
釀酒酵母發酵制備蒜氨酸的工藝研究
2022-12-05 12:39:18戴慧敏王立三涂媛蔡俊
中國調味品 2022年12期
關鍵詞:產量
戴慧敏,王立三,涂媛,蔡俊*
(1.湖北工業大學 工業發酵湖北省協同創新中心 工業微生物湖北省重點實驗室 發酵工程教育部重點實驗室,武漢 430068;2.勸農生態農業公司,湖北 恩施 444324)
蒜氨酸,化學名為S-烯丙基-L-半胱氨酸亞砜,是一種存在于百合科蔥屬植物鱗莖中的較穩定的非蛋白質類含硫氨基酸。高純度的蒜氨酸是無臭無味的白色晶體,易溶于水但不溶于乙醇[1-3]。現代科學表明,蒜氨酸有降低膽固醇、預防肥胖、抗腫瘤和抗癌等功能[4-8],與肌苷酸或葡萄糖反應后的分離物具有烤肉味和蒜香味[9],可以被廣泛應用于調味品的生產。此外,蒜氨酸對多種微生物尤其是食品腐敗菌的生長有極強的抑制作用,并且對人體無毒害[10-13],是國內外均認可的食品添加劑[14]。因此,蒜氨酸既可作為良好的食品防腐劑來延長果蔬的保鮮期[15-16],也可以被開發成一種高效的藥物制劑,具有很好的應用前景。
目前蒜氨酸的生產方法主要包括化學合成法[17-19]、組織培養法[20-21]和從植物中提取。化學合成法得率高,但是由于原料中的烯丙基溴具有強毒性,對人體和環境會造成較大的危害,反應收率低,后處理麻煩,并且化學合成會產生兩種立體異構體L-(+)蒜氨酸和L-(-)蒜氨酸,而天然蒜氨酸僅以L-(+)形式存在,因此限制了它的應用范圍。從植物中提取包括水提醇沉法和有機溶劑提取法,水提醇沉法的工藝不夠成熟,而有機溶劑存在潛在的毒性,并且蒜酶的存在會導致蒜氨酸的分解,給蒜氨酸的提取造成了一定的難度,進一步增加了成本[22-24],因此開發綠色安全的生物法生產蒜氨酸勢在必行。
