塞罕壩國家森林公園褐柳鶯繁殖生物學研究
2022-10-08 03:16:50汪揮勝祃來坤王姣姣侯建華
四川動物 2022年5期
關鍵詞:研究
汪揮勝,祃來坤,2,王姣姣,侯建華*
(1.河北大學,河北 保定 071000;2.河北民族師范學院,河北 承德 067000)
鳥類生活史是生態學研究的重要領域,對生活史理論的發展作出了重要貢獻(Partridge&Harvey,1988)。了解鳥類生活史特征對于理解其種群動態、個體特征以及溫帶、熱帶物種之間的差異具有重要作用(Martin,1995)。繁殖是鳥類生活史中最重要的組成部分,包括配偶選擇、巢址選擇、產卵、孵化和育雛(鄭光美,2012),這些通過自然選擇形成的一系列適應性行為,有利于個體和種群的生存(Saether & Bakke,2000)。但全球僅1/3的鳥類有較詳細的繁殖資料,而雀形目Passeriformes不足7%(Xiao,2017),因此,繁殖資料的收集仍是現階段鳥類研究中重要和長期的工作。
褐柳鶯隸屬于雀形目柳鶯科Phylloscopidae,在河北塞罕壩國家森林公園為夏候鳥,活動于灤河岸邊不同蓋度和高度的灌木叢中,種群密度大(Liu,2017)。目前,僅有1篇關于俄羅斯遠東地區褐柳鶯應對花鼠捕食的反捕食策略的研究(Forstmeier &Weiss,2004),缺乏褐柳鶯的完整繁殖信息。本研究于2021年5—8月對繁殖于塞罕壩國家森林公園的褐柳鶯進行了詳細研究,并采用全程錄像的方法對其繁殖過程進行了觀察。
1 研究區域與研究方法
1.1 研究區域
塞罕壩國家森林公園(116°51′~117°39′E,42°2′~42°36′N,海拔1 350~1 650 m)位于河北省最北端,植被群系主要包括落葉針葉林、常綠針葉林、針闊混交林、闊葉林、灌叢或灌草叢、草原與草甸等。屬寒溫帶大陸性季風氣候,年均氣溫-1.2℃;年均降水量452 mm,年均相對濕度68.8%;無霜期69 d,雪季可達7個月(Liu,2019)。根據褐柳鶯的繁殖習性,調查范圍集中在塞罕壩國家森林公園三道河口管理處與五間房之間的沿朝格都爾河河道兩側的林地、灌叢及苗圃地(圖1)。
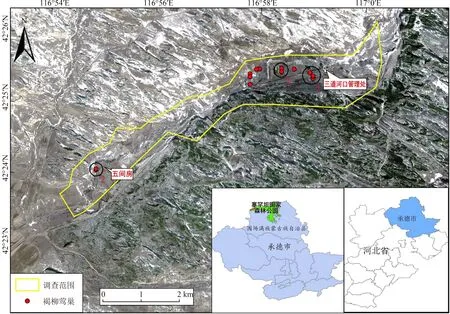
圖1 研究區域及巢分布Fig.1 Study area and nest distribution
