瀕危植物香果樹根萌苗和實生苗的種群特征比較
2022-10-15 09:02:48薛蘋蘋郭連金毛小濤肖志鵬吳艷萍王寶云范紅霞
天津師范大學學報(自然科學版) 2022年5期
關鍵詞:生長
薛蘋蘋,郭連金,王 帆,毛小濤,肖志鵬,吳艷萍,王寶云,曾 穎,范紅霞
(1.上饒師范學院生命科學學院,江西 上饒 334001;2.山東省慶云縣農業局,山東 德州 253700)
植物種群的自然更新是一個復雜的生態學過程,它對種群的增殖、擴散、延續以及穩定具有重要作用[1].植物種群自然更新的前提是有足夠數量的幼苗存活[2],它是打破瀕危植物瀕臨滅絕的關鍵.目前,對大多數瀕危植物的研究主要集中于種群結構及空間分布格局[3-4]、種子雨與種子庫[5]、種子萌發及幼苗生長[6]、光合作用與蒸騰作用[7]以及遺傳多樣性[8-9]等方面.原生生境中部分瀕危植物如峨眉冷杉、蒙古扁桃、沙冬青等雖然擁有較高的種子產量和擴散范圍,但由于種子被捕食、霉爛以及遭遇環境脅迫等,種子萌發數量或形成幼苗的數量較少[5,10],且大部分物種的實生苗生長緩慢和抗性較差[11],因此在原生生境中有些瀕危植物的自然更新主要依賴于伐樁和根上萌蘗產生的萌苗來完成[12-13].有研究表明樹木的伐樁及其損傷的根系可產生大量的萌苗[14-15],但伐樁萌苗和根萌苗對植物種群恢復的貢獻鮮見報道[16-17],關于伐樁萌苗、根萌苗和實生苗對植物種群更新貢獻的比較更為少見[18].科學認識一種植物的天然更新特性是制定合理的人工促進天然更新措施以及進行自然資源保護的生態學基礎[15].
香果樹(Emmenopterys henryi)為中國特有的茜草科(Rubiaceae Juss.)單種屬落葉喬木,是研究茜草科系統發育、形態演化及中國植物地理區系的重要材料.近年來,由于環境及人為干擾等因素的影響,該物種已瀕臨滅絕,被列為國家Ⅱ級重點保護稀有植物和林業部公布的國家珍貴樹種[19].香果樹種子微小,壽命短,屬于光敏感型種子,千粒重僅為0.50 g左右,原生生境中發芽率極低[20].原生生境中的香果樹種群主要存在實生苗和根萌苗2種更新方式[13,21-22].香果樹群落郁閉度較高,且幼苗主要存在于喬木樹冠下,光照條件較差[13],對于此環境中香果樹這2種起源幼苗的生長狀況以及它們在香果樹種群自然更新中的作用尚無人報道.本研究以武夷山國家級自然保護區內的香果樹種群為研究對象,比較實生苗和根萌苗的年齡結構、數量特征、生長特征、自然更新能力以及對樹蔭的忍耐力,旨在闡明原生生境中香果樹不同起源幼苗的生存狀況及香果樹種群的更新策略,為香果樹種群恢復提供理論依據和實踐指導.
1 研究地自然概況
研究地位于武夷山國家級自然保護區(27°33′~27°54′N,117°27′~117°51′E),是世界同緯度現存面積最大、保存最完整的中亞熱帶森林生態系統,區內自然資源豐富,保存有完好的地帶性常綠闊葉林群落,是東南亞大陸生物多樣性最豐富的地區,也是具有全球生物多樣性保護意義的地區.本區氣候屬于典型的亞熱帶季風氣候,平均氣溫為12~13℃,年降水量1 480~2 150 mm,年蒸發量1 492 mm左右,相對濕度78%~84%,無霜期253~272 d.土壤類型主要有紅壤、黃紅壤、黃壤和山地草甸土[23].香果樹在研究地主要星散分布于溝谷水溪旁的闊葉林中,其伴生種主要有銀鵲樹(Tapiscia Sinensis)、杉木(Cunninghamia lanceolata)、紅脈釣樟(Lindera rubronervia)、鹽膚木(Rhus chinensis)、野鴨椿(Euscaphis japonica)等.