植物體內的蒜氨酸是在半胱氨酸合成酶的作用下,由絲氨酸與乙酰輔酶A結合生成 O-乙酰絲氨酸后,與烯丙基硫醇結合生成 S-烯丙基-L-半胱氨酸,再氧化 S-烯丙基-L-半胱氨酸形成[25]。在微生物體內,絲氨酸乙酰轉移酶(SAT)和O-乙酰絲氨酸(硫醇)裂解酶(OASS)共同組成酶復合體,即半胱氨酸合成酶。微生物也能利用半胱氨酸合成酶將環境中的無機硫還原成S2-,進入到半胱氨酸的合成中。而OASS能利用半胱氨酸生物合成體系,以親核試劑和硫醇代替各種S2-合成非蛋白質氨基酸[26-27]。目前已有研究報道了利用大腸桿菌中重組表達的OASS合成藥物中間體S-苯基-L-半胱氨酸及L-α-丙氨酸[28],為利用OASS在微生物體內合成蒜氨酸提供了參考。
本研究以烯丙基硫醇為底物,從實驗室保藏的19種酵母菌株中,篩選出一株S-烯丙基-L-半胱氨酸產量較高的釀酒酵母CCCG,研究了溫度、初始pH、接種量、裝液量、搖瓶轉速、烯丙基硫醇添加量等發酵條件對釀酒酵母產S-烯丙基-L-半胱氨酸含量的影響,通過單因素試驗和正交試驗,經釀酒酵母的發酵及H2O2的氧化得到蒜氨酸的最佳發酵條件,為生物法合成蒜氨酸提供了新的思路。
1 材料與方法
1.1 材料與試劑
1.1.1 菌種
實驗室保藏釀酒酵母菌株:CGY2、CGY、CCCG、CLS、CCY、CHY、CBY、CGWY、CEPY、CKY、CMY、CRY、BDYD、JYYJS、JYYR、JYFI、GAQ1、GAQ2、
GAQ4。
1.1.2 試劑
葡萄糖、蛋白胨、酵母浸粉、硫酸銨、磷酸二氫鉀、硫酸鎂:國藥集團化學試劑有限公司;甲醇(色譜純):美國Fisher Scientific公司;蒜氨酸標準品(純度≥98%):上海源葉生物科技有限公司;S-烯丙基-L-半胱氨酸標準品(純度≥98%)、烯丙基硫醇(分析純):上海阿拉丁生化科技股份有限公司。
1.1.3 培養基
斜面培養基:葡萄糖20 g/L,蛋白胨20 g/L,酵母粉10 g/L,瓊脂20 g/L,pH 6.0,在115 ℃下滅菌30 min。
種子培養基:葡萄糖20 g/L,蛋白胨20 g/L,酵母粉10 g/L,pH 6.0,在115 ℃下滅菌30 min。
發酵培養基:葡萄糖20 g/L,酵母粉10 g/L,(NH4)2SO48 g/L,KH2PO43 g/L,MgSO40.15 g/L,pH 6.0,在115 ℃下滅菌30 min。
