茴香醛對變異鏈球菌的抗菌活性和抗生物被膜活性
2022-11-29 14:14:36倪雅麗
熱帶生物學報 2022年6期
關鍵詞:生物
云 莉,倪雅麗
(海南省第二人民醫院 藥學部,海南 五指山市 572299)
齲齒是一種慢性進行性破壞性疾病,是由于口腔內共生菌和病原菌之間的生態失調而引起的。牙齒形成生物被膜(牙菌斑)后,牙齒殘留的碳水化合物和糖經生物被膜內的細菌分解產生酸,導致牙齒表面的牙釉質脫落。生物被膜是由微生物群落在牙齒表面形成的生態環境,由細菌進行代謝產生的蛋白質、多糖和核酸組成的胞外聚合物(EPS)基質構成[1?2]。 變異鏈球菌(Streptococcus mutans)是口腔中常見的一種革蘭氏陽性細菌,能引起口腔內微生物共生生態系統的失調。因此,變異鏈球菌也被認為是口腔內微生物形成生物被膜緊密相關的細菌之一[3]。變異鏈球菌通過復雜的生命活動定植于牙齒表面并形成生物被膜[4?5]。變異鏈球菌可以攝取口腔內殘存的各種碳水化合物進行代謝活動,產生有機酸不斷腐蝕口腔牙齒,導致齲齒的形成。變異鏈球菌代謝產酸是形成生物被膜中主要的毒力因素之一[6?7]。生物被膜表型在生理和功能上與浮游細菌有很大不同,生物被膜中的細菌表現出較低的代謝活性和生理機能,生物被膜結構可以作為細菌的物理屏障,限制抗生素滲透到生物被膜的深層。因此,在生物被膜中生長的細菌會增加其對抗生素的耐受性[8?9]。由于抗生素的濫用,導致常規療法下細菌對抗生素的耐藥性迅速增加,因此,迫切需要開發新型抗菌劑以抑制生物被膜的形成和降解成熟生物被膜。
近年來,使用天然植物產品治療口腔疾病日益受到關注并被廣泛研究[10]。茴香醛(p-methoxybezaldehyde)是廣泛存在的一類天然產物,主要是從茴香、小茴香和大蒜中分離得到,目前已被廣泛用于制藥行業的抗菌藥物生產[11?12]。《本草匯言》中記載,小茴香乃溫中快氣之藥。最近研究結果表明,小茴香中主要成分之一茴香醛對許多微生物表現出很強的抗菌活性,包括金黃色葡萄球菌、念珠菌和釀酒酵母[13?15],在化妝品和藥物的制備中也發揮著重要的作用。張冠楠等[16]的研究結果表明,茴香醛能改變金黃色葡萄球菌的菌體形態、細菌細胞膜的通透性;CHE等[17]的研究結果表明,茴香醛能改變細菌細胞壁完整性和細胞膜通透性發揮抑菌效果。然而,關于茴香醛對口腔致病菌中的變異鏈球菌抗菌活性尚未見報道。本研究旨在探討茴香醛對口腔致病菌變異鏈球菌的抗菌效果及潛在的作用機制,以期將茴香醛開發為抗齲齒的天然藥物。
1 材料與方法
