進而抑制種子萌發[13-14]。截至目前,關于溫度對醉馬草種子萌發影響的研究結果已有相關報道,且多采用室內人工氣候箱模擬全球增溫的方法進行研究。例如,一項對烏魯木齊市南山的醉馬草種子進行高溫處理研究發現,隨溫度和處理時間持續,醉馬草種子發芽率、發芽勢、發芽指數和活力指數均表現出先升高后降低的趨勢,且當75℃處理30 d時,醉馬草種子完全不能萌發[53]。上述研究結果表明醉馬草種子萌發對溫度的響應存在最適溫度值或范圍[54]。盡管相關研究學者已對醉馬草種子萌發的最適溫度進行研究,但研究結果并不一致。例如,魚小軍等[23]和Ahmad等[55]的研究結果均發現,醉馬草種子最適萌發溫度為20℃。然而,一些研究結果表明,醉馬草種子在25℃時萌發效果最好[37,56],造成上述研究結果不一致的原因可能是不同研究所使用的試驗材料生境不同。此外,以上研究僅關注醉馬草種子萌發對恒溫的響應,而關于變溫如何影響醉馬草種子的研究較少。岳永寰等[57]的研究發現,醉馬草種子在恒溫15℃~35℃和變溫20/10℃~40/30℃條件下均能較好地進行萌發。這些研究結果充分說明醉馬草種子具有較為寬泛的萌發溫幅,溫度過低或過高均不利于其種子萌發[13-14]。
醉馬草種子萌發對于低溫和高溫脅迫的表現不盡相同。相關研究表明,低溫(5℃)條件下醉馬草種子仍然能夠萌發,而高溫(75℃)條件下醉馬草種子已經失活,不能萌發,這表明醉馬草種子具有更強的耐低溫能力[43,53]。主要因為低溫環境致使醉馬草種子進入休眠狀態進而避開低溫對其萌發產生不利影響;相較而言,溫度過高導致醉馬草種子內部與萌發相關的酶類失活,致使萌發過程中酶促反應不能正常進行,進而使其失活[14-16]。
醉馬草種子萌發對溫度的響應亦受母本環境的調控。例如,一項對高山草原、干旱草原和荒漠草原的醉馬草種子的萌發試驗表明,低溫條件下高山草原的醉馬草種子發芽率顯著高于干旱草原和荒漠草原,而高溫條件下干旱草原和荒漠草原的醉馬草種子發芽率顯著高于高山草原[58]。這可能因為母本長期所生長的環境不同,造成同種植物具有不同的溫度敏感性,從而致使種子萌發對溫度響應的分異性[54,59]。
綜上所述,醉馬草種子具有較為寬泛的萌發溫幅,其在萌發過程中對溫度表現出一定的耐受性和適應性,這也是醉馬草分布面積和覆蓋率擴張的原因之一。全球氣候變暖在一定溫度范圍內會促進醉馬草種子萌發,一旦增溫幅度超過耐受范圍,則會對醉馬草種子萌發產生不可逆轉的損害。
3 醉馬草種子萌發對降水變化的響應
近年來,關于醉馬草種子萌發對降水量變化響應的研究已有眾多報道,多采用PEG 6 000模擬干旱脅迫和降水變化,但研究結論并不一致,包括正效應、中性效應和負效應。例如,一項使用PEG-6 000(聚乙二醇)模擬干旱脅迫對醉馬草種子萌發進程影響的研究發現,低濃度PEG-6 000(5%~10%)可促進醉馬草種子發芽,而高濃度PEG-6 000(15%)則抑制其發芽勢和發芽指數等[60]。另有研究表明,輕微水分脅迫對醉馬草種子萌發無顯著影響[55]。此外,劉雪松[61]研究了采自甘肅榆中和內蒙古阿拉善的醉馬草種子對水分脅迫的響應并發現,水分脅迫會抑制醉馬草種子萌發,且隨著干旱程度加劇,種子萌發受到的抑制作用越強。對于上述研究結果的差異性可歸結于以下幾方面原因:一方面,種子來源不同使得其對于水分脅迫的耐受程度有所差異,致使種子萌發對于水分變化的響應不一致[15];另一方面,不同研究設置的水分梯度不同也會影響試驗結果[16,55,62]。
目前關于長期干旱脅迫去除后醉馬草種子是否具有萌發活力的研究尚未見報道,而短期干旱脅迫進行復水處理后的醉馬草種子萌發狀況已有報道。相關研究表明,短暫干旱脅迫不會導致醉馬草種子完全喪失活性,復水處理后,反而會促進種子萌發[37,55,60]。這說明醉馬草種子具有很強的抗逆性,干旱環境導致醉馬草種子被迫進入休眠狀態,當處于合適的生長條件時停止休眠并開始萌發[16,55]。干旱時種子進入休眠狀態是醉馬草種子逐漸適應干旱環境的萌發策略,能確保其在生活周期的初始競爭中占據一定優勢,從而使醉馬草在天然草地植物群落中占據絕對的生長優勢。
母本環境同樣會影響醉馬草種子萌發對降水變化的響應。位海玲等[60]的研究結果表明,干旱脅迫條件下采自高海拔區域(2 287 m)的醉馬草種子發芽率明顯小于采自低海拔區域(1 650~1 832 m)的種子發芽率。因此,母本生長于中低海拔區域的醉馬草抗旱性高于高海拔區域。
