水稻已克隆抗稻瘟病基因的研究與應用
2022-11-03 10:32:24王小剛蘇思榮李曉蓉張銀霞
智慧農業導刊 2022年21期
關鍵詞:水稻
王小剛,蘇思榮,李曉蓉,張銀霞
(寧夏大學 農學院,銀川 750021)
稻瘟病不僅會導致水稻產量顯著下降,病害發生嚴重的情況下甚至是顆粒無收,而且還會影響稻米品質[1]。因此,防治稻瘟病,降低稻瘟病發病率對世界糧食安全具有重要意義。抗稻瘟病水稻新品種不斷地選育與合理種植可以有效地阻礙稻瘟病的發生,且持續對抗稻瘟病基因進行挖掘和利用對水稻抗稻瘟病育種具有非常重要的意義。據不完全統計,已有大約超過30個抗性基因被成功克隆[2]。本文通過整理已克隆的水稻抗稻瘟病基因,分析其在染色體上的分布,歸納其基因的分類,闡述其遺傳特性、抗病機制及抗病效果。最后對已克隆的水稻抗稻瘟病基因在水稻抗性育種中的應用進行了總結,以期為水稻抗稻瘟病育種提供理論依據。
1 水稻已克隆抗稻瘟病基因在染色體上的分布
從目前整理的已克隆基因結果來看,有30個屬于NBS-LRR蛋白,就是典型的Pi-b基因,且是第一個克隆出來的主效抗稻瘟病基因,其位于2號染色體上。Pi-9是第一個被克隆出來的廣譜抗稻瘟病基因。Pi-21是第一個被克隆出來的部分抗性基因。Bsr-d1為MYB轉錄因子,是在廣譜持久抗病材料地谷中發現并克隆出來的部分抗性基因。這些基因不均勻地分布在各條染色體上(圖1)。其中第11號、第6號染色體上分布較多,第5號、第7號染色體上暫時沒有發現已克隆的抗稻瘟病基因。由圖可知,在6號、9號、11號和12號染色體上,形成已克隆抗稻瘟病基因簇。
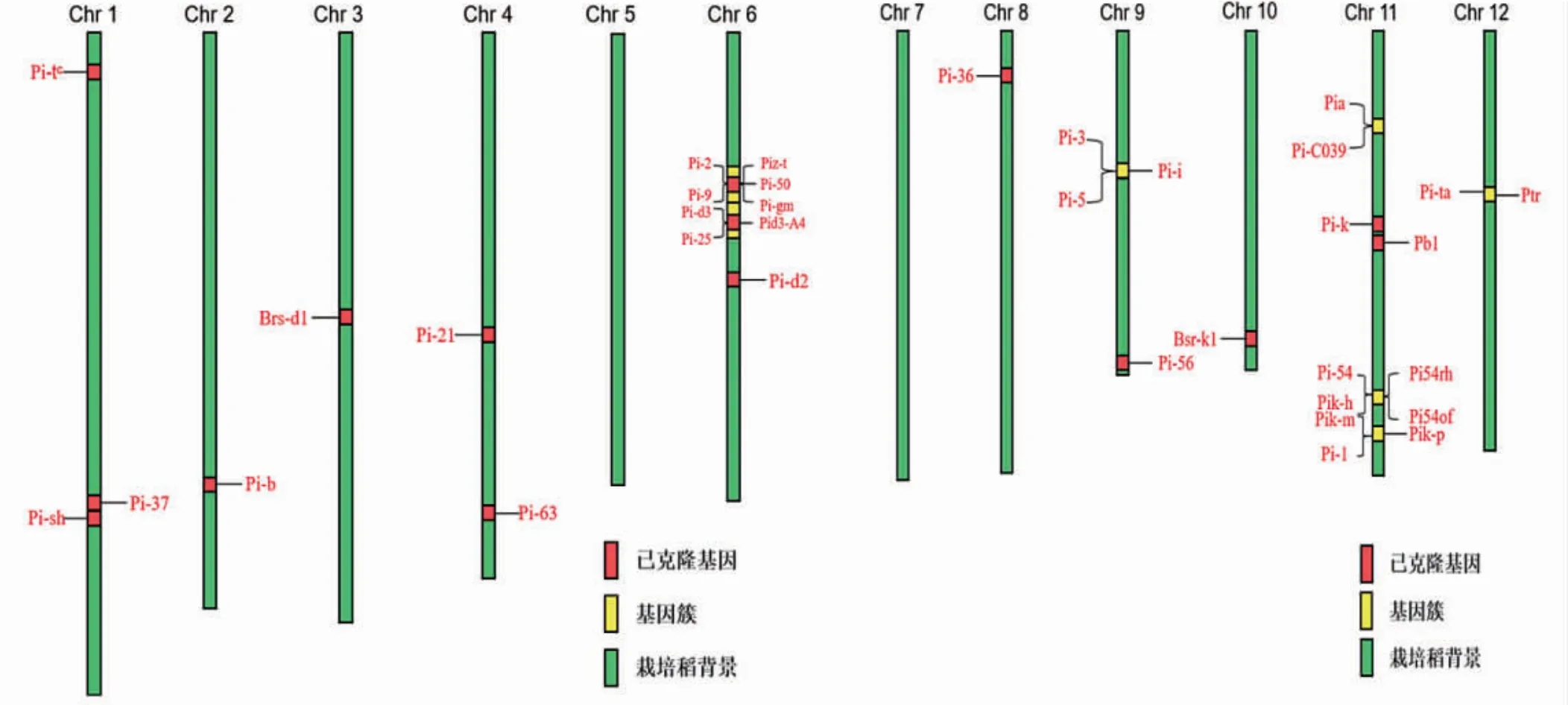
圖1 已克隆水稻抗稻瘟病基因在染色體上的分布
2 已克隆水稻抗稻瘟病基因的分類研究
已克隆抗稻瘟病基因由于編碼的蛋白質結構特征不同,其作用機理相應也不同,為了更清楚地研究抗性機理,本文對水稻抗稻瘟病基因進行了分類和分析。
根據不同的基因編碼蛋白將其分類。
2.1 核苷酸結合位點-富含亮氨酸重復序列蛋白(NBS-LRR)
