兩種生態型大麻雜交F2代主要農藝性狀遺傳變異分析
2022-09-30 07:20:12杜光輝郭鴻彥郭孟璧許艷萍張慶瀅
西南農業學報 2022年7期
張 坤,陳 璇,杜光輝,郭鴻彥,郭 蓉,呂 品,郭孟璧,許艷萍,張慶瀅,楊 明
(1.云南大學農學院/資源植物研究院,昆明 650201;2.云南省農業科學院經濟作物研究所,昆明 650205)
【研究意義】大麻(CannabissativaL.)為大麻科大麻屬一年生草本植物,多雌雄異株[1]。大麻植物學分類還存在一定爭議,有學者認為大麻至少存在indica和sativa兩個亞種或變種(或生態型)[2-4],二者形態差異較大,前者普遍株型矮小,節間較短,分枝緊湊,后者則株型高大,節間較長,分枝稀疏。作為一種傳統的經濟作物,大麻各種用途都與農藝性狀有密切關系,但長期以來由于中國工業大麻品種(指花、葉四氫大麻酚含量<0.3%,無毒品利用價值的大麻品種)選育親本多來自國內種質資源,親本來源范圍小,且大多數屬于sativa生態型,使得工業大麻品種同質化較嚴重,遺傳基礎趨同[5]。此外,由于大麻為短日照植物,易受光照、溫度、土壤等環境因素影響[6-7],導致品種適應范圍有限。因此,育種工作者想通過擴大親本來源,選育出能滿足不同用途和栽培環境的品種。【前人研究進展】株高、莖粗等表型性狀與大麻的生物產量和收獲指數有密切聯系[8],培育具有優良農藝性狀的新品種是提高大麻產量及品質的有效方法之一。長期的生產實踐表明,纖維用大麻植株以節數少、分枝少、株高和莖粗適中及葉少為優[9];而以籽用為主時,良好的株型結構能提高光合利用率[10],進而提高麻籽產量,在一定范圍內,籽粒產量與株高、莖粗、分枝數等農藝性狀存在正相關性[11];以藥用為主時,因藥用成分(大麻素,Cannabinoid)主要存在于雌株花葉中[12-13],花葉產量又與分枝數、株高等農藝性狀有密切關系。湯志成等[14]通過對16份大麻種質資源(其中12份為野生種質資源,4份為栽培品種)表型性狀進行遺傳多樣性分析,結果表明中國野生大麻種質資源表型變異大,遺傳多樣性豐富。