1.2 儀器與設備
LD-20AD高效液相色譜系統(配有SPD-20A紫外檢測器) 日本島津公司;質譜引導的高通量自動純化系統(配有電噴霧離子源(ESI)) 沃特世科技(上海)有限公司;WH-2微型漩渦混合儀 上海滬西分析儀器廠有限公司;JY92-Ⅱ超聲波細胞粉碎機 寧波新芝生物科技股份有限公司。
1.3 方法
1.3.1 菌種篩選
分別將活化的菌種接種到發酵培養基中,接種量為10%,轉速為180 r/min,在30 ℃培養12 h后,加入烯丙基硫醇,繼續培養19 h。由于S-烯丙基-L-半胱氨酸經過H2O2的氧化后即為蒜氨酸,因此先篩選出能產生S-烯丙基-L-半胱氨酸的菌株。培養結束取樣處理后,利用HPLC測定其中S-烯丙基-L-半胱氨酸的含量,而后在經過處理的樣品中加入30% H2O2,持續攪拌24 h后測定其中蒜氨酸的含量[29-30],篩選出S-烯丙基-L-半胱氨酸和蒜氨酸產量較高的菌株,并利用質譜法進一步驗證所篩選的菌株具有產S-烯丙基-L-半胱氨酸的能力。
1.3.2 樣品處理
水分含量測定:菌懸液經8000 r/min離心去上清液,無菌水清洗菌體3次,稱取濕菌體重量M1,置于65 ℃烘箱中烘干至恒重,稱取干菌體重量M2。

(1)
式中:M1為濕菌體的重量,g/L;M2為干菌體的重量,g/L。
破壁提取胞內S-烯丙基-L-半胱氨酸:菌懸液經8000 r/min離心去上清液,無菌水洗滌3次,稱取菌體濕重,重懸濕菌體定容至恒定體積。定容后取適量菌懸液稀釋,于血球計數板計數,取3次平行計數結果。采用超聲破壁法,設定功率為300 W,于冰浴條件下超聲破碎30 min(設定工作1 s,間隔1 s)[31- 32],取適量破壁后的菌懸液稀釋,于血球計數板計數,取3次平行計數結果。
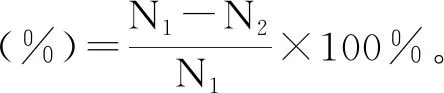
(2)
式中:N1為破壁前細胞數;N2為破壁后細胞數。