綜上所述,目前關于醉馬草種子萌發對降水量變化響應的研究報道較多,但研究結論并不一致,這可能因為不同研究的水分梯度設置不同。因此,迫切需要建立統一的研究方法以厘清醉馬草種子萌發對降水變化的潛在響應機理和過程。
4 禾草內生真菌調控種子萌發對溫度和降水變化的響應
禾草內生真菌指在宿主禾草體內完成大部分或整個生活史且與宿主禾草形成互惠共生體的一類真菌[30,63-64]。截至目前,已證實在全球300多種禾草中存在內生真菌,并且從中分離鑒定了45個香柱菌屬(Epichloё)內生真菌[65-69]。諸多研究發現我國北方天然草原的醉馬草種子中內生真菌帶菌率可達100%[26-28],且發現2種內生真菌存在于醉馬草體內,分別是甘肅內生真菌和醉馬草內生真菌(表2)[24,70]。眾多研究結果表明,內生真菌能夠增強醉馬草對干旱、高低溫、鹽堿和重金屬等逆境的耐受程度[23,31-35]。此外,醉馬草-內生真菌共生體主要通過種子進行垂直傳播,因此內生真菌的存在勢必對醉馬草種子萌發產生一定的影響[38,69]。

表2 2種與醉馬草共生的內生真菌Table 2 Two endophyte symbiotic with Achnatherum inebrians
目前相關研究表明,高溫、干旱等逆境條件下內生真菌能夠促進宿主植物種子萌發,主要集中在高羊茅(Festucaarundinacea)、多年生黑麥草(Loliumperenne)、多花黑麥草(Loliummultiflorum)和野大麥(Hordeumbrevisubulatum)等草本植物[71-74],而關于內生真菌如何調控醉馬草種子萌發對溫度和降水變化響應研究相對缺乏。例如,不同低溫脅迫對醉馬草種子萌發影響的研究發現內生真菌可以提高醉馬草種子的發芽率和發芽速度,促進胚根和胚芽的生長[43]。與之相似,柳莉等[71]的研究結果亦表明內生真菌能夠顯著增加低溫環境下醉馬草種子萌發能力。內生真菌促進醉馬草種子對低溫的耐受能力可能是以下4方面原因:首先,相比于未攜帶內生真菌的醉馬草種子而言,體內存在內生真菌的醉馬草種子更加的成熟飽滿,種子內部儲存的能量更為充足[73];其次,低溫條件下內生真菌導致參與多個代謝途徑的一系列基因發生了差異表達,進而提高了醉馬草對低溫環境的耐受程度[43];再次,內生真菌菌絲可以突破種皮并為醉馬草種子萌發提供碳水化合物等能量物質[74];最后,內生真菌的次生代謝物中存在醉馬草種子萌發所需的信號物質,例如菌根菌自身所分泌的Strigolactones是種子萌發的重要信號物質之一[43]。
研究表明,內生真菌可以促進干旱條件下醉馬草種子萌發,且干旱脅迫越嚴重這種促進作用愈加明顯[69]。此外,內生真菌對醉馬草種子抗旱性的促進作用亦受其母本環境影響。例如,有研究結果發現,內生真菌對濕潤地區醉馬草種子抗旱性的促進作用強于干旱地區的醉馬草種子[69]。這可能因為長期處于干旱地區,醉馬草-內生真菌共生體已經形成了通過滲透調節和抗氧化保護系統等一系列生理生化反應誘導植物提高其抗旱能力的應對機制,而濕潤地區水分并不是限制醉馬草生長和擴繁的主要因素,使得內生真菌對于醉馬草種子抗旱性的促進作用未得到激發[23]。
5 研究展望
截至目前,關于增溫和降水變化對醉馬草種子萌發影響研究已取得一定進展[10-11,22-25],但缺乏增溫和降水變化對種子萌發影響的作用機理和分子機制研究。同時,禾草內生真菌在調控醉馬草種子萌發對增溫和降水變化響應過程中扮演怎樣的角色仍具有較大的不確定性。基于此,建議今后從以下幾方面進一步開展相關研究:
(1)降水變化不僅表現為降水量的變化,還體現在降水頻度等格局變化方面[8]。在水分影響醉馬草種子萌發的研究中,大多使用PEG-6000進行室內模擬干旱脅迫,關于醉馬草種子萌發表現對降水頻度等格局變化響應的研究尚未見報道。
(2)種子萌發受氧氣、埋藏深度和水熱等多種非生物因子及其交互作用共同調控,然而現有研究多數聚焦于單一環境因子,缺乏多種環境因子及其交互作用對醉馬草種子萌發的影響研究。
(3)醉馬草即可作為一種草原毒害草進行有效防控,亦可作為一種潛在牧草資源和抗逆先鋒種進行深度開發利用,因此,闡明其在全球氣候變化大背景下種群更新機制尤為重要。后續研究應系統闡明全球氣候變化對醉馬草種子萌發、幼苗生長和成苗特性的影響。
(4)應用微生物學、分子生物學和多組學等理論方法,進一步開展內生真菌對醉馬草種子抗旱和耐高低溫的作用機理研究,以期從分子水平深入解析內生真菌對逆境下醉馬草種子萌發的作用機理。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