1.1 菌種和試劑變異鏈球菌(S. mutans,ATCC25175)購自廣東省微生物菌種保藏中心;茴香醛(貨號A108958-5ml,阿拉丁)購自阿拉丁試劑(上海)有限公司;變異鏈球菌在腦心浸液肉湯(BHI,中國北京陸橋技術有限公司)培養基中培養;Live/Dead BacLight Kit試 劑 盒(ThermoFisher,L7012)。
1.2 最低抑制濃度檢測采用GRENIER等[18]的二倍稀釋法,將128 g·L?1茴香醛溶液和2.56 g·L?1洗必泰溶液稀釋成系列溶液,測定茴香醛和洗必泰對變異鏈球菌的最小抑菌濃度(MIC)和最小殺菌濃度 (MBC)。變異鏈球菌培養至對數生長期后,無菌BHI 液體培養基稀釋至2×105cfu·mL?1,取100 μL稀釋菌液加至含有不同濃度茴香醛的96孔板中,使孔中總體積為200 μL,細菌終濃度1×105cfu·mL?1。菌液+PBS為陰性對照組,BHI液體培養基+PBS為空白對照組,洗必泰處理孔為陽性對照組。在37 ℃培養箱中培養24 h后觀察不到混濁度的孔為茴香醛對變異鏈球菌的最低抑制濃度。
1.3 最小殺菌濃度測定將1.2實驗組中未出現渾濁度的孔取100 μL涂布于BHI固體培養基上,在37 ℃培養箱中培養24 h后觀察平板菌落生長情況,平板上無菌落形成視為茴香醛對變異鏈球菌完全殺滅,此濃度為最低殺菌濃度。每組設置3個復孔。
1.4 時間殺菌曲線實驗根據CHEN等[13]的殺菌動力學測定方法,變異鏈球菌培養至對數生長期后,無菌BHI液體培養基稀釋為2×106cfu·mL?1的菌懸液。稀釋菌懸液與不同濃度的茴香醛(終濃度為0.5×MIC,1×MIC和2×MIC)等體積混勻,置于37 ℃恒溫水浴0、10 、20 、30 、40 、60、90 、120、180 min。0.2%洗必泰(CHX)為陽性對照,無菌PBS+菌液作為陰性對照。孵育至指定時間后,4 ℃下6 000 r·min?1離心10 min,棄上清液,使用無菌BHI重懸,根據需要稀釋10倍,取50 μL涂布于無菌BHI固體培養基上,在37 ℃ 溫育24 h后計算細菌菌落。實驗重復3次并取平均值。
1.5 細胞膜完整性實驗
1.5.1 DNA 和 RNA泄露檢測參考WANG等[19]的方法,通過檢測細菌核酸的釋放量判斷細菌細胞膜的完整性。將變異鏈球菌在37 ℃培養至對數生長期,無菌PBS稀釋至2×108cfu·mL?1,分別以終濃度1 ×,2×和4× MIC的茴香醛處理,細菌終濃度為1×108cfu·mL?1。空白對照組以PBS處理。37 ℃下孵育0、1、2和4 h后, 4 ℃下6 000 r·min?1離心10 min。取上清液,OD260下測定吸光值。在進行DNA和RNA實驗的同時,采用平板涂布法檢測活細菌數目,方法同“1.4 時間殺菌曲線實驗”平板涂布部分,實驗重復3次并取平均值。