胞內S-烯丙基-L-半胱氨酸含量測定:破壁后的菌懸液經8000 r/min離心取上清液,將上清液用微濾膜(0.22 μm)過膜后利用高效液相色譜法計算得到提取液中S-烯丙基-L-半胱氨酸的濃度,最后計算得到每克干細胞中S-烯丙基-L-半胱氨酸的含量。
S-烯丙基-L-半胱氨酸干重產量(mg/g)=
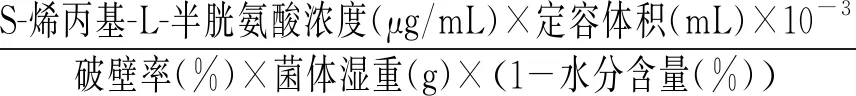
(3)
1.3.3 S-烯丙基-L-半胱氨酸和蒜氨酸的定量和定性分析
1.3.3.1 S-烯丙基-L-半胱氨酸和蒜氨酸的定量分析
色譜條件:色譜柱Inertsil ODS SP C18(4.6 mm×150 mm, 5 μm);流動相甲醇∶甲酸水溶液為5∶95(體積比);紫外檢測波長220 nm;柱溫30 ℃,流速0.8 mL/min,進樣量20 μL。
1.3.3.2 標準曲線繪制
分別配制濃度為200,400,600,800,1000 μg/mL的S-烯丙基-L-半胱氨酸標準溶液和蒜氨酸標準溶液。以不同的S-烯丙基-L-半胱氨酸和蒜氨酸標準溶液濃度為橫坐標,以HPLC法測定的峰面積數值為縱坐標,繪圖得到S-烯丙基-L-半胱氨酸和蒜氨酸標準曲線。
1.3.3.3 S-烯丙基-L-半胱氨酸和蒜氨酸的定性分析
質譜條件:離子方式:電噴霧正離子化(ESI+);毛細管電壓3.5 kV;錐孔電壓25 V;離子源溫度100 ℃;脫溶劑氣溫度300 ℃;質量掃描范圍100~200 m/z;選擇性離子檢測(SIM)162 m/z和 178 m/z。
1.3.4 單因素試驗設計優化發酵條件
通過單因素試驗改變發酵條件,分別考察烯丙基硫醇添加量、初始pH、溫度、搖瓶轉速、接種量、裝液量對S-烯丙基-L-半胱氨酸產量的影響,單因素試驗設計表見表1,按照每個因素分別進行試驗研究。

表1 單因素不同水平試驗設計表Table 1 Single factor test design at different levels
1.3.5 正交試驗設計優化發酵條件
在單因素試驗的基礎上,采用七因素三水平,通過正交試驗L18(37)選取最優發酵條件,因素水平設計表見表2。