2 研究方法
2.1 樣地調查
在對武夷山自然保護區進行全面調查的基礎上,在香果樹分布比較典型的區域設置4個樣地,面積為20 m×20 m,自然概況如表1所示.
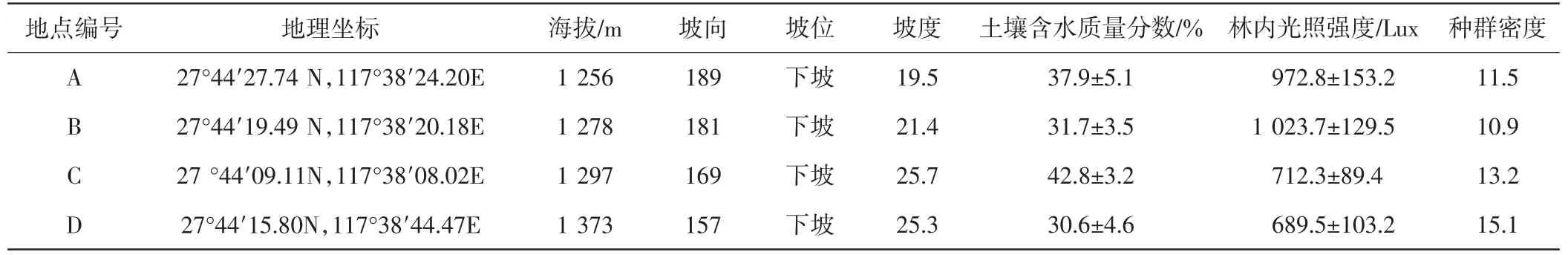
表1 香果樹樣地概況Tab.1 Genernal situation of the habitats of E.henryi
調查樣地生境:①確定生境的地貌地形、土壤、坡向、坡位、光照、溫度、土壤含水量等生態因子,使用ZDS-10型光照計測定光照強度,使用DHM2型通風干濕溫度計測定大氣溫度和濕度,在距地面0.5 m處進行測定[13].②確定不同起源的幼苗,觀察種子萌發和幼苗根系情況以確定幼苗來源,標記實生苗和根萌苗.
2.2 不同起源幼苗的種群結構及生長特征
2015年11月,將樣地內所有幼苗按照起源分類,采用幼苗主莖芽鱗痕的數目、節間的長短規律以及莖干表皮與周皮的紋路確定其年齡,每增長1 a設置1個年齡級,即1 a、2 a、3 a,以此類推.統計林內1~8 a幼苗中每級個體株數,作為該種群動態分析的基礎數據.根據每級個體數的統計繪制年齡結構圖.根據種群各齡級株數編制生命表,根據生命表中的標準化存活數以及死亡數繪制存活曲線和死亡曲線[24].記錄每株幼苗的高度、基徑、葉數,并利用活體葉面積儀(LA-S,美國CID公司)測定其葉面積,摘取葉片自然晾干,測定其干重并計算比葉重,以了解不同起源香果樹幼苗的生長動態.
2.3 不同微生境中幼苗的生長特征
隨機標記林內香果樹母樹或其他喬木的樹冠下、冠緣、林窗以及林緣空地的香果樹實生苗和根萌苗各10株,記錄并測定其1年生側枝的基徑、枝長、葉數、節數、葉面積以及葉厚.利用打孔器于每個新生枝條上隨機選取20個健康成熟的葉片,在主脈兩側各打3個圓片,其中30個圓片測其鮮重,自然晾干后再測其干重,以計算葉片含水量,另外30個圓片放入冰箱后迅速帶回實驗室進行葉綠素含量的測定[25].
2.4 不同微生境中幼苗的光合作用特征
于2015年8月,選擇晴天,利用便攜式光合儀(CI-340,美國CID公司)對所標記香果樹幼苗的葉片進行光合作用測定,從8∶00—16∶00每隔2 h測定1次,測試采用開放式氣路,測定植物的凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)以及蒸騰速率(Tr).為了使2種幼苗的光合作用具有可比性,均選取1年生枝條由上到下的第3對葉片,每株選擇6片,每次測定時間為30 min.