1.5.2 對細菌可溶性蛋白質泄露的影響細胞完整性可以通過測定上清液中細菌蛋白質的釋放量來確定,上清液中蛋白質含量測定參考BRADFORD等[19]實驗方法。變異鏈球菌培養至對數生長期(濃 度 為1× 108cfu·mL?1)用 茴 香 醛(1×MIC和2×MIC)處理。以PBS處理為空白對照。實驗組和處理組均在37 ℃下孵育0、1、2和4 h,4 ℃下2 500 r·min?1離心5 min,取上清液,OD595下測定吸光值。實驗重復3次并取平均值。
1.5.3 激光共聚焦掃描顯微鏡實驗參考鐘亨任等[20]的方法,通過Live/Dead BacLight Kit試劑盒檢測變異鏈球菌細胞膜的完整性。將變異鏈球菌培養至對數生長期,經PBS稀釋至2×108cfu·mL?1。用500 μL的茴香醛(終濃度為1×MIC,2×MIC或4×MIC)與等體積的稀釋菌液在室溫下共同孵育60 min。等體積的異丙醇和無菌0.9%NaCl處理稀釋菌液分別作為陽性和陰性對照。離心收集變異鏈球菌,并與Live/Dead BacLight Kit試劑盒在避光條件下孵育15 min。孵育后離心去除殘余熒光試劑,通過激光共聚焦掃描顯微鏡(CLSM,Leica,TCS-Sp8)觀察細菌。通過BioFilmAnalyzer v.1.0軟件計算細胞活力。
1.6 茴香醛抑制變異鏈球菌的生物被膜活性測定參考HE等[21]測定茴香醛抑制變異鏈球菌形成生物被膜的活性。將變異鏈球菌培養至對數生長期后,含3%蔗糖的無菌BHI 液體培養基稀釋至2×106cfu·mL?1備用。將變異鏈球菌稀釋液接種到96孔板上,茴香醛的終濃度分別為0.5×MIC,1×MIC和 2×MIC,37℃下培養24 h。以PBS (pH 7.2) 作為空白對照。變異鏈球菌形成生物被膜通過結晶紫測定。最小生物被膜抑制濃度(MBIC50)定義為至少抑制細菌50%生物被膜形成時的最低茴香醛濃度。
1.7 生物被膜清除活性測定將變異鏈球菌培養至對數生長期,含 3% 蔗糖的無菌BHI稀釋至1×106cfu·mL?1備用。將200 μL稀釋菌液接種至96孔板中,在37 ℃下培養24 h后,輕輕將上清液去除,并取200 μL無菌液體培養基稀釋的茴香醛(終濃度0.5×MIC,1×MIC和 2× MIC)加至各實驗孔中。在37 ℃ 下孵育24 h,無菌 PBS (pH7.2) 用作空白對照,采用結晶紫法定量方法測定生物被膜的清除活性。