呂詠梅等[15]對大麻不同時期7個農藝性狀進行了廣義遺傳力分析,結果表明大麻工藝成熟期株高、莖粗、鮮重等農藝性狀廣義遺傳力較快速生長期要大。Petit等[16]利用123份大麻種質材料開展了與纖維品質有關的28個農藝性狀的遺傳變異研究,結果表明株高、莖粗和花期等農藝性狀在不同生態環境下差異顯著。【本研究切入點】農藝性狀對大麻栽培生產和目標產物的獲得有極大影響。目前對大麻不同亞種雜交后代農藝性狀進行遺傳變異及相關性研究鮮有報道,尚沒有對大麻分離群體在不同生態環境下開展農藝性狀遺傳的研究。盡管獲得優良農藝性狀的方法很多,但雜交育種方法仍是最常見且最有效的方法之一。陳璇等[5]認為在工業大麻品種選育過程中,應擴大品種的遺傳背景,盡可能選擇地理分布較遠,目標性狀差異較大的親本,避免品種同質化現象。【擬解決的關鍵問題】本研究選用兩個農藝性狀差異較大且分屬于不同生態型的大麻材料作為親本,通過化學誘導構建F2代全雌性株系群體,將F2代株系群體種植于3個不同生態環境下,對每個生態環境下植株的株高、莖粗、葉長寬比(葉形)等10個農藝性狀進行測定,并對各農藝性狀進行變異分析、相關性分析及遺傳力分析,探究大麻F2代株系群體在不同生態環境種植下各農藝性狀間的關系及遺傳特點,以期為大麻不同用途品種選育及相關性狀的QTL定位提供理論參考。
1 材料與方法
1.1 試驗地點
2018年7—10月在云南省玉溪市峨山縣化念鎮、昆明市嵩明縣小街鎮和昭通市巧家縣馬樹鎮3個生態環境下開展F2代群體多點種植評價試驗。峨山縣化念鎮種植點經緯度為東經102°19′,北緯24°07′,海拔1120 m,年平均氣溫20 ℃,年平均降雨量828 mm,土質為沙壤土;嵩明縣小街鎮種植點經緯度為東經103°12′,北緯25°30′,海拔1910 m,年平均氣溫14.1 ℃,年平均降雨量996 mm,土質為紅壤土;巧家縣馬樹鎮種植點經緯度為東經103°25′,北緯26°75′,海拔2420 m,年平均溫度8 ℃,年平均降雨量726.4 mm,土質為紅壤土。
1.2 供試材料