2.5 數據處理
采用SPSS19.0對數據平均值進行單因素方差分析、多重比較以及相關性分析,采用Origin9.0制圖.
3 結果與分析
3.1 香果樹幼苗的組成
武夷山香果樹種群幼苗相對較少,2015年11月份平均每個種群中僅有32.5株1 a幼苗存活,其中根萌苗約占38%,不同起源幼苗的具體年齡結構如圖1所示.由圖1可以看出,隨著年齡的增加,香果樹幼苗的數量持續下降,至5 a后種群數量逐漸穩定在2.3株左右,幼苗的密度約為56.2株/hm2,期間3 a、5 a和7 a實生苗缺失.
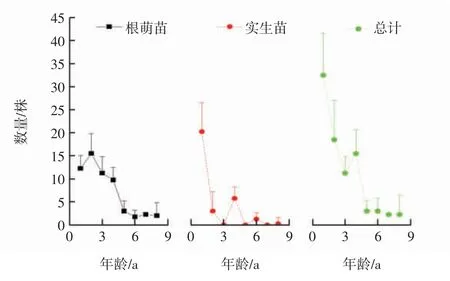
圖1 香果樹幼苗的年齡結構Fig.1 Age class structure of the saplings of E.henryi
3.2 香果樹幼苗的生命表及存活曲線
根據香果樹種群各齡級幼苗的數量編制生命表,結果如表2所示,繪制幼苗存活曲線結果如圖2所示.由表2中的死亡率和消失率可知,香果樹幼苗總體在3 a時出現負值,根萌苗在1 a和6 a時出現負值,表明此年齡段幼苗數量不足.香果樹幼苗介于Deevey-Ⅱ型和Deevey-Ⅲ型之間,經數學模型檢驗可知,香果樹幼苗總體和實生苗的存活曲線經冪函數擬合所得F值與R2均大于線性函數的數值,故兩者更接近于Deevey-Ⅲ型;根萌苗存活曲線的擬合方程中,冪函數擬合方程的F值較大而線性函數的R2較大,從而表明根萌苗的存活曲線介于Deevey-Ⅱ型和Deevey-Ⅲ型之間.
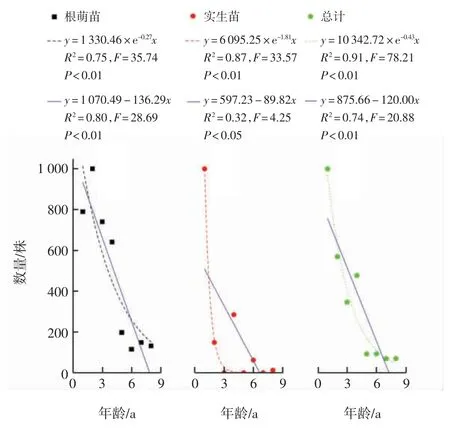
圖2 香果樹幼苗存活曲線Fig.2 Survival curve of the saplings of E.henryi
3.3 香果樹幼苗的生長特征
武夷山香果樹原生生境中根萌苗的生長較早,每年3月中旬開始展葉,而實生苗較晚,4月初香果樹種子露白萌發,4月中旬出現真葉.觀察測量實生苗和根萌苗的形態指標,結果如表3所示.由表3可以看出,香果樹根萌苗的高度、基徑、葉面積、比葉重均大于實生苗的數值,葉數小于實生苗的數值,兩種幼苗在1 a、2 a時的形態指標差距較小,隨著年齡增加差距逐漸增大.5個形態指標中,兩種幼苗的苗高差距最大,根萌苗苗高為實生苗的2~3倍.
