科學史視角下“基因在染色體上”的探索過程?
2022-11-22 08:12:14王玉龍廣東實驗中學廣東廣州510055
生物學通報 2022年1期
關鍵詞:實驗
王玉龍 (廣東實驗中學 廣東廣州 510055)
在學習了摩爾根(Morgan)紅眼、白眼果蠅雜交實驗后,學生對基因在染色體上的結論將信將疑。教師進一步解釋布里吉斯(Bridges)的果蠅雜交實驗中出現的初級例外情況后,學生逐漸接受了基因在染色體上的觀點,但很難想到染色體變異的方向,對布里吉斯本人為何能提出相應的假說解釋依然充滿疑惑。對上述疑惑的解答需要從基因在染色體上(染色體遺傳)的探索過程講起。
1 基因與染色體的關系假說
1883—1892年,魏斯曼(Weismann)所提出的“種質論”認為,生物可分為“種質”和“體質”2 個部分。體質由種質發育而來,在世代之間不連續,種質在世代之間是連續的。雖然該理論之后被證明是錯誤的,但有力駁斥了“用進廢退”和“獲得性狀遺傳”的觀點。魏斯曼結合受精現象的觀察認為,遺傳的物質基礎存在于細胞核內部,并預測了遺傳物質在細胞核內某種載體之上,為染色體-DNA(核酸)-基因關系的確立奠定了基礎。
1900年,3位科學家幾乎同時重新發現了孟德爾(Mendel)的劃時代論文《植物雜交的試驗》,總結了孟德爾遺傳理論的精華,可概括為“顆粒遺傳學說”。該學說后面逐漸取代了達爾文(Darwin)堅持的“泛生論”和魏斯曼提出的“種質論”的遺傳學說,1909年約翰遜(Johannsen)將“顆粒遺傳學說”中的“顆粒”命名為基因[1]。
基因究竟在哪里?鮑維里(Boveri)以海膽為材料,取出卵細胞的細胞核,植入精子,進一步處理得到2 個精子受精的“受精卵”,通過研究其發育情況,證明了細胞核在遺傳中的決定作用,同時發現不同的染色體有不同的性質。鮑維里還根據其對蛔蟲的研究,發現染色體具有遺傳的連續性和獨立性,認為受精卵分裂形成的細胞,其中的染色體一半來自父方、一半來自母方。1903年,薩頓(Sutton)通過研究蝗蟲生殖細胞的形成過程,發現了基因和染色體存在明顯的平行關系,進而推測基因在染色體上。1905年,威爾遜(Wilson)認為生物體的性狀多于染色體數目,若基因和染色體有關,則一條染色體上含有多個基因,即會存在基因連鎖。
在摩爾根研究果蠅之前,關于基因和染色體的關系假說已存在,只是缺少相應的實驗證據予以證明。但摩爾根本人的學術研究經歷使其并不認同孟德爾遺傳學說,他曾用家鼠與野鼠做雜交實驗,結果發現毛色性狀的遺傳毫無規律,使其對孟德爾學說產生了懷疑;當陸續有更多的動物遺傳實驗難以用孟德爾學說解釋時,也就對此學說有了根深蒂固的成見[2]。