本研究所用親本材料為中國第一個工業大麻品種‘云麻1號’(簡稱YM1)和國外引進品種‘DameBlanche’(簡稱Dame),將2份親本材料種植于昆明市盤龍區進行表型特性觀測統計(表1)。利用化學誘導性別轉換技術(專利號:ZL201410506082.7)開展了兩份材料的自交純合,獲得可用于F2代株系群體構建的兩份相對純合自交2代親本材料。

表1 兩份大麻親本材料的表型特性
1.3 F2代全雌性株系群體構建
從YM1與Dame自交2代材料中各選取1株雌株作為雜交親本,其中YM1雌株經化學誘導使其開雄花,授粉Dame雌株,收籽得到F1群體。選取F1群體中一個單株進行扦插繁殖得到一個克隆群體,化學誘導部分植株開雄花,授粉收籽即得到全雌F2群體。F2播種后,隨機選取300個單株進行編號,并將每一個單株扦插培育成株系,所有株系同時移栽至峨山縣、嵩明縣和巧家縣3個生態環境下,每個生態環境下每個株系確保3株克隆株(重復)。種植株行距為1.0 m×1.6 m,統一施肥、除草等田間管理。
1.4 農藝性狀測定及廣義遺傳力估算
參照大麻種質資源描述規范和數據標準[17]并結合本研究實際情況,于盛花末期對F2代株系群體中植株的株高、莖粗、第一分枝高、分枝數、分枝長、葉長(復葉)、葉寬(復葉)、葉長寬比(復葉)、小葉寬、鮮重共10個農藝性狀進行數據統計。其中,分枝長為植株中部最長兩個分枝的平均值,小葉寬為復葉最中間一片小葉的寬度。并按照以下公式對各性狀的廣義遺傳力進行估算[18]。
廣義遺傳力估算公式為:

1.5 數據處理
使用Excel 2016軟件對試驗數據進行描述統計和圖表繪制,使用SPSS 26.0軟件進行方差分析、相關性分析和均方計算。
2 結果與分析
2.1 雙親和F2代農藝性狀比較
F2代株系群體在3個生態環境種植下,除葉長在嵩明和巧家地區沒有表現出顯著差異(P>0.05)外,其它農藝性狀在3個生態環境下均表現出顯著差異。其中,株高、第一分枝高、分枝數、分枝長和葉長寬比5個農藝性狀平均值在嵩明地區種植的F2代株系群體中最大;莖粗、葉長、葉寬和小葉寬4個農藝性狀平均值則在巧家地區種植的F2代株系群體中最大;而在峨山地區種植的F2代株系群體中,所有農藝性狀平均值均為最小(表2)。