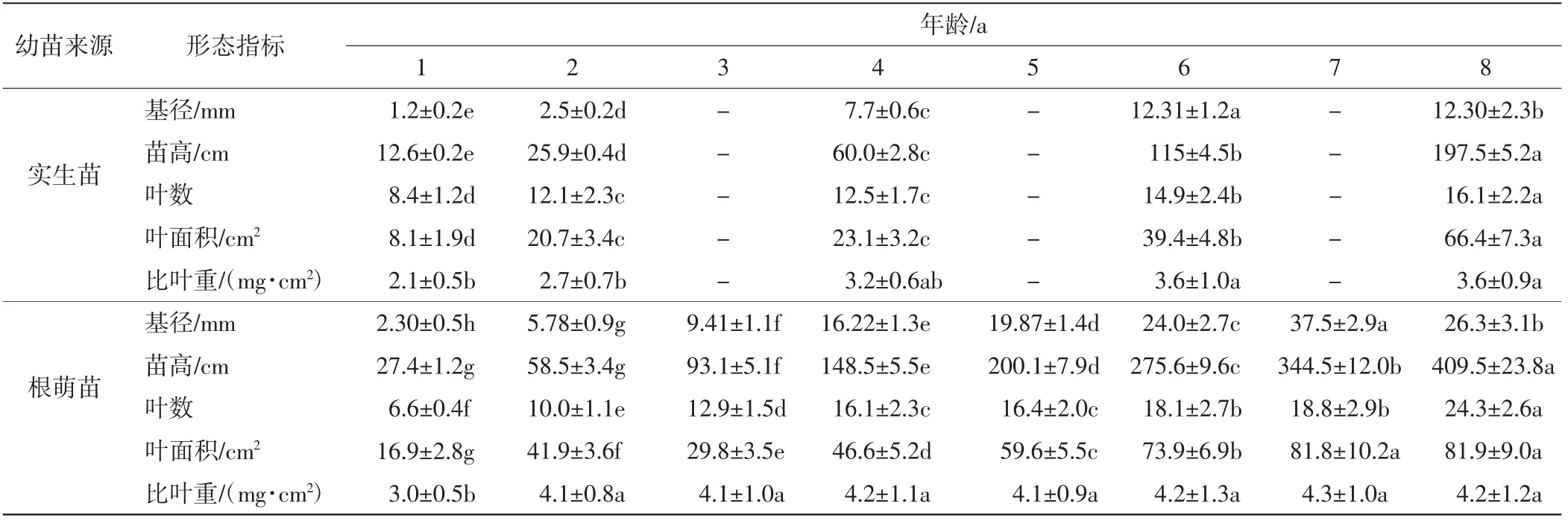
表3 香果樹幼苗生長特征Tab.3 Growth characteristics of the saplings of E.henryi
3.4 香果樹幼苗的耐陰性
對位于原生生境中不同位置的香果樹幼苗的新生側枝進行形態觀測,結果如表4所示.由表4可以看出,冠下幼苗新生側枝的基徑和葉厚最小,葉面積最大;位于冠緣幼苗新生側枝的枝長、節間長度、葉面積較大,葉厚相對較小;位于林窗的幼苗新生側枝的基徑、葉厚較大;同類型幼苗相比,位于林緣空地幼苗新生側枝的基徑、枝長和葉厚均最大,林緣空地下實生苗的葉片面積最小,而根萌苗的葉面積普遍高于實生苗.即相同微生境中實生苗的各項指標均顯著小于根萌苗,其中根萌苗的葉面積、節間長度以及枝長為實生苗的4~5倍,基徑和葉厚約為實生苗的1.4倍.