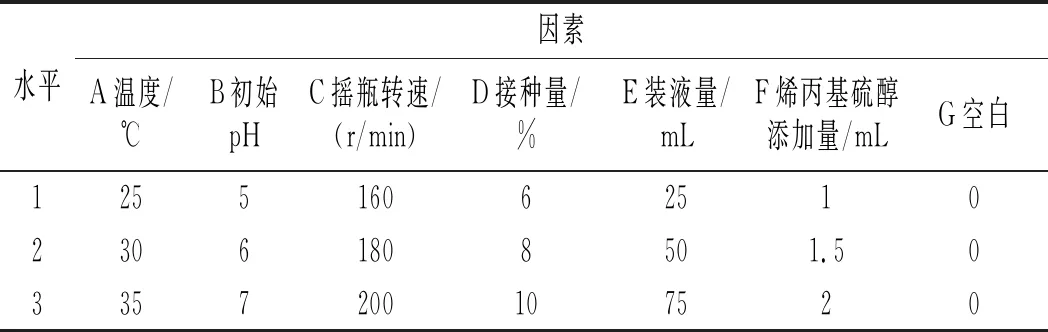
表2 正交試驗因素水平表Table 2 The factors and levels of orthogonal test
2 結果與分析
2.1 高效液相色譜法測定S-烯丙基-L-半胱氨酸和蒜氨酸的標準曲線回歸方程
按照1.3.3.1中的方法,在200 μg/mL~1 mg/mL范圍內,進樣量與其峰面積均呈良好的線性關系,S-烯丙基-L-半胱氨酸的回歸方程為:Y=5496.3X-44949(R2=0.9997),蒜氨酸的回歸方程為:Y=7232.5X+39674(R2=0.9998)。
2.2 高產S-烯丙基-L-半胱氨酸的菌株的篩選
通過高效液相色譜法測定15種酵母細胞合成S-烯丙基-L-半胱氨酸的產量,結果表明有10種酵母菌株具有利用烯丙基硫醇合成S-烯丙基-L-半胱氨酸的能力,在產S-烯丙基-L-半胱氨酸的菌株中,有兩株酵母干重產量超過50 mg/g DCW,分別為酵母CCCG和酵母BDYD,干重產量分別為(52.35±0.86) mg/g DCW和(50.16±1.00) mg/g DCW;通過對兩種菌株重復驗證試驗發現,酵母CCCG合成S-烯丙基-L-半胱氨酸的干重產量明顯高于酵母BDYD,因此選擇釀酒酵母CCCG作為發酵制備蒜氨酸的菌株。

圖1 不同酵母菌株S-烯丙基-L-半胱氨酸產量Fig.1 S-allyl-L-cysteine yield of different Saccharomyces cerevisiae strains
2.3 S-烯丙基-L-半胱氨酸及蒜氨酸的定性分析
對S-烯丙基-L-半胱氨酸標品、酵母CCCG的細胞破碎液、蒜氨酸標品及酵母CCCG的細胞破碎液經雙氧水處理后的樣品分別進行ESI-MS分析,進一步驗證S-烯丙基-L-半胱氨酸和蒜氨酸的生成,試驗結果見圖2和圖3。

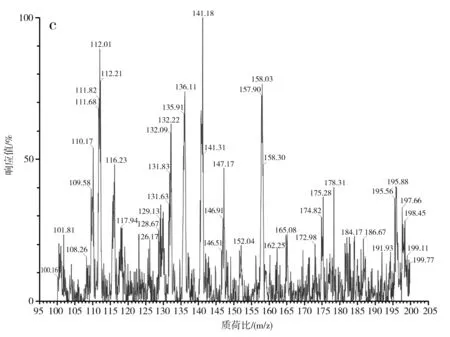