表2 大麻親本及其F2代農藝性狀數據比較
在3個生態環境種植下的F2代株系群體中,各性狀最大值均大于高值親本,最小值均小于低值親本(表3),表現出廣泛的性狀分離,豐富的性狀分離使群體中大部分性狀平均值最終居于雙親之間。而在峨山地區種植的F2代群體中,葉長寬比平均值則小于低值親本;在巧家地區種植的F2代群體中,分枝數平均值大于高值親本;3個生態環境下,小葉寬趨向或低于低值親本。
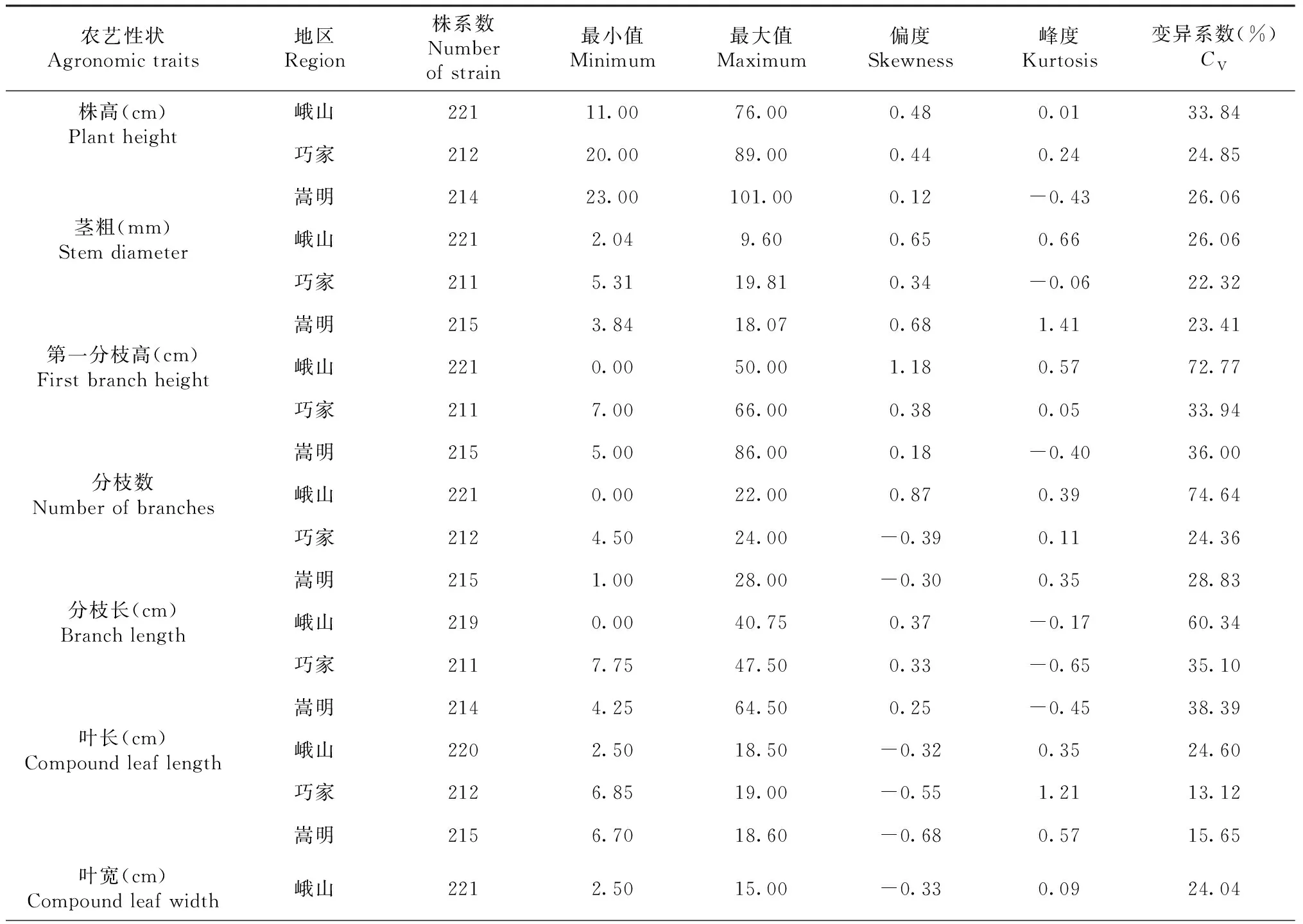
表3 F2代株系群體10個農藝性狀遺傳變異比較
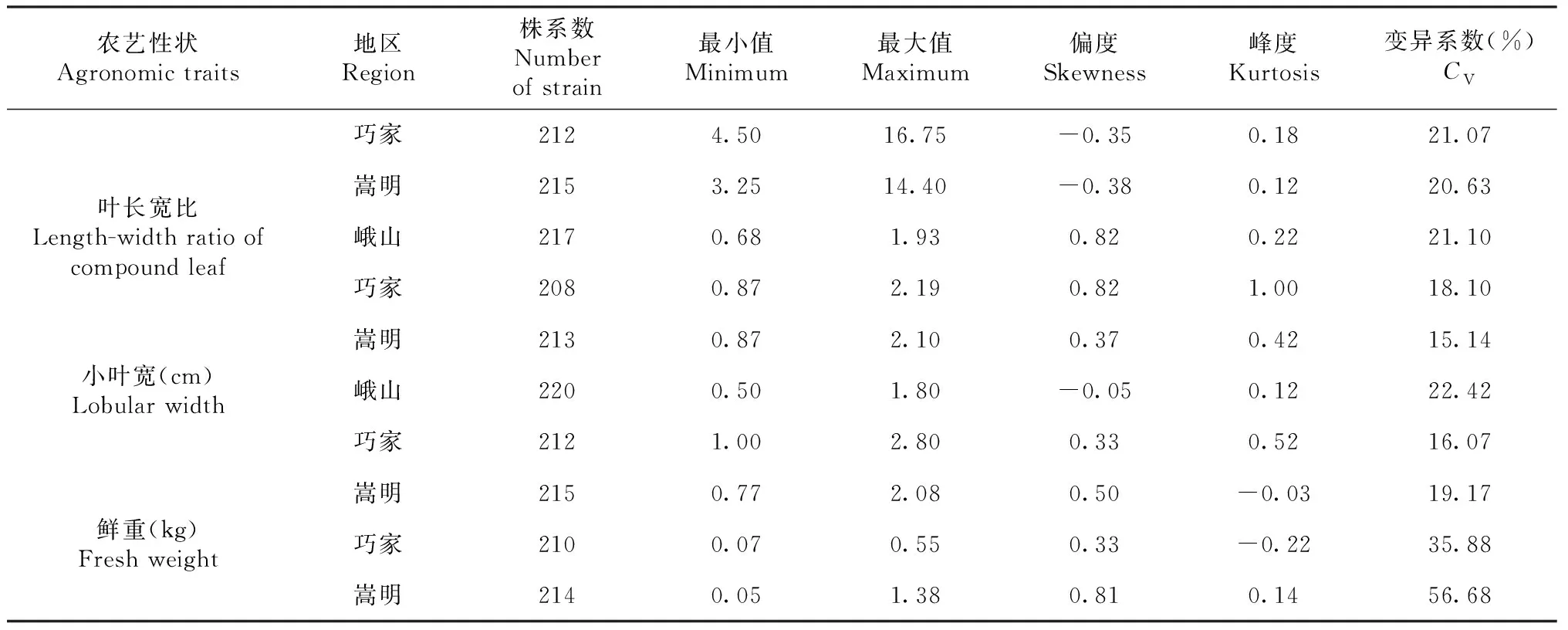
續表3 Continued table 3
2.2 F2代農藝性狀變異分析
通過對3個生態環境下F2代株系群體中10個農藝性狀進行變異分析,結果表明:10個農藝性狀變異程度差異較大,變異系數均大于10%,變異幅度為13.12%~74.64%(表3),10個農藝性狀平均變異系數(3個生態環境下變異系數之和/3)由大到小依次為:第一分枝高>鮮重>分枝長>分枝數>株高>莖粗>葉寬>小葉寬>葉長寬比>葉長。10個農藝性狀中,平均變異系數最小的為葉長,但其變異系數也達到了17.79%,表明這10個農藝性狀通過雜交選擇改良潛力較大。
某些性狀的變異程度在不同生態環境下表現出較大差異。峨山地區除鮮重數據缺失外,其余農藝性狀變異系數均大于另外2個地區。第一分枝高、分枝數、分枝長3個與分枝有關的農藝性狀在不同地區變異系數差異較大,分枝數在峨山和巧家2個生態環境下變異系數相差近50%,第一分枝高也相差近39%,而其它7個農藝性狀在不同生態環境下變異系數差異則相對較小。以上結果表明,對大麻不同農藝性狀進行選擇時要考慮其在特定環境中的變異程度,并根據不同的生態環境選育適應的品種。
2.3 F2代農藝性狀相關性分析