1.2 研究方法
2021年5—8月通過跟蹤處于繁殖階段的成鳥和系統搜索法尋找褐柳鶯的繁殖巢(正在筑巢或者至少包含1枚卵或者幼鳥)(Martin & Geupel,1993),搜索區域包括研究區的針葉林、灌木林、灌草叢以及人工苗圃,找到后用GPS進行定位,共18個,對其中13個使用微型攝像機(HiLEME,中國)進行持續監測,其余5個采用隔天人工探查的方式進行監測,直到繁殖失敗或者雛鳥離巢結束。
微型攝像機安裝在可完整監視到整個巢口的樹枝上或者人工輔助放置在巢旁的木棍上。收集和記錄繁殖巢以下參數:(1)巢參數:巢材、巢距地面高度(cm)、巢所在樹高(cm)、巢口直徑(mm)、巢深(mm)、巢長(mm)、巢寬(mm)和巢高(mm);(2)卵參數:包括窩卵數、卵徑長(L/mm)、卵徑寬(B/mm)、卵體積(V=0.51LB)(Hoyt,1979)和卵重(g);(3)雛鳥參數(從雛鳥出殼日起,每隔2 d測量一次雛鳥身體參數,測量時間為每天下午):雛鳥體質量(g)、跗趾長(mm)、嘴峰長(mm)和雛鳥離巢日期;(4)繁殖成功巢指至少有1只雛鳥存活到離巢,繁殖成功率為繁殖成功巢占所有巢的比例(Yang.,2011)。長度采用游標卡尺(卡夫威爾,中國;精度:0.01 mm)測量,質量采用電子秤(DH,中國;精度:0.01 g)測量。為了減少干擾,所有巢參數的測量均在該巢繁殖活動結束后進行。
1.3 數據分析
2 結果
2.1 繁殖期及繁殖成效
褐柳鶯于4月末至5月上旬到達研究區,5月底開始筑巢,繁殖活動持續到8月中旬,8月下旬到9月上旬開始陸續遷走,繁殖期持續約4個月,6—7月為營巢高峰期,88.89%(16/18)在這期間產下了第一枚卵。
褐柳鶯繁殖成功率為27.78%(5/18)。在13個失敗巢中,9巢被捕食,巢捕食導致繁殖失敗占比69.23%。根據錄像監控以及現場痕跡調查,6巢的捕食者為赤狐,1巢捕食者為牛頭伯勞,其他巢的捕食者未被記錄(表1)。
2.2 巢及巢址特征
18個褐柳鶯巢為側面開口的球形巢(圖2),均分布在人工苗圃中,其中,云杉幼苗上9個,占總數的50.00%;樟子松幼苗上8個,占44.44%;地面1巢,占5.56%。巢位高度為(31.83±5.40)cm(=18),巢所在樹高為(188.44±20.89)cm(=18)。巢口直徑(56.51±2.08)mm、巢深(73.79±2.81)mm、巢長(132.23±3.30)mm、巢寬(106.07±4.68)mm、巢高(99.91±2.93)mm。巢距地面的高度顯著低于巢距灌木頂部的距離(=-4.873,<0.01),表明巢筑在植被中下部。