Pi-b的NBS區包含3個保守結構、17個LRR區和8個半胱氨酸殘基,其呈簇狀分布。Pi-b可以通過溫度、光照和其他環境變化來誘導和調節;Pi-ta是第二個被克隆的抗稻瘟病基因,含有2個外顯子和1個1 463 bp的內含子,其與效應蛋白質AVR-Pita直接識別;Pi-9有17個不完整的富含亮氨酸重復,該基因在植株中組成型表達,不受稻瘟病侵染誘導。
2.2 受體蛋白激酶(RLK)
Pi-d2有825個氨基酸,其蛋白質與細胞外區特殊的膜外凝集素相互作用。23個疏水氨基酸組成跨膜螺旋結構,絲氨酸/蘇氨酸激酶構成細胞質區域。
2.3 富含脯氨酸結構域蛋白(PCP)
Pi-21由266個氨基酸組成,其C端包含富含脯氨酸域,N端具有蛋白互作結構域,參與水稻防衛反應,介導水稻的全生育期抗性。
2.4 富含ARM重復序列蛋白
Ptr是一個非典型的廣譜抗性蛋白,有4個Armadillo重復區,抗病性與Pi-ta有關,是單子葉植物所特有的基因。
2.5 富含四肽重復序列蛋白(TPRs)
新近發現并克隆的水稻稻瘟病抗性基因Bsr-k1是一個對稻瘟病和白葉枯病都具有廣譜抗性的隱性基因。Bsr-k1蛋白能與免疫應答相關的OsPAL基因家族的大部分mRNA結合,木質素合成增多,增強了水稻對稻瘟病菌的抗性。
3 水稻已克隆抗稻瘟病基因的遺傳特性
3.1 水稻已克隆抗稻瘟病基因的顯隱性
已克隆的抗稻瘟病基因大部分為顯性基因,例如Piz-t、Pi-z、Pi-2、Pi-9、Pig-m等均為顯性抗病基因[3]。極少數為隱性基因,例如Bsr-k1。隱性抗病基因結構保守及其作用機制的識別將有助于進一步了解抗病機理。
3.2 水稻已克隆抗稻瘟病基因的專化性
已克隆的抗稻瘟病基因大多表現小種專化,如Pid2對稻瘟病菌株ZB15表現為專一抗性;Pi-37對菌株CHL1340和CHL1159具有專化抗性。少數表現非小種專化,如Pi-21基因的近等位基因系AA-pi21對10種稻瘟病菌具有中等抗病性,屬于功能喪失性突變。
3.3 水稻已克隆抗稻瘟病基因的廣譜性