圖2 S-烯丙基-L-半胱氨酸電噴霧電離源-質譜圖Fig.2 ESI-MS of S-allyl-L-cysteine
由圖2中a和b可知,S-烯丙基-L-半胱氨酸標品和酵母CCCG的細胞破碎液的準分子離子流圖均在3.42 min左右出峰。由圖2中c可知,酵母CCCG的細胞破碎液在3.42 min處出現m/z=162.25,為S-烯丙基-L-半胱氨酸的準分子離子[M+H]+,因此確定酵母CCCG具有合成S-烯丙基-L-半胱氨酸的能力。
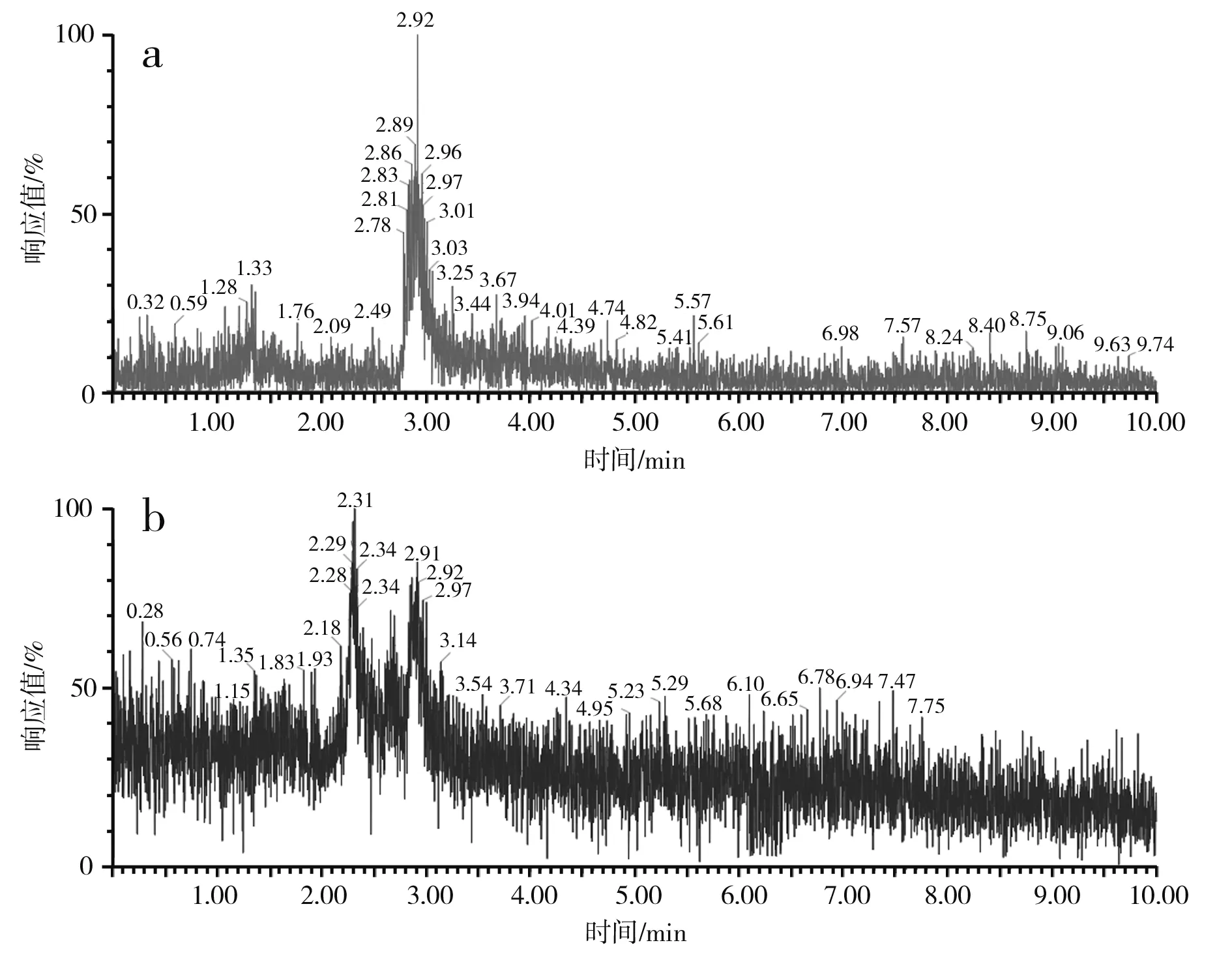
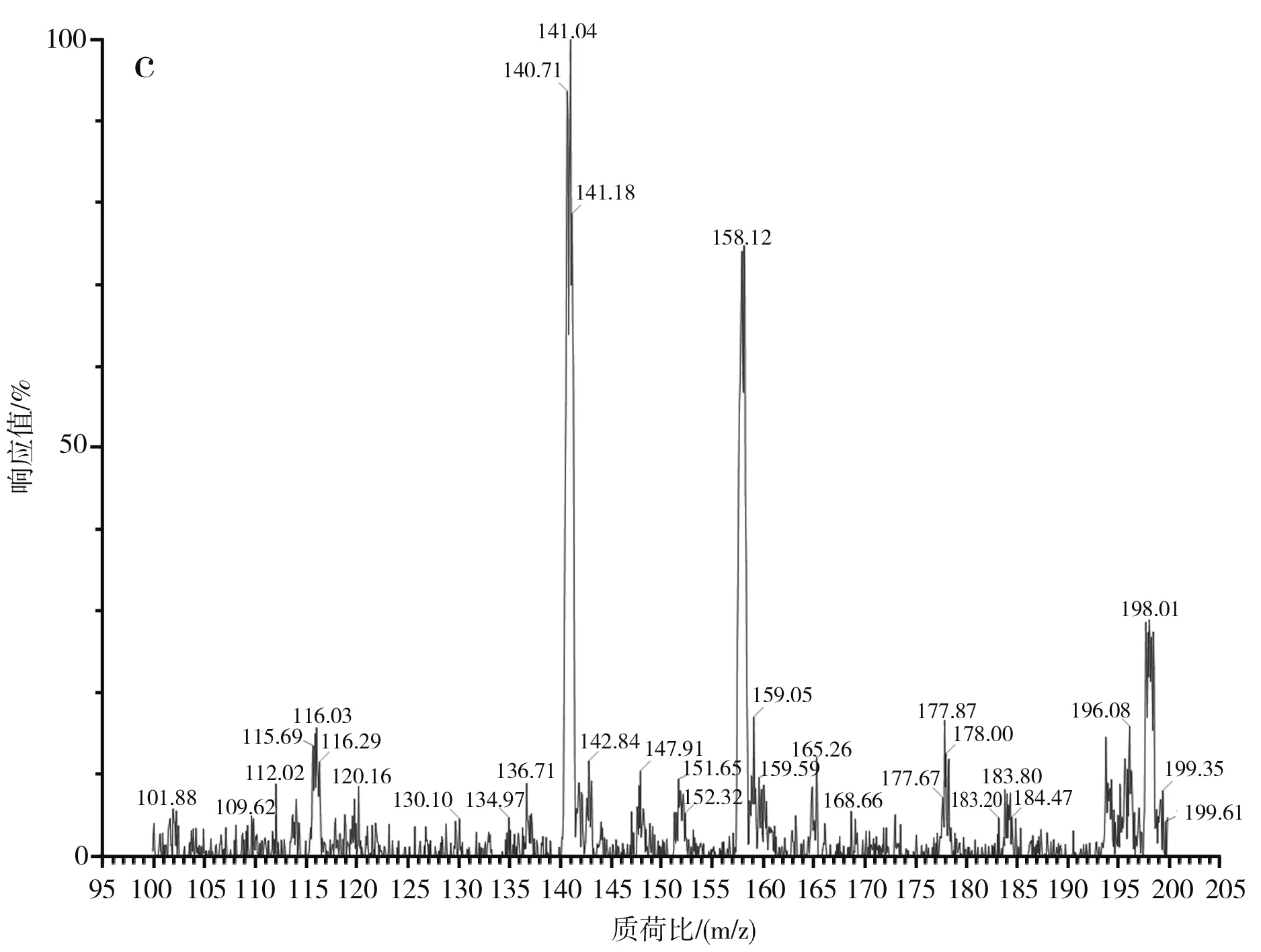
圖3 蒜氨酸電噴霧電離源-質譜圖Fig.3 ESI-MS of alliin
由圖3中a和b可知,蒜氨酸標品和酵母CCCG的細胞破碎液經H2O2處理后的準分子離子流圖均在2.88 min左右出峰。由圖3中c可知,酵母CCCG的細胞破碎液經H2O2處理后的樣品在2.887 min處出現m/z=178.00,為蒜氨酸的準分子離子[M+H]+,確定了產物蒜氨酸的生成。
2.4 發酵條件對S-烯丙基-L-半胱氨酸產量的影響
不同發酵條件對S-烯丙基-L-半胱氨酸產量的影響見圖4 。
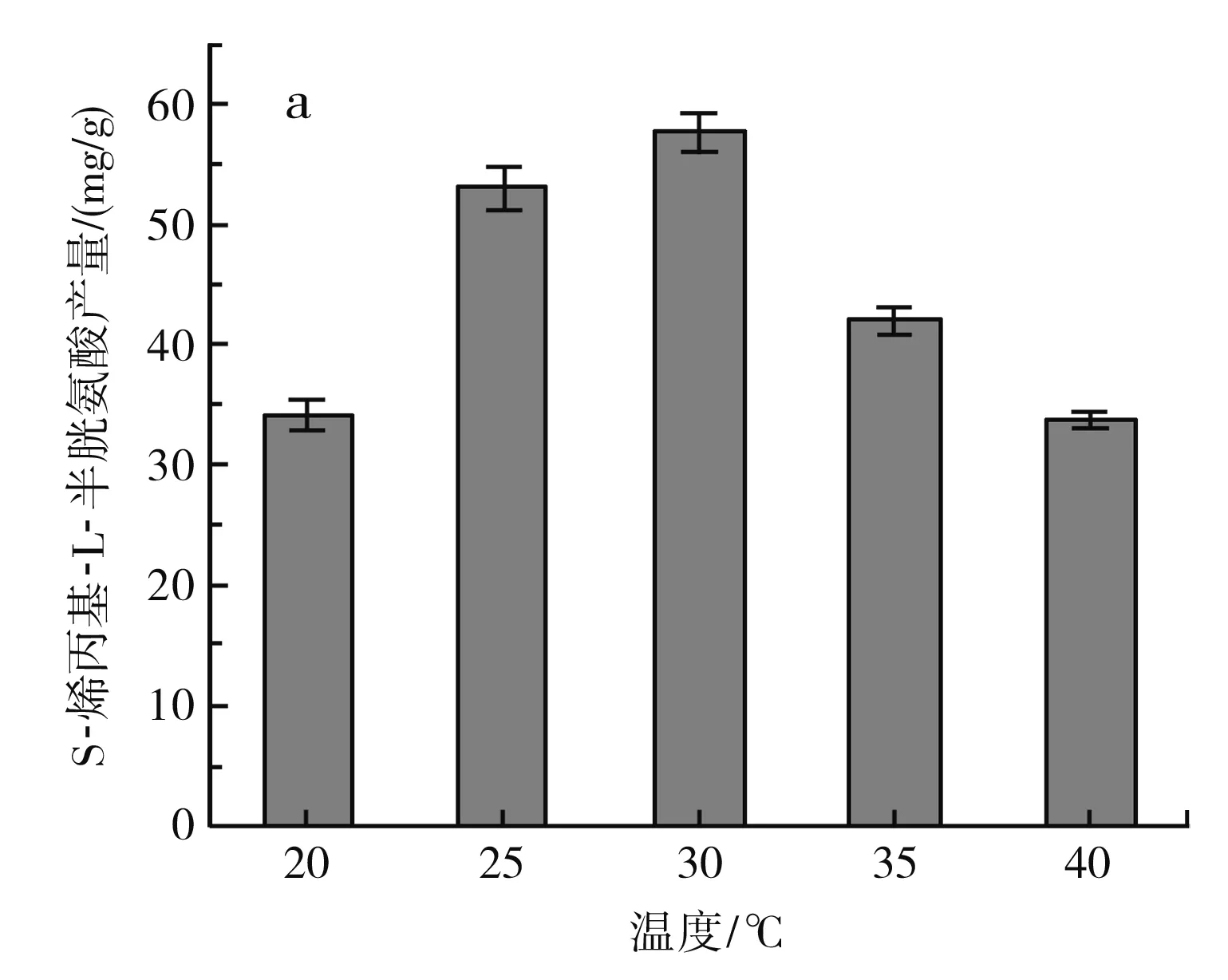
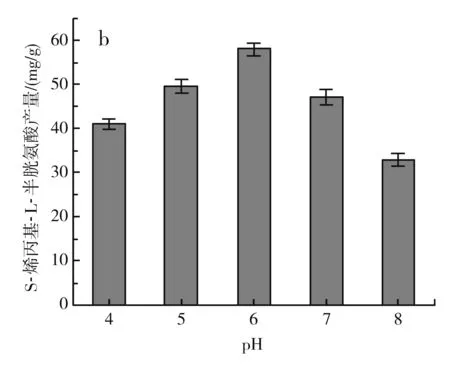



圖4 不同發酵條件對S-烯丙基-L-半胱氨酸干重產量的影響Fig.4 Effect of different fermentation conditions on dry weight yield of S-allyl-L-cysteine
由圖4中a可知,溫度在20~30 ℃時,S-烯丙基-L-半胱氨酸的產量隨著溫度的升高而增加,在30 ℃時達到最大,為(57.7±1.65) mg/g DCW,但發酵溫度繼續升高,S-烯丙基-L-半胱氨酸的產量下降,可能是由于溫度過高,發酵過程中添加的一部分烯丙基硫醇未反應就已經分解,從而降低了S-烯丙基-L-半胱氨酸的產量。