1.8 細菌表面疏水性測定變異鏈球菌表面疏水性變化采用微生物粘著碳烴化合物法進行測定。在37 ℃有氧條件下用相同濃度茴香醛(0.5×MIC,1MIC×和 2× MIC)分別孵育變異鏈球菌0或30 min后離心,均用無菌PBS洗滌2次,并重懸于相同的緩沖液中。在550 nm處測量吸光度(記錄為OD1),加入 20% (v/v) 二甲苯后劇烈搖動試管,將混合物靜置直至水相與有機相分離,在550 nm處測量水相的吸光度(記錄為OD2)。疏水性百分比由下式計算:H= (OD1?OD2)/OD2×100%。使用0 min (H1)和30 min (H2)值之間的差異確定最終疏水性。
1.9RNA提取和RT-qPCR實驗采用RT-qPCR方法評估茴香醛對變異鏈球菌毒力基因表達的影響。在37 ℃下將變異鏈球菌用不同濃度的茴香醛(0.25×MIC和0.5×MIC)孵育8 h,收集后按照Trizol?試劑(Invitrogen, CA, USA)說明進行總RNA 提取。將純化的 RNA 溶解于20 μL DEPC水中,并在?80 ℃下儲存備用。cDNA合成試劑盒(NOVA, CA,中國)用于合成 cDNA,茴香醛對變異鏈球菌gtf基因 (gtfB,gtfC和gtfD) 表達的影響通過qRT-PCR測定,引物如表1所示。

表1 引物序列
2 結果與分析
2.1 茴香醛對變異鏈球菌的抑菌活性茴香醛的對變異鏈球菌的抑菌活性通過測定MIC值進行定性和定量評估,洗必泰用作陽性對照,以確定茴香醛的抑菌作用。實驗結果表明,茴香醛對變異鏈球菌的MIC為4 g·L?1,MBC為8 g·L?1, 洗必泰對變異鏈球菌的MIC為0.005 g·L?1,MBC為0.01 g·L?1。說明茴香醛對變異鏈球菌具有較好的抑菌活性。
2.2 茴香醛對變異鏈球菌的時間殺菌曲線0.5×MIC、1×MIC和2×MIC的茴香醛對變異鏈球菌的時間殺菌曲線如圖1所示。0.2%洗必泰(CHX)在處理10 min后可完全殺死細菌,空白對照組中顯示細菌數目快速增加,而經不同濃度的茴香醛干預后,作用0~10 min后能有效抑制細菌,在2×MIC濃度下作用30 min后可快速殺死細菌,即使在0.5×MIC的濃度下仍然能夠抑制細菌的增長,呈現出濃度依賴性和時間依賴性的殺菌作用。
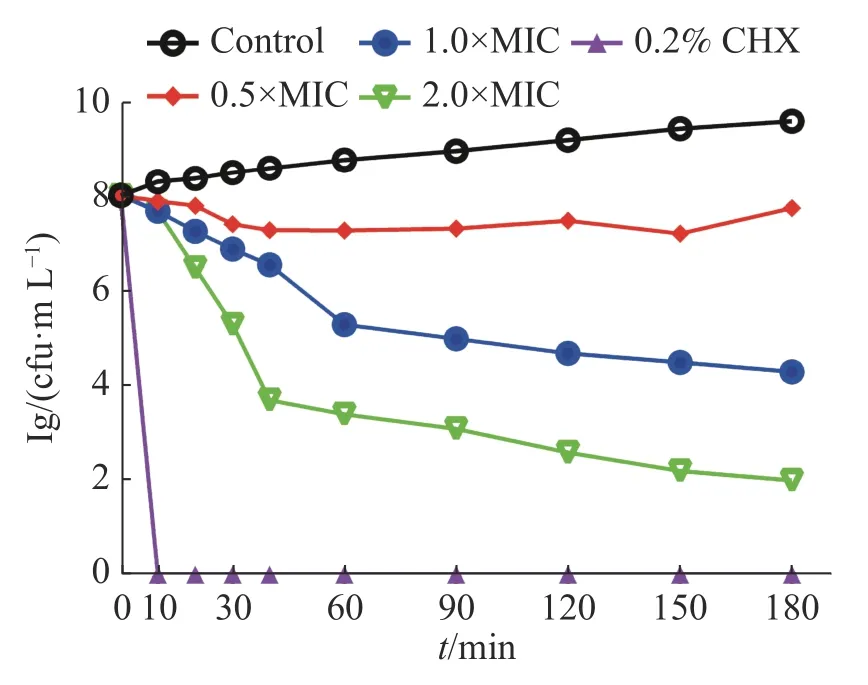
圖1 茴香醛對變異鏈球菌的時間殺菌曲線
2.3 細胞膜完整性檢測