水稻廣譜抗稻瘟病基因大部分以基因簇的形式呈現。如第6號染色體上有抗性基因Pi-9、Pi-2等形成基因簇。其中,來自13個國家/地區的43個菌株,在四倍體小粒野生稻(Oryza minuta)中鑒定出的Pi-9基因都對其產生抗性。抗稻瘟病基因Pi-54/Pik-h、Pi-1、Pi-49在第11號染色體長臂上以基因簇分布所呈現。在魔王谷中鑒定的Pi-49對153個稻瘟菌株有抗性。抗稻瘟病基因Pi-tb、Pi-39、Pi-ta在第12號染色體短臂上以基因簇分布所呈現。Pi-ta對大部分來自歐洲的稻瘟病菌株有抗性。為了能夠縮短抗稻瘟病育種周期,也為了拓寬抗病基因的應用領域,就要不斷挖掘廣譜抗病基因。
4 水稻已克隆抗稻瘟病基因的抗性機制研究
水稻稻瘟病菌免疫機制的研究促進了水稻稻瘟病抗病機理的深入探討。通過分析發現水稻抗稻瘟病基因的抗病機制可以分為5類。
(1)R基因介導的稻瘟病完全抗性機制。不同的R基因表現出的抗稻瘟病特性有差異,部分R基因具有廣譜抗稻瘟病性;有些R基因抗譜相對較窄,對某些生理小種具有專化性。
(2)轉錄因子參與調控的稻瘟病部分抗性機制。目前,植物所特有的WRKY型轉錄因子通過轉錄激活或轉錄抑制,調控水稻抗性相關基因的表達,進而調控稻瘟病抗性。
(3)泛素連接酶參與調控的稻瘟病部分抗性機制。泛素連接酶可以直接與AVR蛋白互作,如APIP6,APIP10。當效應蛋白Avr-Piz-t進入水稻細胞后,會立即被APIP6和APIP10識別,促進Avr-Piz-t降解并阻止病原菌的侵入。
(4)磷酸激酶參與調控的稻瘟病部分抗性機制。調控蛋白的磷酸化狀態是稻瘟病菌侵染水稻免疫信號激活的關鍵。SDS2是RLK蛋白,其通過RLCK118將磷酸化修飾信號傳遞給NADPH氧化酶OsRbohB,促進植物細胞內ROS的迸發,加強了水稻抵御稻瘟病菌侵染的能力。
(5)其他非R基因參與調控的稻瘟病部分抗性機制。與植物激素合成相關蛋白,如合成酶OsACS2等。提高OsACS2的表達量,促進PR基因表達,提高水稻對多種稻瘟病菌生理小種的抗性,在抗病育種中具有較好的應用潛力[4]。