由圖4中b可知,當pH在4~6之間時,S-烯丙基-L-半胱氨酸的產量逐漸增加,當pH為6時,S-烯丙基-L-半胱氨酸的產量達到最大,約為(58±1.44) mg/g DCW,pH繼續升高,S-烯丙基-L-半胱氨酸的產量下降,可能是由于pH過高影響了酵母的生長,從而影響了S-烯丙基-L-半胱氨酸的產量。
由圖4中c可知,當接種量在6%~10%之間時,S-烯丙基-L-半胱氨酸的產量隨著接種量的增加而增加,當接種量為10%時,S-烯丙基-L-半胱氨酸的產量達到最大,為(56.2±0.89) mg/g DCW,當接種量繼續增大時,S-烯丙基-L-半胱氨酸的產量減少,可能是因為接種量較少時,菌體的對數生長期會延長,導致處于生長繁殖期的菌體的發酵能力較差,S-烯丙基-L-半胱氨酸的合成不夠,接種量較大時,發酵時間縮短,降低了S-烯丙基-L-半胱氨酸的產量,因此選取接種量10%為最佳。
由圖4中d可知,搖瓶轉速在140~180 r/min時,S-烯丙基-L-半胱氨酸的產量逐漸增加,當搖瓶轉速繼續增加時,S-烯丙基-L-半胱氨酸的產量下降,故確定最佳搖瓶轉速為180 r/min。
由圖4中e可知,裝液量在25~50 mL之間時,S-烯丙基-L-半胱氨酸的產量逐漸增加,當裝液量在50 mL時,S-烯丙基-L-半胱氨酸的產量最大,為(60.2±0.9) mg/g DCW,裝液量繼續增加,S-烯丙基-L-半胱氨酸的產量下降,因此確定最佳裝液量為50 mL。
2.5 烯丙基硫醇添加量對S-烯丙基-L-半胱氨酸產量的影響
由圖5可知,當烯丙基硫醇添加量在0.5~1.5 mL之間時,S-烯丙基-L-半胱氨酸的產量隨著烯丙基硫醇添加量的增加而增加,當烯丙基硫醇添加量為1.5 mL時,S-烯丙基-L-半胱氨酸的產量達到最大,為(53.6±1.43) mg/g DCW,可能是由于烯丙基硫醇添加量過大時,會影響菌體的生長,從而導致S-烯丙基-L-半胱氨酸的產量下降,因此確定最佳的烯丙基硫醇添加量為1.5 mL。