對在3個生態環境種植下的F2代群體中10個農藝性狀進行相關性分析。由表4~6可知,3個生態環境下,株高、莖粗、第一分枝高、分枝數、分枝長、葉長6個農藝性狀間均呈顯著或極顯著正相關。其中,株高與第一分枝高的相關系數最大,在3個生態環境下相關系數均在0.900以上;莖粗除在嵩明點與小葉寬、在巧家點與葉長寬比和小葉寬相關不顯著外,與其他農藝性狀均呈極顯著正相關;第一分枝高除在峨山點與小葉寬、在巧家點與葉寬、葉長寬比、小葉寬相關不顯著以及在嵩明點與小葉寬呈顯著負相關外,與其他農藝性狀均呈顯著或極顯著正相關;分枝數除在峨山點與小葉寬、在巧家點與葉寬、葉長寬比相關不顯著以及在嵩明點和巧家點與小葉寬呈極顯著負相關外,與其他農藝性狀均呈顯著或極顯著正相關。鮮重在有數據的嵩明點和巧家點與株高、莖粗、第一分枝高、分枝數、分枝長、葉長6個農藝性狀間均呈極顯著正相關。而葉長寬比、小葉寬在3個生態環境下與某些農藝性狀呈負相關。
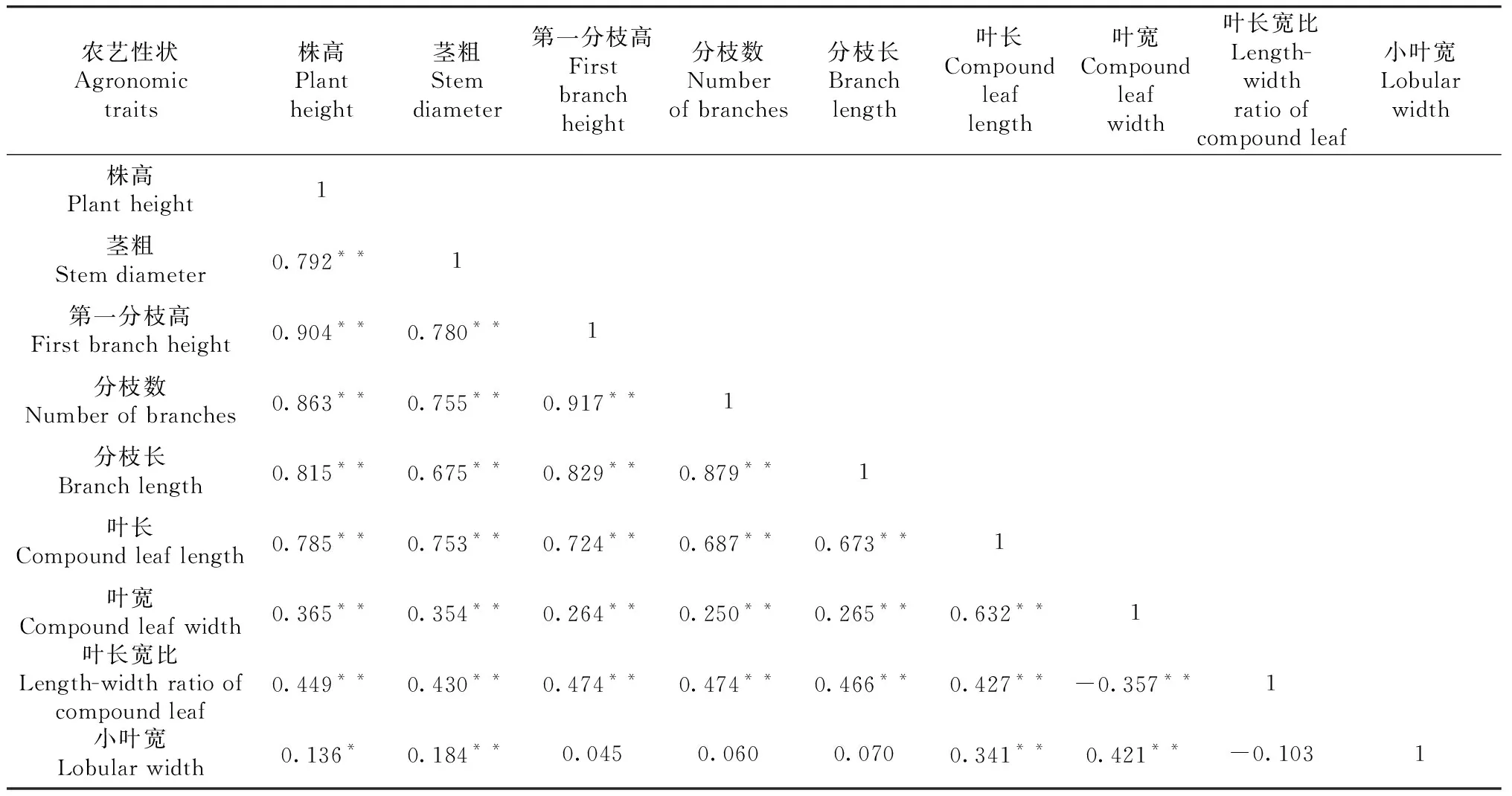
表4 峨山地區F2代群體中9個農藝性狀相關性分析
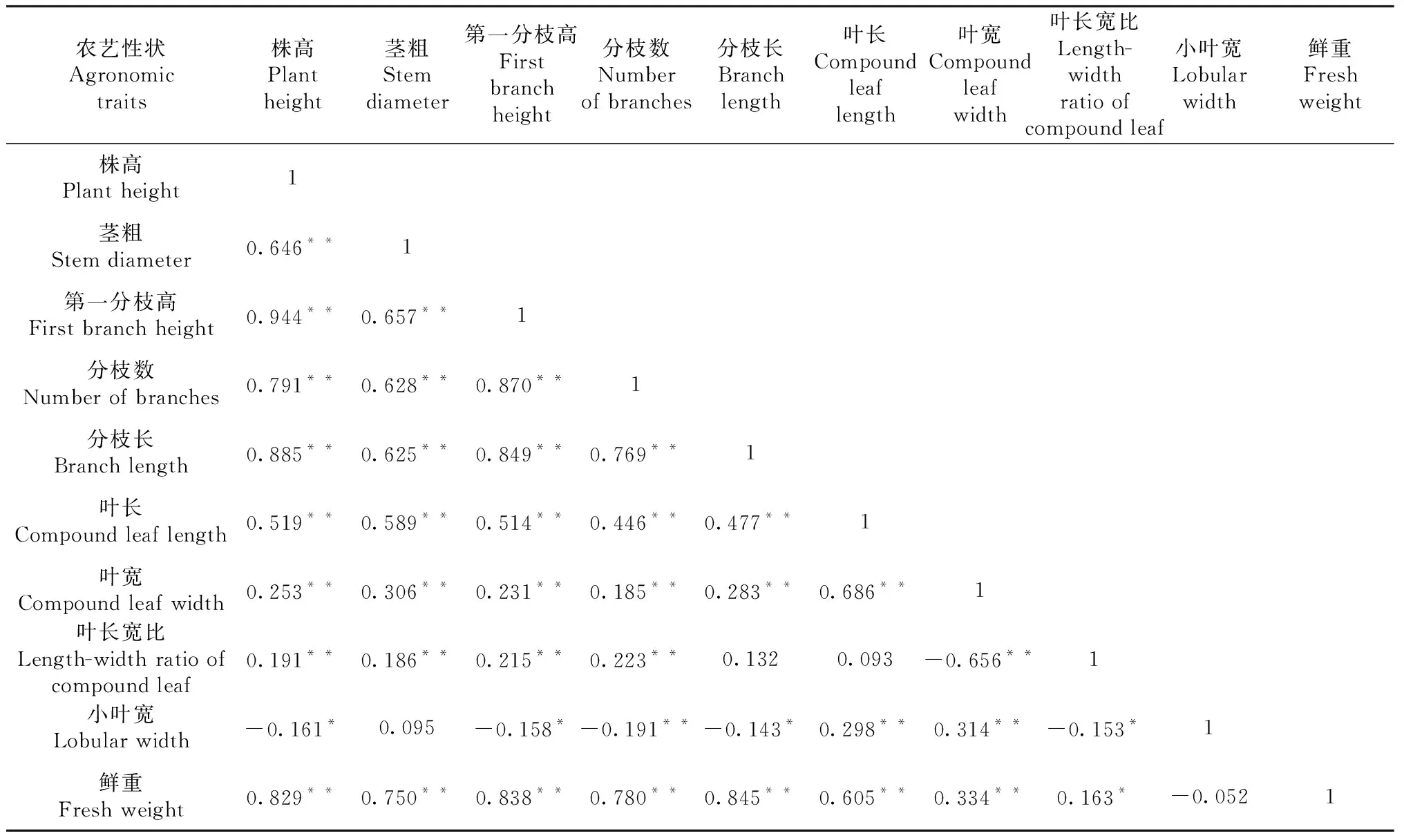
表5 嵩明地區F2代群體中10個農藝性狀相關性分析
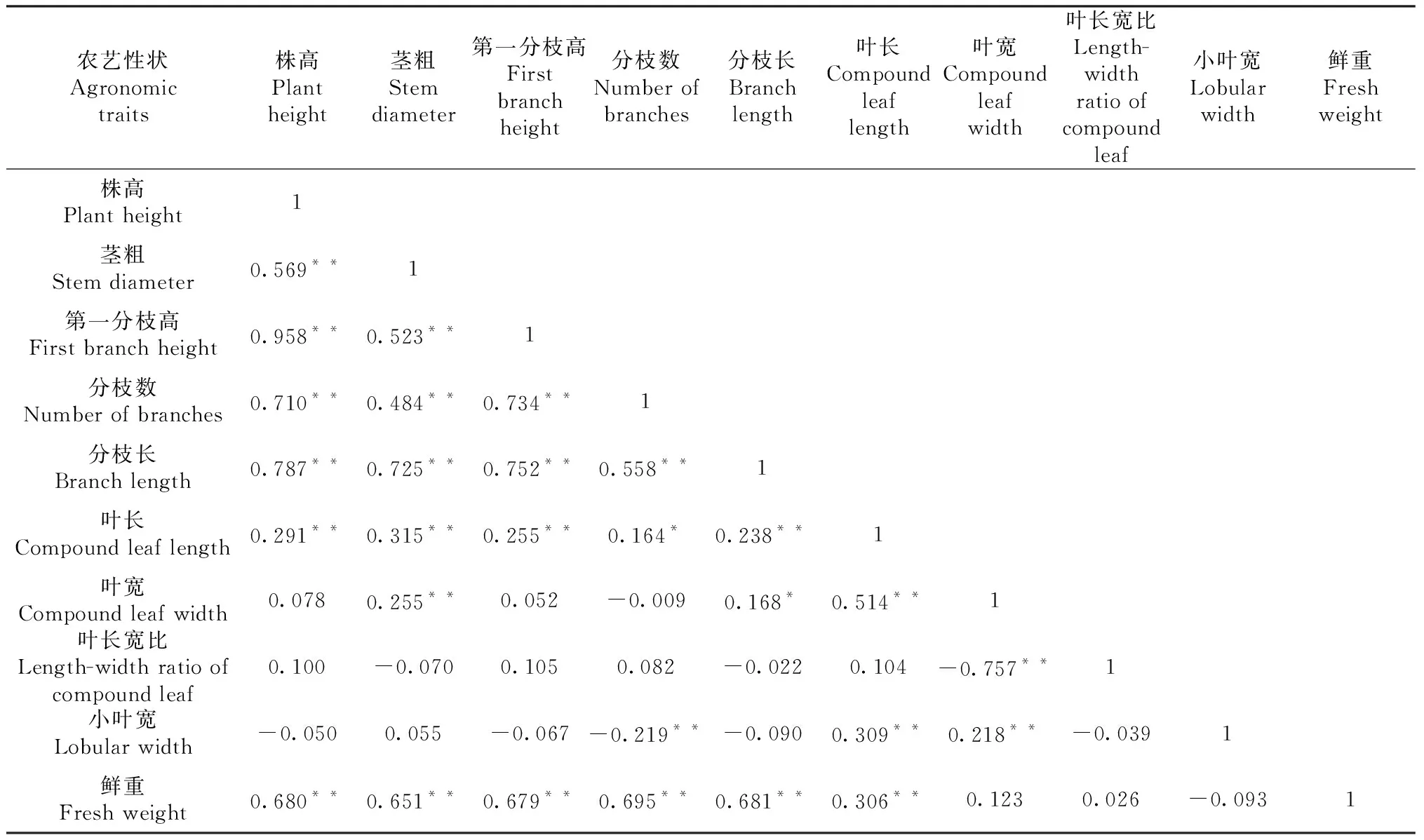
表6 巧家地區F2代群體中10個農藝性狀相關性分析
主要農藝性狀間相關性分析表明,不同生態環境下大麻主要農藝性狀相關性存在差異,如分枝數與葉長在3個生態環境下都表現出正相關性,但顯著性存在差異。在巧家地區種植的F2代群體中2個性狀相關性達到顯著水平,相關系數為0.164,而在峨山和嵩明地區種植的F2代群體中2個性狀相關性則達到了極顯著水平,相關系數分別為0.687和0.446。
2.4 F2代農藝性狀遺傳力分析
對10個農藝性狀進行頻率分布統計,結果表明10個農藝性狀頻率分布均表現出單峰分布,偏度和峰度絕對值在大部分生態環境下小于1(表3),且具有連續性較好的正態分布趨勢(圖1),表明10個農藝性狀屬多基因控制的數量性狀,但也有部分農藝性狀在不同的生態環境下表現出正偏性,如第一分枝高在峨山地區種植下,其偏度絕對值大于1,表現出明顯正偏性,而在另外兩個生態環境下偏度和峰度絕對值均小于1,表明不同的生態環境會對部分農藝性狀產生較大影響。