2 性染色體的發現
1891年,亨金(Henking)發現昆蟲精巢細胞中含有一個不同的染色體,并將其命名為X 染色體。1901年,麥克隆(McClung)提出X 染色體與性別有關,但錯誤地認為X 染色體決定雄性[3]。1905年史蒂文斯(Stevens)首次提出甲殼蟲的Y 染色體與雄性有關,雄性為XY,雌性為XX。但當其研究果蠅裝片時,由于裝片制作效果不理想,并未發現Y染色體,認為果蠅的雌性為XX,雄性只有一條X染色體[3]。性染色體及性別決定的發現,為后續伴性遺傳的研究奠定了基礎。
3 摩爾根果蠅雜交實驗
1908年,摩爾根建議其學生佩恩(Payne)驗證拉馬克(Lamarck)“用進廢退”學說的正確性,佩恩以果蠅為材料,在暗室中飼養,觀察其是否會因為不需要眼睛而產生無眼后代。結果發現一直飼養至69 代時,均未出現無眼果蠅,進而證明了“用進廢退”學說的不合理性。摩爾根也由此發現了果蠅是理想的遺傳學實驗材料,便創建了“蠅室”,飼養果蠅進行遺傳學實驗。1910年5月,摩爾根偶然在蠅室內野生型果蠅中,發現了一只白眼雄果蠅,他馬上意識到該果蠅的重要性,在其死亡之前完成了與紅眼雌果蠅的雜交。所得F1代中有1 237 只紅眼果蠅和3 只白眼果蠅,摩爾根認為這3 個例外情況是進一步發生突變導致的,未對其進行討論。繼續將F1的紅眼果蠅相互交配,所得F2代中有2 459 只紅眼雌果蠅,1 011 只紅眼雄果蠅,782 只白眼雄果蠅。摩爾根在上述實驗的基礎上,繼續進行回交實驗,讓F1的紅眼雌果蠅和最初出現的白眼雄果蠅雜交,結果出現紅眼雌性、紅眼雄性、白眼雌性和白眼雄性4 種表型,比例接近1∶1∶1∶1[5]。
通過分析數據可知,紅眼果蠅與白眼果蠅的比例并非3∶1,但摩爾根認為考慮到白眼果蠅存活率較低的原因,可認為該比值為3∶1,但白眼果蠅僅限于雄性,這一點與孟德爾實驗并不相同。摩爾根雖然對孟德爾學說抱有很大成見,但在事實面前,摩爾根還是以顆粒遺傳學說作為解釋。由于當時并未發現果蠅中有Y 染色體,加之摩爾根對基因在染色體上的學說也有很大疑慮,所以其解釋過程并非如高中《生物》教材中所介紹的那般,更不可能提出如同很多高中生物學教師所講的“白眼基因位于X 染色體的非同源區段,位于X、Y 同源區段,位于Y 的非同源區段”等假說。而是使用了如下解釋過程,如圖1所示。

圖1 1910年摩爾根論文中的遺傳圖解[5]