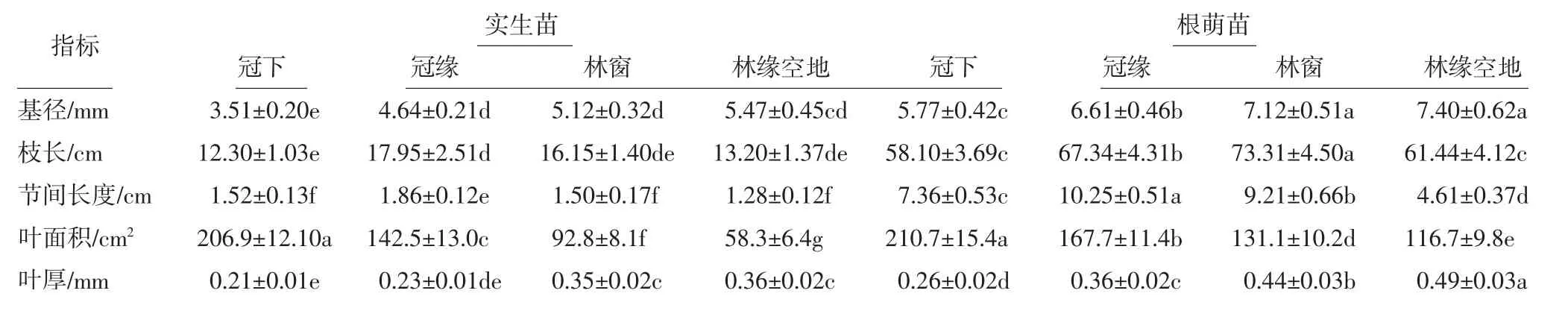
表4 香果樹新生苗側枝形態特征Tab.4 Morphological characteristics of annual branches of E.henryi
不同微生境中香果樹幼苗新生側枝上成熟葉片的各項生理指標差異較大,如表5所示.其中,冠下的幼苗葉片含水量和胞間CO2濃度顯著大于其他微生境中的數值,林緣空地處的幼苗葉片含水量最低,林窗下幼苗葉片的胞間CO2濃度最低;葉綠素含量、Ca/Cb、凈光合速率以及氣孔導度則以冠緣或林窗生境中最大,其他微生境較小;蒸騰速率與葉片含水量相反,冠下幼苗最低,林緣空地幼苗最高.香果樹根萌苗新生側枝成熟葉片的含水量均顯著小于同生境中實生苗的數值,除冠下幼苗外根萌苗的蒸騰速率均顯著大于實生苗的數值.
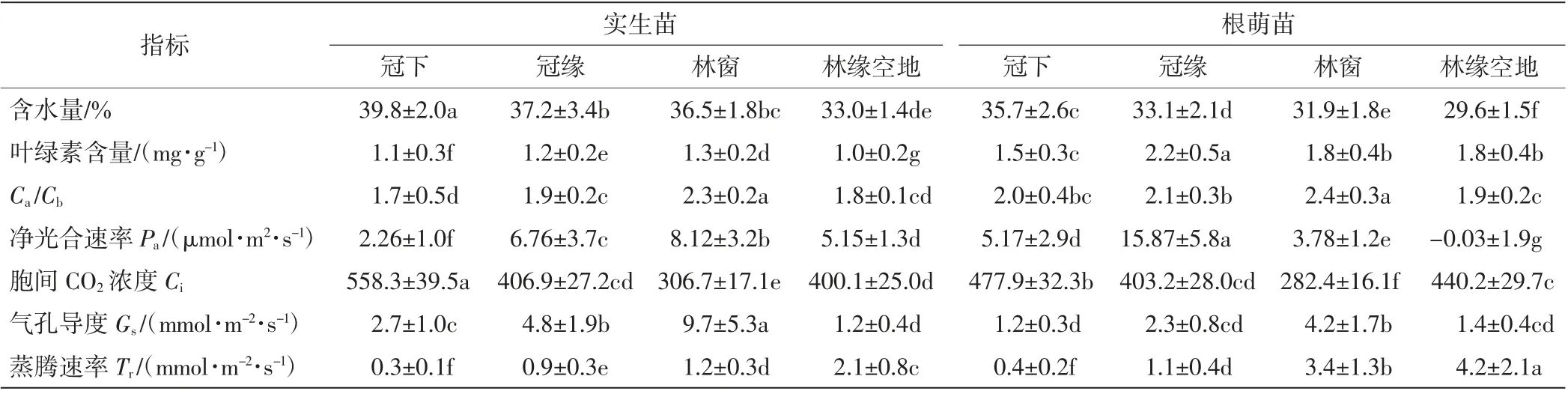
表5 香果樹新生苗側枝光合生理指標Tab.5 Photosynthetic physiological index of annual branches of E.henryi
4 討論與結論
4.1 香果樹根萌苗和實生苗的種群結構
分析種群的年齡結構是揭示種群動態變化和預測種群發展趨勢的重要手段[26],武夷山香果樹種群由于大多生長于溪流邊,林內存在大量礫石,幼苗很難存活,其存活曲線整體呈Deevey-Ⅲ型,種群呈衰退型[23].本研究發現香果樹幼苗較少,盡管1 a實生苗的數量較多,但大多數個體在生長過程中死亡,所調查的4個樣地中僅存活了1株8 a實生苗,3 a、5 a和7 a實生苗缺失,這是由于香果樹每2~4 a開花結果一次[27],其種子壽命極短,常溫下僅能存活10個月[21,28],次年存留于土壤中的種子無法萌發,導致實生苗的年齡不連續.根萌苗的數量較多,占幼苗總體的65.44%,1 a苗個體每個樣地約有12.25株,但8 a苗則減少到2株左右,由此可見香果樹幼苗數量極低,無法維持種群穩定,種群呈衰退型.擬合曲線顯示1~8 a香果樹幼苗的存活曲線更接近于Deevey-Ⅲ型,這表明其低齡幼苗死亡率極高.