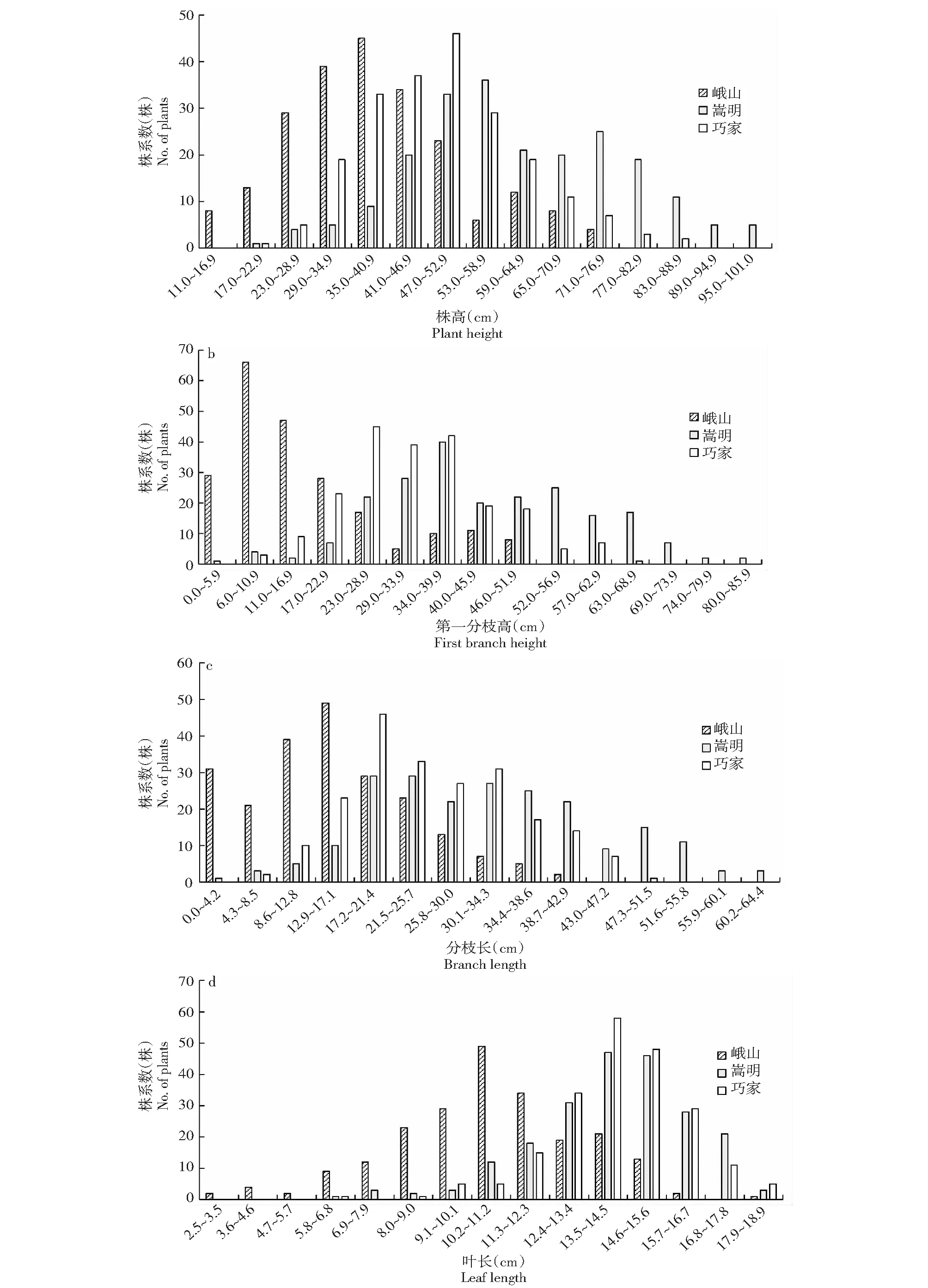
圖1 3種生態環境下4個農藝性狀頻率分布Fig.1 Frequency distribution of 4 agronomic traits in 3 ecological environments
將10個農藝性狀進行表型方差分析,結果表明,10個農藝性狀同時受到基因型、環境、基因型與環境互作(G×E)的極顯著影響(P<0.01)。估算出10個農藝性狀廣義遺傳力(表7),10個農藝性狀廣義遺傳力由大到小依次為分枝長>株高>第一分枝高>葉長>分枝數>莖粗>葉寬>小葉寬>鮮重>葉長寬比,其中前5個農藝性狀廣義遺傳力均超過65%,表明這5個農藝性狀主要受遺傳影響,具有較高的遺傳效應,廣義遺傳力最高的是分枝長和株高,分別為82.575%和81.811%。而廣義遺傳力最小的農藝性狀是葉長寬比,僅為29.508%,表明該性狀遺傳效應較小,主要受環境影響,由親本難以預測子代的表型。

表7 10個農藝性狀方差組成及廣義遺傳力估算
3 討 論
3.1 F2代群體中豐富的遺傳變異可為大麻品種選育提供材料來源
中國大麻種質資源豐富,種植歷史可追溯至5千年以上[19],并在長期的種植過程中形成了具有地方特色的種質資源,這些種質資源大多集中在籽用、纖維用和籽纖兼用等方面,且主要屬于sativa生態型,普遍存在植株高大、分枝稀疏、葉片大等特征,顯著區別于原產印度等南亞地區的indica生態型。隨著工業大麻多用途和機械化的開發應用,尤其是藥用工業大麻的興起,矮稈、早熟、多分枝、富含大麻素等農藝性狀備受關注。雜交群體農藝性狀的多樣性水平和變異程度對于培育作物新品種具有重要意義[20]。