在上述解釋中,R代表紅眼“因子”(基因),W代表白眼“因子”,雌果蠅有2 條X 染色體,雄果蠅只有1 條X 染色體。摩爾根將親本白眼雄果蠅表示為WWX,紅眼雌果蠅表示為RRXX,白眼雄果蠅產生的精子中均含有W,但一半的精子含有X,一半無X 染色體。紅眼雌果蠅產生的配子全部含有R因子和X 染色體,摩爾根認為R與X 染色體緊密偶聯,F1的紅眼雄果蠅產生的配子為RX和W2 種(不能產生WX和R的配子)。可見摩爾根十分謹慎,并未提出R在X 染色體上、Y 染色體上沒有R的假說。
為驗證自己的解釋,摩爾根還設計了4 組雜交實驗(其中有一組與回交實驗相同,等于是3 組雜交實驗)。第1 組用白眼雌果蠅和白眼雄果蠅雜交,其產生的雄配子應為WX、W,雌配子為WX,雜交子代應全為白眼果蠅,如圖2所示。

圖2 摩爾根對白眼雌果蠅與白眼雄果蠅的雜交解釋[5]
第2 組實驗用F2的紅眼雌果蠅與白眼雄果蠅雜交,由于F2的紅眼雌果蠅應該有RRXX和RWXX2 種類型,故雜交結果應該為:有些雜交組合子代應全為紅眼,有些雜交組合應出現比例相等的紅眼雌性、紅眼雄性、白眼雌性和白眼雄性4 種類型。如圖3所示。

圖3 摩爾根對F2 紅眼雌果蠅與白眼雄果蠅的雜交解釋[5]
第3 組實驗用白眼雌果蠅與F1紅眼雄果蠅雜交,子代應出現等比例的紅眼雌果蠅和白眼雄果蠅,如圖4所示。

圖4 摩爾根對白眼雌果蠅與F1 紅眼雄果蠅的雜交解釋[5]
上述幾組雜交實驗結果與預期完全吻合,進而說明了摩爾根假說的合理性。高中《生物》教材中直接用紅眼基因位于X 染色體上的假說解釋此實驗現象,當然事實上確實如此,作此解釋也更為科學,但卻超出了摩爾根彼時的認識,當時的摩爾根并未接受基因位于染色體上的學說。后續的若干研究成果提供了越來越清晰的實驗證據,摩爾根才接受了基因在染色體上的事實。
4 基因在染色體上線性排列的遺傳圖譜
自摩爾根的白眼雄果蠅之后,又陸續發現了黃體、朱砂眼等突變型果蠅,且發現這些性狀與性染色體緊密偶聯。摩爾根用黃體紅眼雌果蠅與灰體白眼雄果蠅雜交,F1中雄蠅全是黃體紅眼、雌蠅全是灰體紅眼,F1相互交配得F2,F2中的雄蠅表現為黃體紅眼∶灰體白眼∶灰體紅眼∶黃體白眼=99∶99∶1∶1,F2中的雌蠅表現為黃體紅眼∶灰體紅眼=1∶1。經分析可知F2中雄蠅的重組率為1%,用F2中的黃體白眼雄果蠅與Fl中的灰體紅眼雌果蠅(雙重雜合子)交配,后代不論雌、雄均表現出了重組性狀,重組率均是1%。1911年,摩爾根對此現象所作的解釋是:上述2 對“因子”均位于性染色體上,即表現為“相引”。“相引”的因子在染色體上呈線性排列,減數分裂時同源染色體聯會并發生交叉互換。1911年摩爾根的學生斯特提文特(Sturtevant)想到在染色體上線性排列的基因,可通過子代重組的概率推算出2 個基因之間的距離,并根據此想法,繪制了第1 張染色體的遺傳圖譜[2],如圖5所示。