R基因介導的稻瘟病完全抗性已廣泛應用于水稻育種和生產中。雖然非R基因介導的部分抗性相對較弱,但其抗性廣譜,抗性持久穩定。
5 水稻已克隆抗稻瘟病基因的應用研究
近年來,有學者研究發現多基因聚合有助于提高水稻品種抗性的持久性和廣譜性[5]。劉士平等[6]研究發現,多個抗稻瘟病基因聚合后,相關性極顯著的抗性基因均有互相作用的表現,這樣的表現結果能夠抵抗單個抗病基因不能抵抗的生理小種。目前,已報道的利用抗稻瘟病基因聚合提高其抗性的研究見表1。其中,陳志偉等[7-8]將Pi-1、Pi-9和Pi-kh基因導入到福恢673和N175中,分別選育出10個福恢673的近等基因系和16個N175的近等基因系。用福恢673近等基因系Line9配組的雜交稻新組合兩優7283和金泰優683在區試中均表現出產量高、抗病強及生育期適宜等特點。姚姝等[9]將Pi-ta、Pi-b和Wx-mq基因聚合一體,最終選育出了一個水稻新品種“南粳0051”,其不僅高產優質,而且還表現為抗稻瘟病。馬作斌等[10]利用水稻聚合雜交的方法,在后代材料中鑒定出了一個穩定株系A-3,而且發現該株系同時含有Pi-ta和Pi-b 2個抗性基因,后被遼寧省品種審定委員會審定且命名為“鐵粳16”。
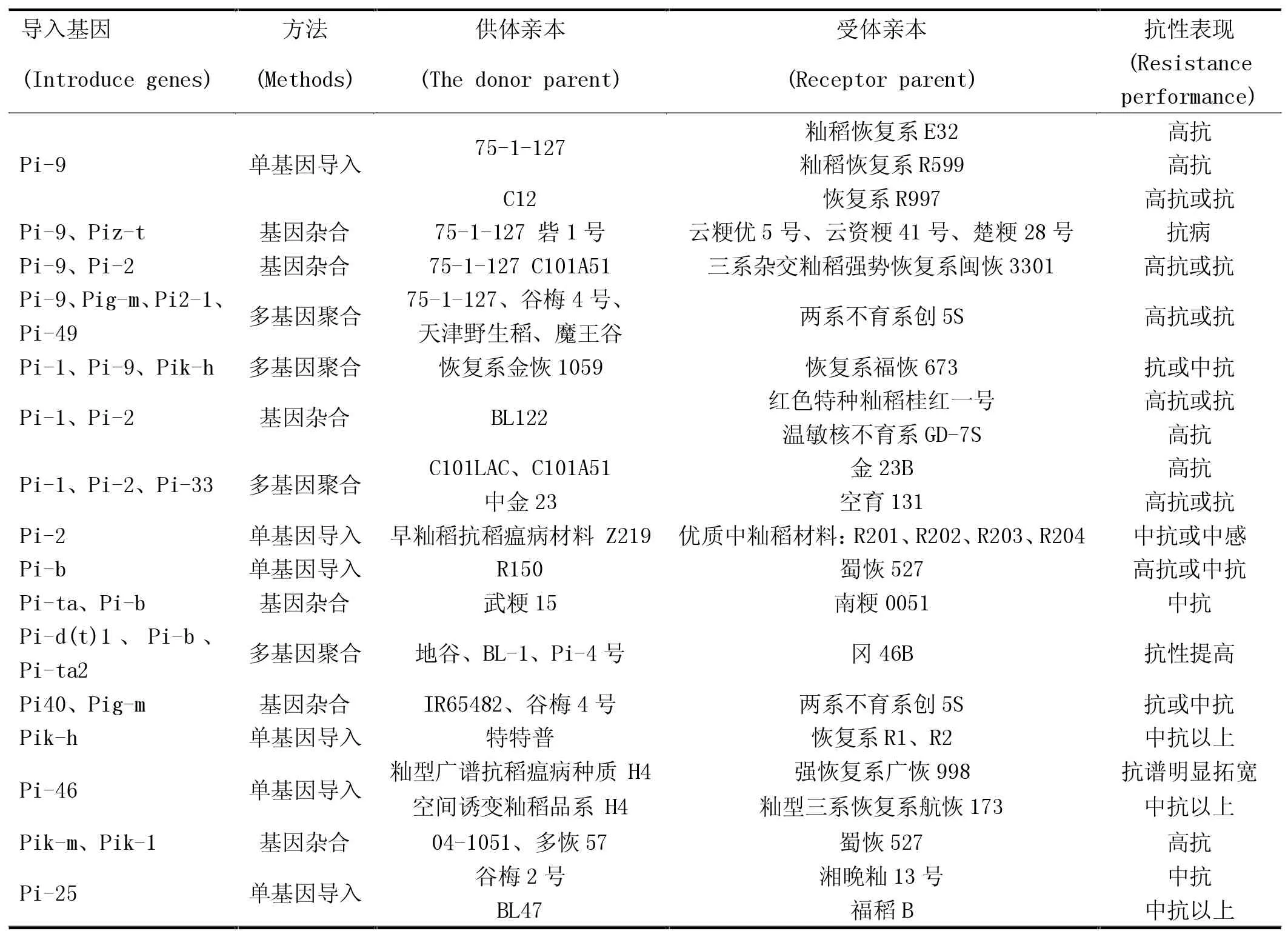
表1 水稻已克隆抗稻瘟病基因的應用
6 結束語
以上研究表明,稻瘟病菌和稻瘟病抗性基因共同進化,然而含有單個抗性基因的材料經過多年種植后其抗性逐年下降,但是聚合多個已克隆且廣譜稻瘟病抗性基因的品種,在大規模應用推廣以后,抗性表現明顯提高,而且延伸了抗病的時效性;但是,還可能產生針對多個抗性基因的超級稻瘟病小種。因此,注重調查當地的稻瘟病菌優勢生理小種,及時發出稻瘟病菌流行預報,改種或者輪種含有新的已克隆抗稻瘟病基因的水稻品種,避免水稻品種的稻瘟病抗性迅速喪失,延長稻瘟病抗性基因的水稻品種使用壽命。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