因此,本研究選取兩份來自不同生態型且遺傳背景差異較大的大麻材料作為親本,并成功構建F2代分離群體,對F2代分離群體中10個主要農藝性狀進行評價,結果表明10個農藝性狀分離廣泛,變異系數均大于10%,其中第一分枝高和鮮重變異系數較大,與湯志成等[14]通過對16份大麻種質資源進行表型變異分析,得出第一分枝高和地上生物量變異程度較大的結果一致。在本研究中,F2代分離群體的性狀變異相比自然變異更加豐富,表明利用大麻不同生態型材料進行雜交,可以在分離后代中獲得豐富的表型變異單株,這為不同用途的大麻品種選育提供了寶貴的材料來源。
3.2 不同生態環境對大麻植株農藝性狀的影響
盡管大麻分枝長、株高等性狀遺傳力較高,主要受遺傳控制,但不同的生態環境仍然對各農藝性狀產生較大影響,如峨山地區種植的F2代株系群體中表現出株型矮小、分枝數少、分枝較短等特點,而在嵩明和巧家地區種植的F2代株系群體則株型相對高大、第一分枝明顯增高、分枝數顯著增多。部分農藝性狀對環境敏感可理解為是對環境的適應,但在實際栽培中要特別注意對環境比較敏感的農藝性狀,這部分農藝性狀往往決定著品種的適應范圍。如適合在北方種植的大麻引種至南方則會提前開花,生物量降低[21]。而南方大麻移栽至北方則往往表現為遲熟,株型高大,生物量增加[22]。
不同生態環境對F2代株系群體中部分農藝性狀產生顯著影響的原因可能在于土壤、海拔和氣候等環境因素不同而導致,尤其是光照對大麻生長發育的影響,因大麻是短日照植物,對光周期特別敏感[23],而大麻生物量主要由營養生長期積累,當光照時長低于開花所需臨界時長大麻植株會進入生殖生長[24],導致生物量積累速度放緩,株型變得矮小。本研究中,雖然同一生態環境下F2代分離群體開花時間相對一致,但在不同緯度的3個環境下開花時間還是產生了差異,開花時間順序呈現出峨山略早于嵩明,嵩明略早于巧家,導致除葉長外各農藝性狀在3個生態環境下均產生顯著差異。因此,在大麻育種和栽培過程中,應特別注意環境對大麻農藝性狀和生物產量的影響,并為不同用途和栽培環境選育合理的株型品種。同時也可通過改良栽培措施,促進某些性狀的發育,從而達到增產的目的。
3.3 性狀相關性分析和廣義遺傳力估算對農藝性狀選擇的影響