圖5 斯特提文特確定的第1 個遺傳圖譜[2]
5 連鎖群
摩爾根依據基因在染色體上線性排列的關系,提出了“連鎖群”的概念(位于同一染色體上,相互之間存在連鎖遺傳關系的基因群)。1911年,發現了果蠅有3 個連鎖群,摩爾根由果蠅的染色體組成預測果蠅應有4 個連鎖群。1914年穆勒(Muller)發現了果蠅的卷曲翅基因,與無眼基因及此后發現的無剛毛胸基因構成了果蠅的第Ⅳ連鎖群[6]。
基于上述實驗現象,很多生物學家接受了基因在染色體上的學術觀點,但仍有一些人持懷疑態度,包括摩爾根本人也有所擔心,所以,需要更為直接的實驗證據證明。
6 布里吉斯的果蠅雜交實驗
布里吉斯、斯特提文特和穆勒是摩爾根“蠅室”的3 個重要成員,1913年布里吉斯重復了摩爾根的白眼雌果蠅與紅眼雄果蠅雜交實驗,經科學和耐心的統計,獲得了20 496 只子代,其中有10 735 只紅眼雌果蠅,9 849 只白眼雄果蠅,但同時有極少數例外情況,即有5 只白眼雌果蠅,7 只紅眼雄果蠅(之后稱此2 種例外為“初級例外”)。
按照摩爾根的學說,子代雄果蠅應全部為白眼,雌果蠅應全部為紅眼,不應出現該例外的情況。布里吉斯通過對果蠅的染色體觀察,已知X染色體與常染色體的比值為1 是雌性,比值為1/2是雄性,即雌性為XX,雄性為X,當時并未發現Y 染色體的存在。通過分析可知紅眼對白眼為顯性,結合之前紅眼基因在性染色體上的觀點,布里吉斯在解釋該例外白眼雌果蠅時,認為其2 條Xw染色體應該均來自于母本白眼雌果蠅,造成該現象的原因是母本雌果蠅在減數分裂時2 條X 染色體未分離,形成了XwXw的卵細胞,該卵細胞與正常減數分裂產生的無X 染色體的精子結合,形成了XwXw白眼雌果蠅。例外的紅眼雄果蠅其XW的染色體應來自于父方的XW,該精子與1 個不含X 染色體的卵細胞結合,形成了XW的紅眼雄果蠅,而不含X 染色體的卵細胞產生的原因,同樣是由于母本雌果蠅在減數分裂時2 條X 染色體未分離導致的,故以上2 種初級例外情況的成因是相同的,出現的概率也是一致的。
1914年,布里吉斯通過更仔細的細胞學觀察發現,雄果蠅中還含有1 個Y 染色體,但Y 染色體不決定性別,只決定雄果蠅的可育性,性別取決于X 染色體的數量,即XX、XXY 為雌性,X、XY、XYY 為雄性,XXX 的個體致死,XO 雄果蠅不育。進而指出解釋關于初級例外的表述中親本所產生的不含X 的精子應改為含Y 的精子,推測初級例外的白眼雌果蠅應為XwXwY,并指出該果蠅減數分裂可形成的配子有:XwY、Xw、XwXw、Y,通過細胞學檢查發現確實如此[7]。