4.2 香果樹根萌苗和實生苗的生長特性比較
有研究表明,在植物自然更新過程中,根萌苗比實生苗更具優勢[29-30],但也有學者認為某些植物(如喜光先鋒物種Cordia alliodora、Casearia gossypiosperma)的實生苗和根萌苗一樣具有較高的生長速率[31].本研究結果發現,香果樹根萌苗萌芽時間早,生長迅速,在基徑、苗高、葉數、葉面積、比葉重等方面均高于實生苗的數值,二者各年齡(1 a、2 a、4 a、6 a、8 a)的平均比值依次為2.45、2.37、1.10、1.85、1.32,且兩種幼苗的基徑、苗高和葉數隨著年齡增加差距逐漸增大,這表明香果樹的根萌苗通過獲取母樹的營養,在短時間內地上和地下部分功能發育完善,隨著年齡的增加,其競爭優勢更加突出.
4.3 香果樹幼苗自然更新能力和對樹蔭的忍耐力
自然界中通過種子進行更新的物種較少[31],這可能是由于種子被捕食率高、種子活力較低、實生苗死亡率高等原因導致.香果樹開花量大,結實較少,雖然果實中含種子數量較多,但由于種子微小、壽命極短、幼苗生長緩慢,導致種子活力、種子萌發過程及幼苗極易受到環境影響;而根萌苗生長迅速,抵抗力強,不易受環境影響[32].這使得利用根、莖的萌蘗力增加幼苗數量成為香果樹自然更新的重要途徑.本研究發現香果樹幼苗主要存在于4種微生境中,即樹冠下、冠緣下、林窗以及林緣空地中.通過對香果樹根萌苗及實生苗的新生側枝進行觀測,發現香果樹根萌苗在這4種微生境中的生長均優于實生苗,在相同微生境中兩種幼苗同一形態指標的數值差異均具有統計學意義,且冠下根萌苗側枝基徑和苗高的生長速率高于其他生境,這表明香果樹根萌苗比實生苗更適合生存于陰暗環境中.光合生理指標則顯示根萌苗葉片含水量低、蒸騰作用強、胞間CO2濃度低,且冠下的葉片葉綠素含量高,光合有效輻射高,這些均表明根萌苗在陰暗環境中利用光能的能力比實生苗強,在香果樹種群更新中優于實生苗.
4.4 香果樹自然更新建議
香果樹原生生境中根萌苗數量較多,在其自然更新中具有不可替代的作用.本研究發現根萌苗在冠緣和林窗下的生長顯著優于冠下和林緣空地,實生苗也有類似規律,這表明冠緣和林窗是香果樹幼苗生存較適宜的環境,這可能是因為光照及該微生境的水熱組合較有利于幼苗生存.林緣空地的幼苗節間較短,生長緩慢,長勢較差,根萌苗的光合有效輻射甚至為負值,表明光照太強會抑制幼苗的形態建成,不利于其光合作用,故應對此生境幼苗進行遮蔭處理.而冠下幼苗所處位置光照較弱,應清理地被物,增加透光率,以利于其進行光合作用,促進快速生長.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