2.3.1 DNA和 RNA泄露檢測通過測定細菌培養液中細菌的核酸泄漏量,研究茴香醛能否靶向作用于變異鏈球菌的細胞膜。通常情況下,大分子物質(包括DNA和RNA)不能透過細菌完整的細胞膜。然而,一旦細菌細菌膜的完整性遭到破壞,大分子物質就會泄漏到細菌外環境中。圖2結果表明,當變異鏈球菌用茴香醛處理時,隨著茴香醛濃度的增加,細菌培養液在260 nm處的吸光值逐漸增加,表明細胞膜損傷程度和細胞膜通透性增強,從而導致細菌的DNA和RNA不斷泄漏。相比之下,空白對照組的吸光度隨著孵育時間的增加而略有變化,這是細菌進行生命活動而產生的代謝產物。結果表明,隨著茴香醛濃度的增加,對變異鏈球菌的細胞膜造成損傷的程度和細胞膜通透性也在加大,導致大分子(DNA和RNA)外流。
2.3.2 細菌蛋白質泄露檢測如圖2-B所示,與空白對照組相比,以不同濃度的茴香醛處理變異鏈球菌后,菌液中可溶性蛋白質含量顯著增加。2×MIC和4×MIC的茴香醛干預細菌1 h后,胞外可溶性蛋白質含量急劇增加,作用于3 h后胞外可溶性蛋白質含量整體趨于平穩狀態。實驗結果與2.3.1中茴香醛對變異鏈球菌胞外核酸泄漏的影響結果相似,進一步表明茴香醛可以改變細菌細胞膜的完整性,隨著茴香醛濃度的增加細菌細胞膜完整性損傷程度也在加大,引起細菌內的DNA、RNA和蛋白質泄漏至細胞外。細菌細胞膜的完整性遭受損傷,干擾了細菌的正常生理活動,進而抑制細菌生長,甚至引起細菌死亡。
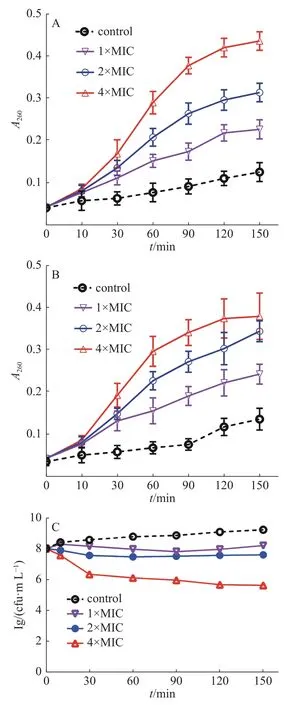
圖2 茴香醛對變異鏈球菌核酸(A)、可溶性蛋白質泄漏(B)和活菌數目(C)的影響
2.3.3 激光共聚焦掃描顯微鏡實驗茴香醛對變異鏈球菌細胞膜完整性的影響通過使用LIVE/DEADBacLight試劑盒檢測(圖3)。與陰性對照(圖3-A)和陽性對照(圖3-B)相比,不同濃度的茴香醛均能誘導變異鏈球菌的細胞膜完整性發生變化。用1×MIC茴香醛處理變異鏈球菌時(圖3-C),細菌被染成橙色,表明茴香醛在該濃度下處理細菌后完整的細胞膜已經開始發生變化。當茴香醛濃度增加至2×MIC和4×MIC時(圖3-D,E),細菌均變成紅色,表明細菌細胞膜的完整性已經嚴重破壞。通過 BioFilmAnalyzer 計算細菌活力(圖3-F),當變異鏈球菌在1×108cfu·mL?1的數量下,2×MIC茴香醛的濃度處理時,大部分的細菌均能被有效抑制。
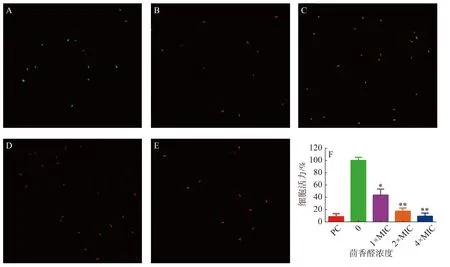
圖3 茴香醛對變異鏈球菌細胞膜完整性的影響
2.3.4 抑制生物被膜活性通過結晶紫法測定茴香醛對變異鏈球菌形成生物被膜的抑制活性和形成生物被膜后的清除活性。如圖4-A中所示,茴香醛在0.5×MIC,1×MIC和2×MIC濃度下均可有效抑制變異鏈球菌的形成和清除成熟生物被膜。如圖4-B所示,茴香醛在0.5×MIC 濃度下對變異鏈球菌形成生物被膜初始附著量為45%~55%。與空白對照相比,變異鏈球菌預先鋪板培養24 h后,茴香醛在1× MIC下對變異鏈球菌生物被膜降解至75%~80%,在 2×MIC 時生物被膜降解至60%~70%。結果表明,茴香醛能有效抑制變異鏈球菌形成生物被膜,也能有效降解成熟生物被膜。