2.3 產卵及孵卵行為
首枚卵出現在5月28日,最晚為7月14日。褐柳鶯每天06∶00—09∶00產下1枚光滑無斑點的白色卵(圖2)。窩卵數(4.00±0.15)枚(=10),主要為4枚(8/10);孵卵期為(14.00±0.45)d(=5);卵徑長(16.88±0.16)mm(=17),卵徑寬(13.11±0.16)mm(=17),卵體積(1 484.15±32.74)mm(=17),鮮卵重為(1.39±0.02)g(=17)(表1)。
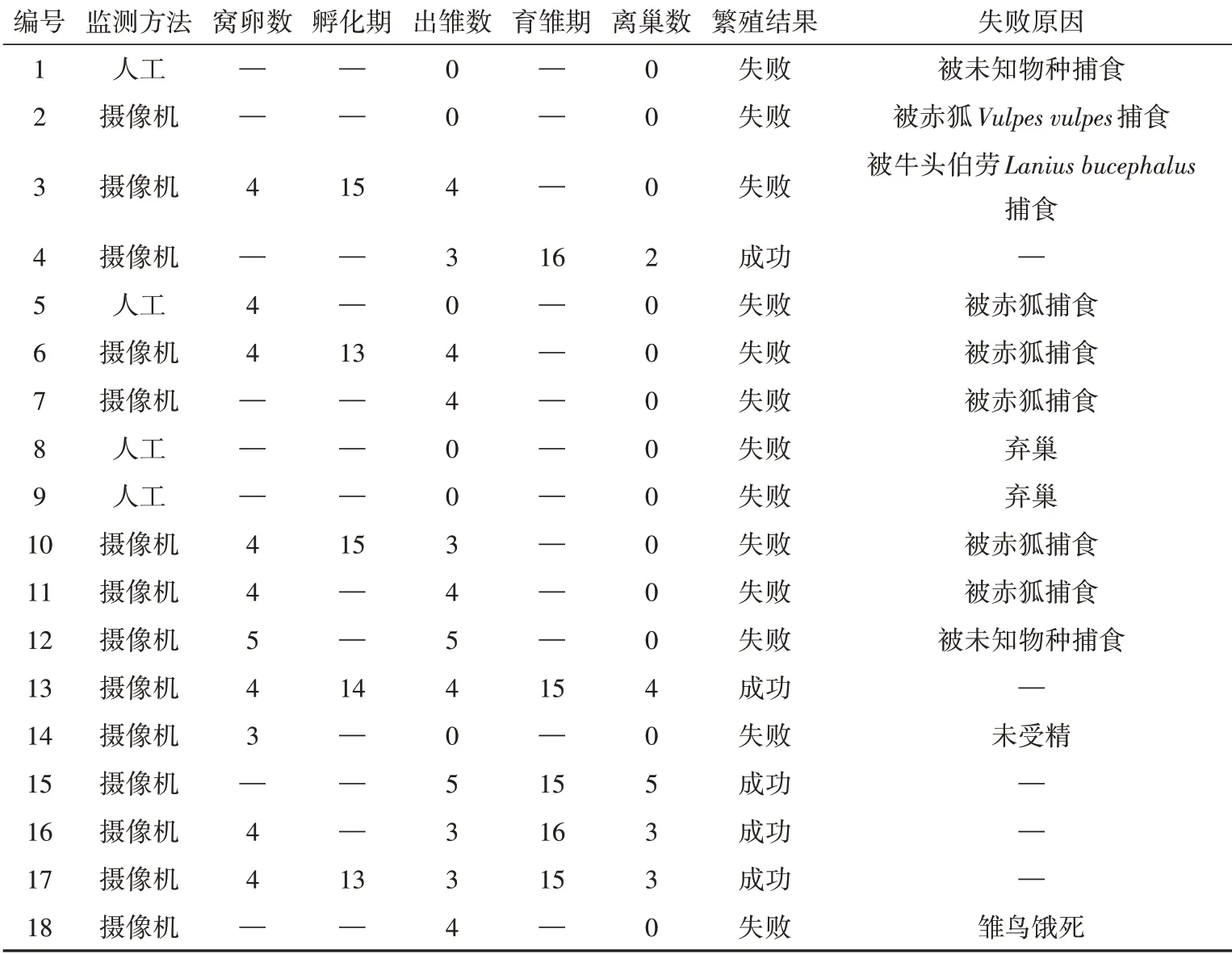
表1 2021年塞罕壩國家森林公園褐柳鶯繁殖數據Table 1 Breeding data for Phylloscopus fuscatus in the Saihanba National Forest Park in 2021

圖2 褐柳鶯的巢址生境(左)、巢和卵(右)Fig.2 Nest-site(left),nest and egg(right)of Phylloscopus fuscatus
雌鳥產完最后1枚卵或倒數第2枚卵開始孵卵,開始孵卵后雌鳥在巢中過夜。根據2巢的完整錄像發現親鳥日孵卵頻次在整個孵卵期沒有明顯的波動,而日孵卵時長波動較大,但每日04∶00—20∶00的總孵卵時長都大于600 min。不同巢的每小時孵卵頻次和孵卵時長不同,但都表現出相同的變化趨勢:黎明的孵卵頻次低,時長較長;黃昏孵卵頻次與其余時間無明顯差異,但孵卵時長較長;每日孵卵的最短時長出現在09∶00—13∶00,從黎明到中午孵化時長呈遞減趨勢,中午到黃昏時長呈遞增趨勢(圖3)。每日翻卵頻次為(163.21±11.15)次(=4),各巢翻卵頻次未呈現規律性變化。
2.4 育雛及雛鳥發育
育雛期為(15.4±0.2)d(=5)。根據錄像,雛鳥破殼后卵殼被雌鳥叼出巢。在育雛過程中,親鳥在雛鳥排出糞囊后,將糞囊叼離巢區。雛鳥孵出當日,親鳥坐巢頻次顯著增加,在雛鳥孵出后0~5 d,親鳥的坐巢次數都高于孵化期,但坐巢的時間總體呈下降趨勢,直到雛鳥11日齡,親鳥完全停止暖雛行為,再繼續喂養雛鳥(5.6±0.2)d(=5),至雛鳥離巢(圖3)。