布里吉斯為進一步驗證上述學說,將“初級例外”中的白眼雌果蠅(XwXwY)和紅眼雄果蠅(XWY)進行雜交,并預測該雜交的子代中除了有白眼雄果蠅和紅眼雌果蠅之外,還將出現少量的“次級例外”,即白眼雌果蠅(XwXwY)和紅眼雄果蠅(XWY),同時紅眼雌果蠅中有XWXwY、XWXw2 種類型,且比例為1∶1,白眼雄果蠅中有XwYY、XwY2 種類型,且比例為1∶1,如圖6所示[8]。實驗結果表明該雜交的子代中,出現了約4.3%的次級例外,該例外是野生型染色體組成的白眼雌果蠅和紅眼雄果蠅雜交所不會產生的,通過對子代果蠅的細胞學觀察,也證實了確實有XYY 和XXY 的染色體組成。
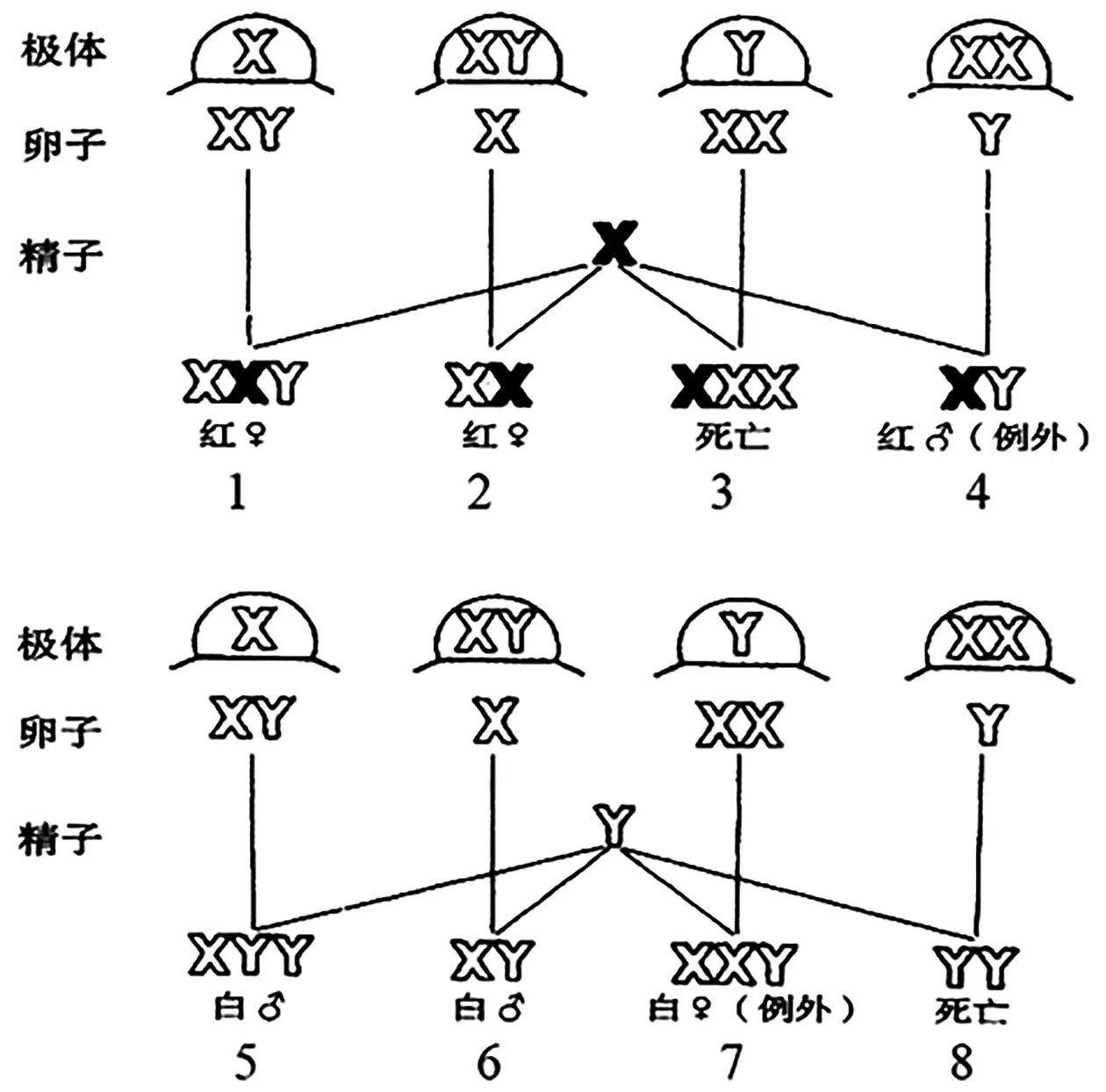
圖6 白眼XwXwY 雌果蠅與紅眼XWY 雄果蠅雜交[8]
布里吉斯發現的初級例外和次級例外情況,在性染色體上也出現例外,即染色體的行為、性染色體連鎖基因的行為、性別決定三者之間完全吻合,這種吻合是很難用偶然巧合解釋的,故由此推斷性連鎖基因位于X 染色體上。
7 黃身雌果蠅與灰身雄果蠅的雜交實驗
1922年,摩爾根用純合黃身雌果蠅與灰身雄果蠅雜交(當時已知灰身對黃身為顯性,且均與X染色體相偶聯),所產生的F1為黃身雌果蠅與灰身雄果蠅。該結果是之前的學說所不能解釋的,通過觀察親本果蠅的染色體組成發現,黃身雌果蠅的X 染色體是由2 條X 染色體相連形成的,同時還有1 條Y 染色體,故其染色體組成可表示為XXY,如圖7所示[9]。該果蠅減數分裂所產生的卵細胞為Y和XaXa,灰身雄果蠅產生的精子為XA、Y,故雜交產生的子代有XaXaY、XAY、XAXaXa(致死)、YY(致死),進而解釋上述雜交實驗現象。該實驗表明黃身基因隨X 染色體而遺傳,基因和染色體的行為又是高度吻合的,進而表明基因在染色體上。

圖7 摩爾根所觀察的變異黃身雌果蠅的染色體組成[9]
8 染色體的細胞學圖像
1928年,蘇聯科學家杜布贊斯基(Dobzhnsky)在摩爾根實驗室用X 射線處理果蠅,讓其發生染色體變異,通過對果蠅的裝片觀察,發現了3 號染色體的一段易位至較小的4號染色體上。杜布贊斯基通過對染色體斷裂點的研究分析,繪制出果蠅的3 號染色體的細胞學圖像,如圖8所示[10]。將該圖與基于遺傳學實驗數據而繪制的基因連鎖圖比較,發現二者是完全吻合的,進而為基因在染色體上提供了較為直接的證據。
9 果蠅的唾液腺多線染色體