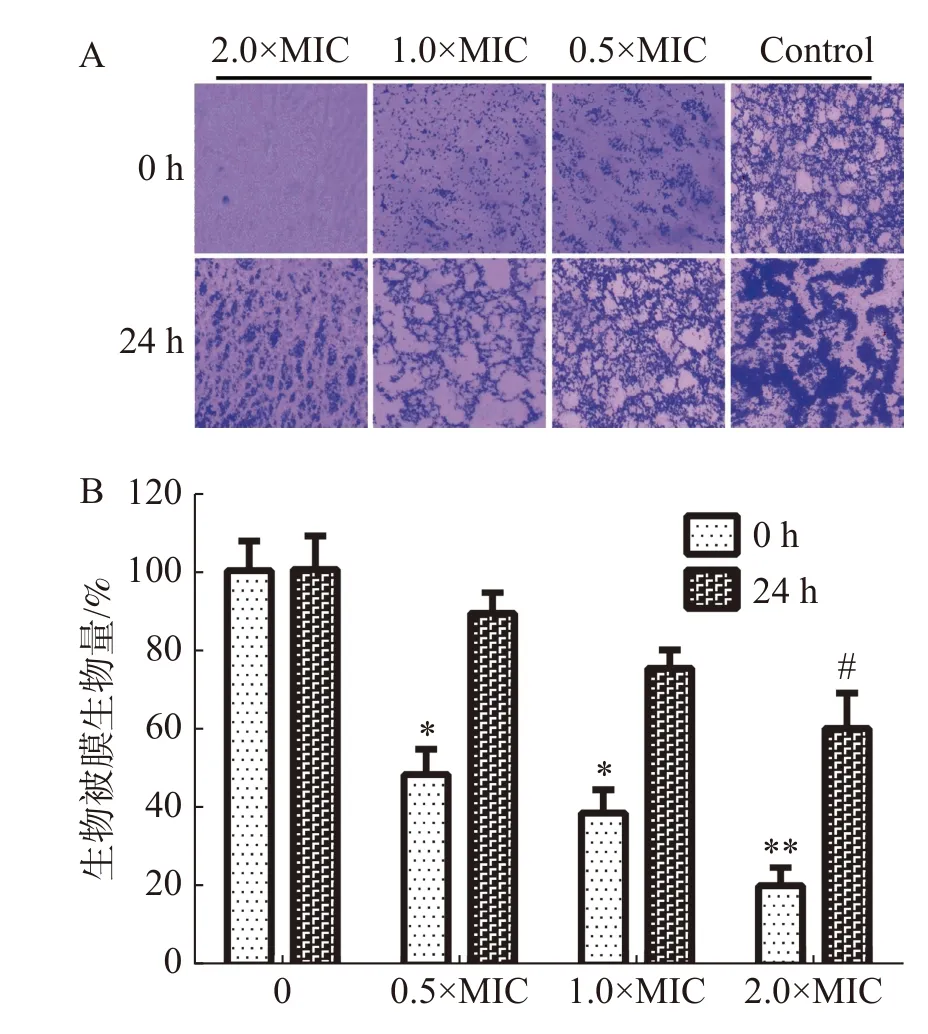
圖4 茴香醛對變異鏈球菌生物被膜的影響
2.3.5 細菌表面疏水性的影響通過檢測變異鏈球菌與碳氫化合物的粘附百分比確定細菌表面的疏水性。圖5結果表明,茴香醛增加了變異鏈球菌的表面疏水性。不同濃度茴香醛(0.5×MIC,1×MIC和2×MIC)處理后的變異鏈球菌表面疏水率分別為(12.35±0.52)%、(14.68±0.44)%和(17.26±0.31)%,顯著高于對照組(6.37±0.13)%。
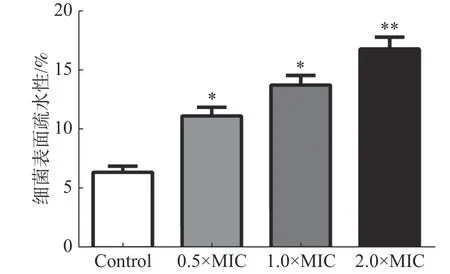
圖5 茴香醛對變異鏈球菌表面疏水性的影響
2.3.6 茴香醛對變異鏈球菌基因表達的影響茴香醛對變異鏈球菌生物被膜形成相關基因表達的影響如圖6所示,結果表明,茴香醛在實驗濃度范圍內影響變異鏈球菌生物被膜形成。gtfB和gtfC基因是合成細菌胞外多糖中的非水溶性胞外多糖的基因,而gtfD是合成水溶性的細胞外多糖的基因。不同濃度的茴香醛處理變異鏈球菌后gtfB、gtfC和gtfD顯著下調(P< 0.05),當0.5×MIC的茴香醛處理變異鏈球菌時,gtfB、gtfC和gtfD表達水平均低于0.8,表明亞濃度的茴香醛能顯著下調gtfs的表達,抑制細菌胞外多糖的產生,干擾細菌生物被膜的形成。
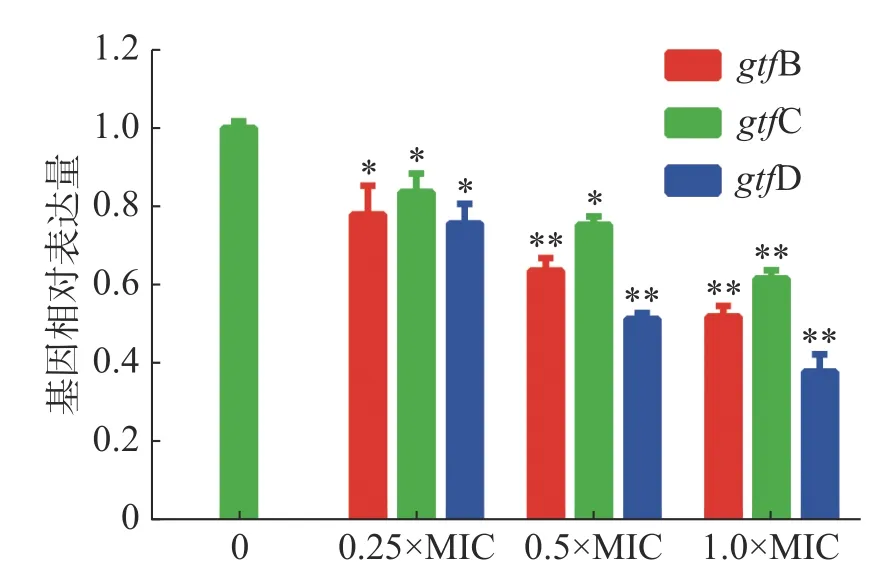
圖6 茴香醛對變異鏈球菌基因表達的影響
3 討 論
口腔疾病一直都是主要的健康問題之一,其中齲齒和牙周病是危害口腔健康的重要疾病。雖然目前醫藥市場上已有甲硝唑、替硝唑和奧硝唑等幾種厭氧菌拮抗劑,但由于口腔環境下活性較低、劑量的使用和各種副作用導致此類藥物的使用范圍受到限制[14]。因此,急需尋找替代性和功能性更強的抗齲齒藥物,天然植物來源藥物由于具有強效抗菌特性和較小的副作用而成為一個重要的研究方向[22]。
茴香醛主要從茴香中分離出來,對許多酵母菌和霉菌菌株也表現出很強的抗真菌活性[23?24]。SHREAZ 等[11]研究結果表明茴香醛對念珠菌的MIC 值為 250 ~ 600 μg·mL?1。YU等[15]發現茴香醛對釀酒酵母和金黃色葡萄球菌的 MIC分別為256 μg·mL?1和 2~4 g·L?1。在本研究中,筆者觀察到茴香醛對變異鏈球菌具有很強的抗菌活性,對變異鏈球菌的MIC 和 MBC 值分別為4 、8 g·L?1。茴香醛在2×MIC 濃度下30 min左右即可殺死所有細菌,即使在0.5×MIC 的濃度下仍然能夠抑制細菌的增長。