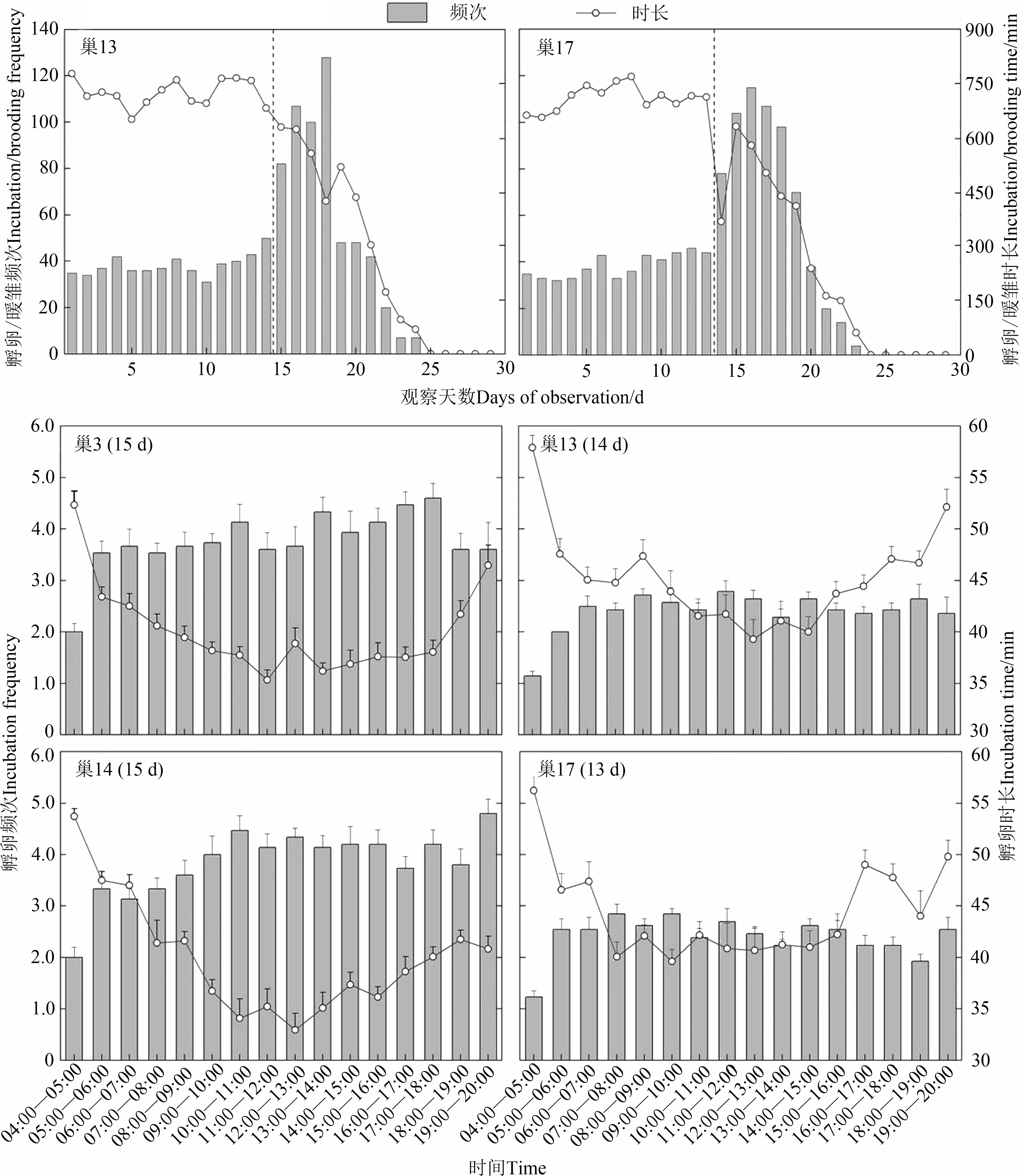
圖3 褐柳鶯的孵卵/暖雛行為Fig.3 Incubation/brooding behaviour of Phylloscopus fuscatus
從每日育雛頻次來看,從第一只雛鳥孵出,到雛鳥離巢的前一天,親鳥每日喂食頻次逐漸增加,直至雛鳥離巢當日,喂食頻次才有所減少。在每小時喂食頻次上,05∶00—20∶00的每小時喂食頻次并未出現較大波動,但04∶00—05∶00的喂食頻次明顯少于其他時間段,而總體呈現出黃昏喂食頻次略低于其他時間段。每日清巢次數為(30.90±1.93)次(=2)。雛鳥排便總是在親鳥喂食后發生,雛鳥排便的頻次和親鳥喂食的頻次高度相關(r=0.705,<0.01)(圖4)。
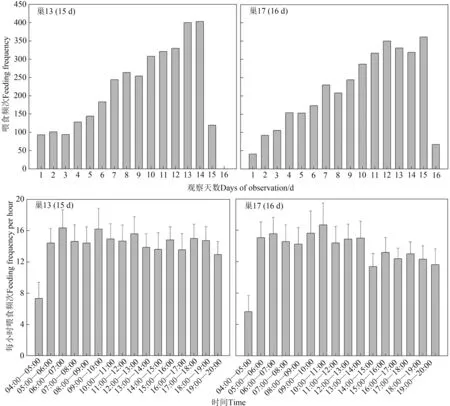
圖4 褐柳鶯喂食行為Fig.4 Feeding behaviour of Phylloscopus fuscatus
雛鳥部分日齡外部形態如下:1日齡雛鳥閉著眼睛,皮膚大部分裸露無毛,僅頭頂和眼周有少量絨毛;皮膚肉紅色,眼泡黑色且突出,不能站立,蜷縮成團。3日齡雛鳥皮膚顏色加深,背部及翅膀皮下出現黑色羽管。6日齡雛鳥頭部呈黑色,背羽、尾羽和初級飛羽開始生長。9日齡雛鳥飛羽和頭部羽毛完全覆蓋皮膚,僅背部有皮膚裸露,并且能夠伸長脖子乞食。12日齡雛鳥全身被羽,飛羽前端為藍色,末端為黑色,能夠長久站立。15日齡雛鳥羽毛和成鳥相似,顏色比成鳥略深,尾羽較短。
Logistic回歸擬合雛鳥跗趾長和體質量的生長曲線,獲得3個身體參數的生長曲線擬合方程,體質量的為:=9.465 87/[1+14.844 94exp(-0.571 79)],跗 趾 長 的 為:=25.120 52/[1+3.794 8exp(-0.3)],嘴 峰 的 為:=6.740 56/[1+2.791 7exp(-0.263 63)](圖5)。
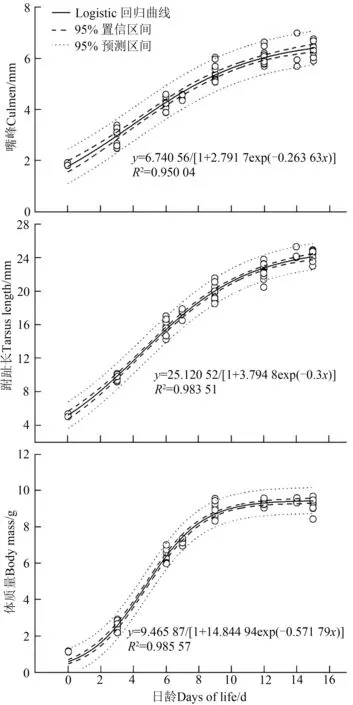
圖5 褐柳鶯雛鳥不同部位的生長曲線Fig.5 Growth curves of different body parts of Phylloscopus fuscatus nestlings