1928年,德國科學家海茨(Heitz)改進了細胞學染色方法,發現染色體的某些部分染色更密集,即染色體會呈現出深淺不同的條帶或橫紋。1930年,海茨與鮑爾(Bauer)一起合作研究了毛蚊的唾液腺染色體,觀察到繩索狀結構,并認為是“巨型染色體”。該結果使遺傳學家佩因特(Paimter)很受啟發,意識到果蠅唾液腺染色體上的條帶可用以研究染色體變異中的斷裂點,也能作為定位基因在染色體上的坐標,并繪制了果蠅的染色體圖(圖9)[11]。佩因特發現染色體條帶密集處其所含的基因數量也較多(圖10)[12],并通過遺傳學實驗發現,基因的連鎖圖發生逆轉時,染色體條帶也發生逆轉,遺傳上表現易位或缺失時,染色體條帶也發生易位或缺失。布里吉斯也以果蠅的唾液腺染色體為材料進行了研究,繪制了果蠅減數分裂中期的染色體圖像與唾液腺染色體的對比圖像,發現在相同放大倍數下,減數分裂中期的4 號染色體是黑點狀,而果蠅唾液腺中的4 號染色體卻能看到很多細小條紋(圖11)[13]。由此意識到果蠅的唾液腺染色體是研究基因的物質基礎的理想材料,并得出了果蠅唾液腺染色體上的基因連鎖圖,如圖12所示[14],該圖譜成為了科學家的參考標準。果蠅唾液腺多線染色體的發現,為基因在染色體上提供了最為直觀和不可辯駁的證據,在物理層面上直接驗證了該結論。

圖9 佩因特繪制的果蠅唾液腺多線染色體[11](局部)

圖10 佩因特繪制的X 染色體上基因的連鎖圖[12](局部)
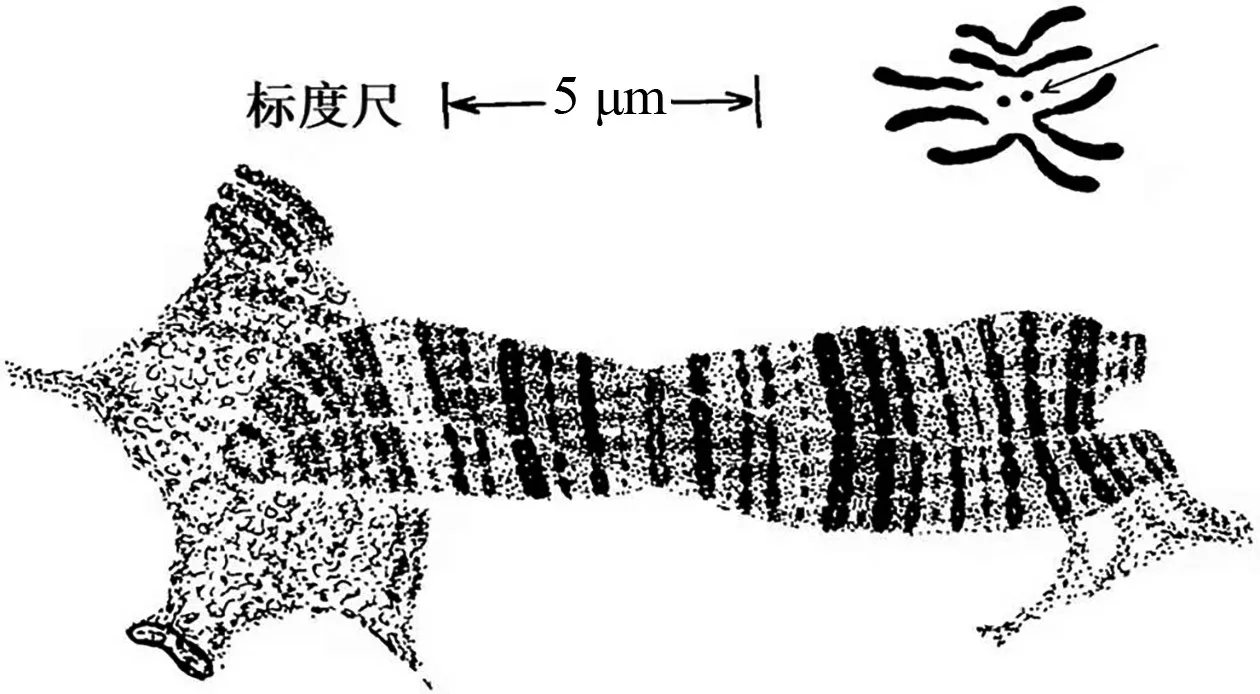
圖11 果蠅唾液腺染色體(下部條紋狀結構)

圖12 布里吉斯繪制的果蠅唾液腺染色體的基因連鎖圖[14]
果蠅是一個經典的遺傳學材料,摩爾根及其“蠅室”的眾多成員充分挖掘和利用了果蠅的研究價值。隨著科學研究的進行,呈現了越來越多的實驗證據,均共同指向了基因在染色體上。但由于基因的微觀性及“不可視”性,一開始包括貝特森(Bateson)在內的眾多知名科學家,對基因在染色體上的染色體遺傳學說表示質疑,甚至包括摩爾根本人當時也不敢肯定基因在染色體上。1933年摩爾根被授予諾貝爾獎時,當年他并未去領獎,不禁讓人猜測是因為他所采用的證明方法正是其當年所嘲笑過的孟德爾方法[2]。隨著越來越多、越來越直接的證據表明基因在染色體上,一些科學家才開始轉變態度,逐漸接受了基因在染色體上,最終使遺傳學的研究轉移至基因和染色體的方向上。
古人云“智者見于未萌,愚者暗于成事”,在染色體遺傳的發展過程中,正是摩爾根、布里吉斯、斯特提文特、杜布贊斯基、佩因特等科學家,將肉眼難以看見的基因通過各種實驗,以現象、數據、圖像等形式逐漸呈現,進而一步步勾勒和構建出了染色體遺傳的理論體系。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