細菌細胞膜在保護細菌內大分子物質和維持細胞活力方面起著至關重要的作用,細菌細胞膜完整性缺失將導致細菌死亡[16,21]。許多研究表明,從棕櫚樹、茴香、大蒜、茶樹、百里香和八角中的提取物可以通過破壞細菌細胞膜完整性,抑制微生物的生長[25?28]。1×MIC的茴香醛處理變異鏈球菌60 min后,細菌的細胞膜完整性發生了很大變化,細胞膜內的核酸和蛋白質均發生不同程度的泄漏,導致細菌死亡。
細菌生物被膜的形成可以分為幾個過程,包括最初粘附在固體表面上,可逆附著在固體表面,產生胞外聚合物(EPS),不可逆附著,最終發展為復雜的三維結構。結晶紫法測定表明,隨著茴香醛濃度的增加,茴香醛在初始粘附階段和成熟階段更有效地減弱了變異鏈球菌生物被膜的形成,在高濃度下亦能降解成熟生物被膜。EPS由葡萄糖基轉移酶 (GTF) 合成,在細菌形成生物被膜方面發揮關鍵作用。其中gtfB、gtfC催化蔗糖合成α-1,3和α-1,6鍵連接的非水溶性多糖,這有助于變異鏈球菌產生EPS基質并促進細菌聚集在穩定的生物被膜中,是斑塊形成和細菌生物被膜結構穩定的重要物質。gtfD催化合成含有α-1,6鍵連接的水溶性多糖,促進細菌生物被膜表面與唾液蛋白相結合[28,29]。石榴皮原花青素能有效抑制變異鏈球菌生物被膜的形成和其毒力基因轉錄水平的表達[29]。在生物被膜形成過程中,茴香醛在亞濃度范圍內能有效下調gtfB、gtfC和gtfD的表達。gtfs基因在變異鏈球菌生物被膜中的表達受到抑制,可能會減少生物被膜中葡萄糖基轉移酶的數量,從而減少 EPSs 的產生,抑制生物被膜的形成。變異鏈球菌的產酸能力是變異鏈球菌的關鍵生理因素,作用于牙齒表面的脫礦質促進齲齒形成。積雪草酸和茶黃素對已形成的變異鏈球菌的生物被膜具有顯著的抑制活性以及殺菌作用,并能降低其產酸和產EPS等致齲毒力[30?31]。本研究結果表明,隨著茴香醛濃度的增加,pH下降的初始速率逐漸降低,茴香醛抑制變異鏈球菌產酸并防止牙齒脫礦,這可能是由于抑制了用于產酸的糖酵解酶。
4 結 論
茴香醛對變異鏈球菌表現出較強的抗菌活性,茴香醛抑制變異鏈球菌的機制之一可能與細菌細胞膜通透性的破壞有關,可能通過抑制葡萄糖基轉移酶 (GTF) 合成進而抑制變異鏈球菌形成生物被膜。鑒于目前茴香醛在藥物合成、化妝品、食品添加劑等領域的成熟應用,以及本研究中茴香醛對變異鏈球菌的抑菌活性和抑制生物被膜活性,未來有望進一步開發為抗齲齒藥物或口腔保健品。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