3 討論
褐柳鶯遷徙到塞罕壩地區繁殖的時間比更高緯度的俄羅斯遠東地區的繁殖種群(5月末遷入)更早,而遷走時間與俄羅斯遠東地區繁殖種群相近,這可能與不同緯度地區的氣候以及遷徙距離差異有關。塞罕壩地區繁殖的褐柳鶯種群越冬地為中國南部和東南亞地區,而俄羅斯遠東地區繁殖的褐柳鶯種群越冬地同樣位于東南亞地區(von Blotzheim,1966)。在其他鳥類中發現,高緯度地區繁殖的種群需經過更長的遷徙路線從越冬地到達繁殖地(Burman,2018),相比于塞罕壩地區的種群,在俄羅斯遠東地區繁殖的種群可能需要遷徙更遠的距離,同時,繁殖地的氣溫也會影響越冬地鳥類開始春遷的時間(Kanerva,2020),俄羅斯遠東地區春季回暖時間更晚。
鳥類窩卵數是鳥類學家長期以來一直探討的問題,也是鳥類生活史對策理論的重要研究內容之一。窩卵數變異原因主要包括地理變異、季節性變化、其他因素(棲息地類型、種群密度等)(鄭光美,2012)。俄羅斯遠東地區緯度為59°51′N,褐柳鶯的窩卵數平均為5.16枚(Forstmeier,2001);塞罕壩地區緯度為42°36′N,窩卵數平均為4.00枚,這與大多數研究一致,即窩卵數隨緯度增加 而 增 加(Griebeler,2010;Winkler,2014)。目前關于窩卵數地理變異原因的討論集中在資源供應、氣候差異以及捕食風險3個方面(Bourgault,2010;Mér?,2014)。受塞罕壩地區與俄羅斯遠東地區的緯度差異以及林地類型影響,塞罕壩地區窩卵數較俄羅斯地區小可能同時受到以上3個方面的影響。資源供應方面:塞罕壩地區為次生人工林地,相較于俄羅斯遠東地區的原始林地,褐柳鶯可獲得的食物資源可能會更少;氣候差異方面:俄羅斯遠東地區的緯度更高,氣溫回暖時間更晚,因此褐柳鶯遷到的時間更晚,而遷走時間相近,導致可用于繁殖的時間更短。在繁殖結束后,俄羅斯遠東地區褐柳鶯種群需要遷徙更長距離到達越冬地,在更長距離的遷徙中,幼鳥的死亡數量可能會高于遷徙距離更短的塞罕壩種群,因此俄羅斯遠東地區更大的窩卵數可以更好維持種群數量穩定。從捕食風險來看:塞罕壩地區褐柳鶯更小的窩卵數不僅適應了低緯度地區更高的巢捕食風險,同時減小了因巢捕食帶來的損失。塞罕壩更長的繁殖季節,讓其在首次繁殖失敗后,有機會進行第二次繁殖,從而進一步減小巢捕食對種群數量穩定帶來的影響。