通過分析各農藝性狀間關系,可以根據育種目的合理協調各農藝性狀的關系,獲得理想株型,提高育種效率。本研究相關性分析表明10個農藝性狀間關系密切,除小葉寬、葉寬、葉長寬比3個性狀外,其余農藝性狀間均兩兩呈顯著或極顯著正相關。其中,株高與莖粗、葉長、葉寬都存在正相關,這與房郁妍等[25]對13份大麻種質材料農藝性狀相關性研究結果一致。各性狀間關系密切提示在大麻育種過程中,應綜合多個農藝性狀進行分析,在對目標性狀進行選擇的同時要考慮到對其關聯性狀的影響。
遺傳力是研究數量性狀遺傳差異的主要參數之一,遺傳力的大小可以為農藝性狀世代選擇提供參考[26],在本研究構建的F2代株系群體中株高和莖粗的廣義遺傳力分別為81.811%和64.204%,這與Petitd等[16]在3個生態環境下對123份大麻種質資源進行評價,得出株高和莖粗廣義遺傳力分別為0.81和0.69的結果基本一致。通過對各農藝性狀進行廣義遺傳力估算,結果表明分枝長、株高、第一分枝高、葉長和分枝數5個性狀廣義遺傳力均超過65%,表明這5個性狀主要由遺傳因素決定,在品種選育時可以從早期世代進行選擇。而葉長寬比和鮮重廣義遺傳力則較低,表明這2個性狀易受到環境的影響,早期世代不宜對這兩個性狀進行選擇。
4 結 論
選用2份來自不同生態型且遺傳背景差異較大的大麻材料作為親本,成功構建F2代全雌性株系群體。對3種生態環境下F2代株系群體中10個農藝性狀進行遺傳變異及相關性分析。結果表明:8個農藝性狀在3種生態環境種植下表現出顯著差異(P<0.05);10個農藝性狀表現出廣泛的性狀分離,為大麻不同用途品種選育創造了豐富的遺傳變異類型;株高、分枝數、分枝長等性狀頻率分布具有典型的數量遺傳特征,可通過QTL對相關基因進行定位;相關性分析表明10個農藝性狀關系密切,提示在育種過程中突出單個性狀的同時要考慮到對其關聯性狀的影響;分枝長、株高、第一分枝高、葉長及分枝數的廣義遺傳力均超過65%,其中分枝長和株高廣義遺傳力超過80%,可為大麻育種過程中農藝性狀世代選擇提供理論參考。以上結果可為大麻不同用途品種選育和相關性狀的QTL定位提供一定的理論基礎。