孵卵投資是鳥類孵卵行為中最受關注的方面,親鳥需權衡在覓食和孵卵之間的時間分配,因為它們需要在維持自身需求的同時保證胚胎的正常 發 育(Cooper & Voss,2013;Dammhahn,2018)。由于褐柳鶯的雌雄二性區別不明顯,無法通過視頻準確判斷雌性是否單獨孵卵,但所有巢的監控記錄中均未觀察到交替孵化行為,且存在其他成鳥向孵卵成鳥遞食的現象;同時在成鳥孵卵過程中,巢所在領地仍有雄性鳴唱,因此推測孵卵行為可能由雌性單獨完成。根據白天每小時孵化時間的分配情況,親鳥采取高頻次、單次孵卵時間短的孵化策略,并且在09∶00—15∶00投入最少的時間,表明雌性覓食主要發生在這個階段。這種孵化策略可能是親鳥對自身需求以及胚胎發育需求進行權衡的結果。這種時間分配模式非常合理,中午由于氣溫上升,卵的溫度會比上午和傍晚更高,此時昆蟲也更加活躍,分配更多時間覓食既能保證卵的溫度不會太低導致胚胎死亡,也能保證親鳥獲取充足的食物。Drent等(1970)發現,由于胚胎在孵化后期需要更多的熱量,親鳥會減少孵化頻次,增加總體的孵化時長,但是本研究在孵化階段沒有發現這種后期孵化時長增加的現象。
清理糞囊是育雛期的一種重要行為,也是清巢行為的重要組成部分,不僅可以減少雛鳥被寄生蟲感染的風險,亦可以降低捕食者通過糞便氣味和痕跡定位巢的概率(Guigueno & Sealy,2012;Ibá?ez-álamo.,2014)。Quan等(2015)發現雀形目雛鳥排便和親鳥喂食具有高度的協同性,雛鳥總是在親鳥喂食后就進行排便,以此保證親鳥能夠及時叼走糞囊,本研究與上述結果相似,褐柳鶯雛鳥的排便與親鳥喂食行為較為統一。
繁殖成效分析表明,本地區褐柳鶯的繁殖成功率僅為27.78%,其中巢捕食是導致繁殖失敗的主要原因,這與許多柳鶯屬spp.鳥類報道的捕 食 率 相 似(Smith,2007;Roncalli,2016)。本地區主要巢捕食者為赤狐,這可能與赤狐在研究區較高的遇見率相關;此外,巢址選擇也是影響巢捕食的重要因素(Jara,2020),本研究中褐柳鶯主要在人工苗圃地中繁殖,這與其他地區的筑巢生境不同(Forstmeier & Weiss,2004),這種巢址選擇模式是否是褐柳鶯應對高捕食壓力的繁殖對策以及對繁殖成效的影響需要進一步研究。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