圖5 烯丙基硫醇添加量對S-烯丙基-L-半胱氨酸干重產量的影響Fig.5 Effects of allyl mercaptan addition amount on dry weight yield of S-allyl-L-cysteine
2.6 正交試驗結果及分析
正交試驗結果分析見表3。
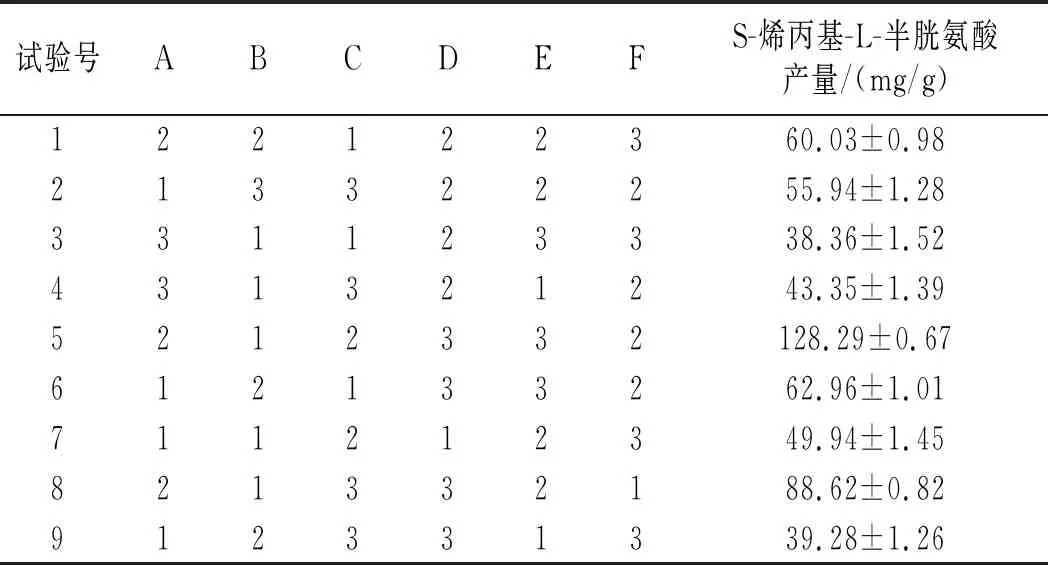
表3 L18(37)正交試驗設計及結果Table 3 L18(37) orthogonal test design and results
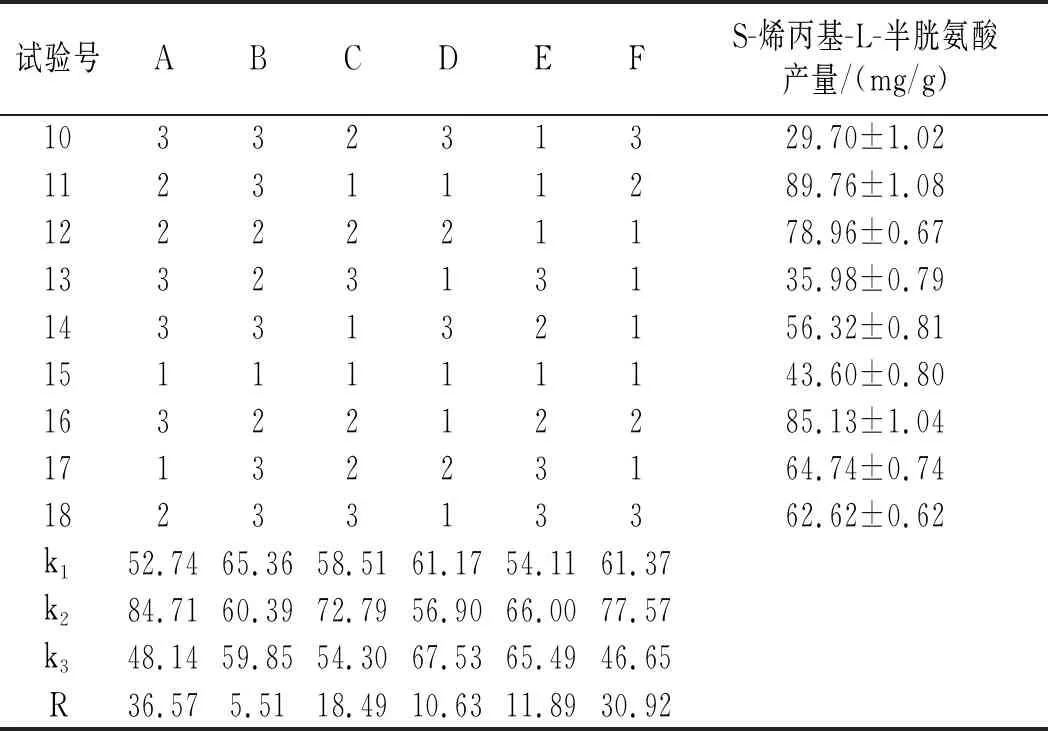
續 表
由表3可知,6個因素的影響程度由高到低為溫度>烯丙基硫醇添加量>搖瓶轉速>裝液量>接種量>初始pH,最優水平組合為A2F2C2E2D3B1,正交試驗方差分析結果見表4。
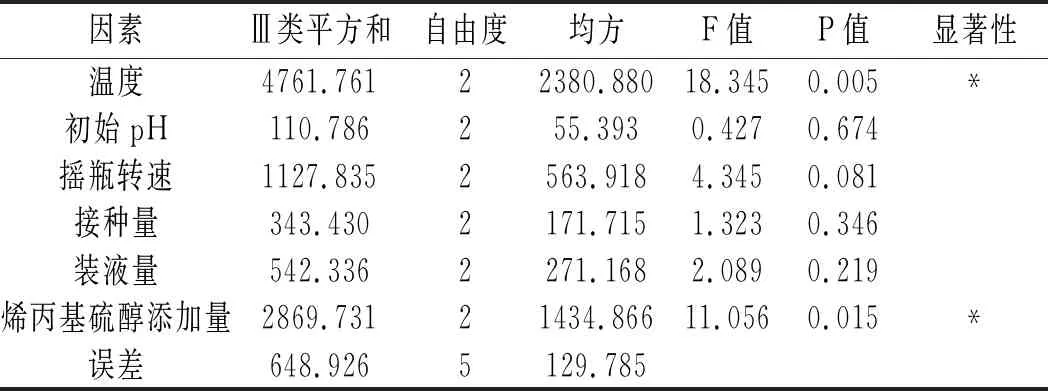
表4 正交試驗方差分析結果Table 4 The variance analysis results of orthogonal test
由表4可知,溫度和烯丙基硫醇添加量均為顯著因素,其中溫度的顯著性大于烯丙基硫醇添加量的顯著性,正交試驗R2=0.938,說明試驗中有93.8%的數據可信。
2.7 正交驗證試驗
對S-烯丙基-L-半胱氨酸標品、經試驗驗證最優組合得到的酵母細胞CCCG的細胞破碎液、蒜氨酸標品及細胞破碎液經雙氧水處理后的樣品分別進行HPLC分析,試驗結果見圖6和圖7。
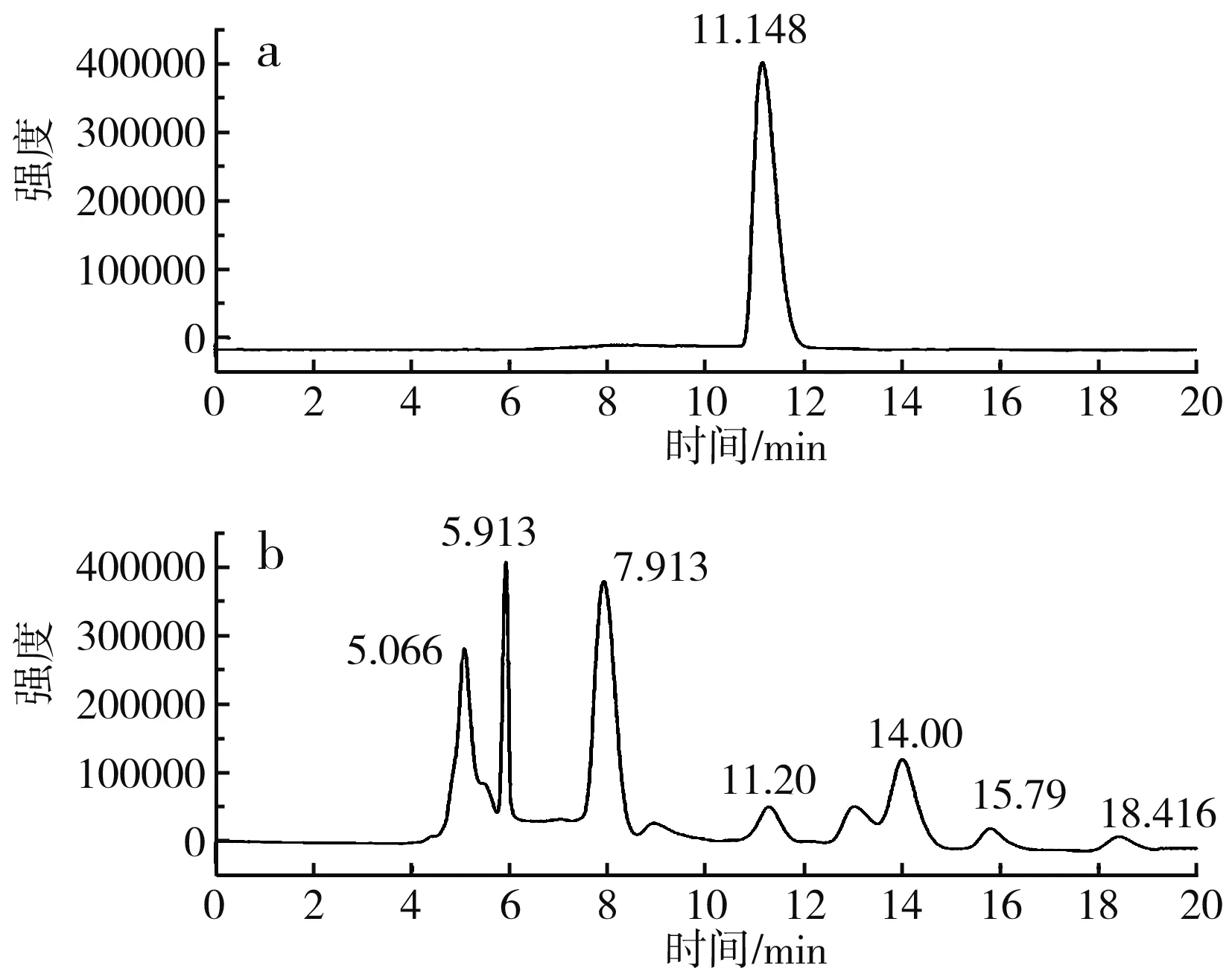
圖6 S-烯丙基-L-半胱氨酸高效液相色譜圖Fig.6 HPLC of S-allyl-L-cysteine
由圖6可知,S-烯丙基-L-半胱氨酸標品的出峰時間為11.148 min,細胞破碎液在11.20 min出峰,與標品出峰時間相近,證明有產物S-烯丙基-L-半胱氨酸的生成。
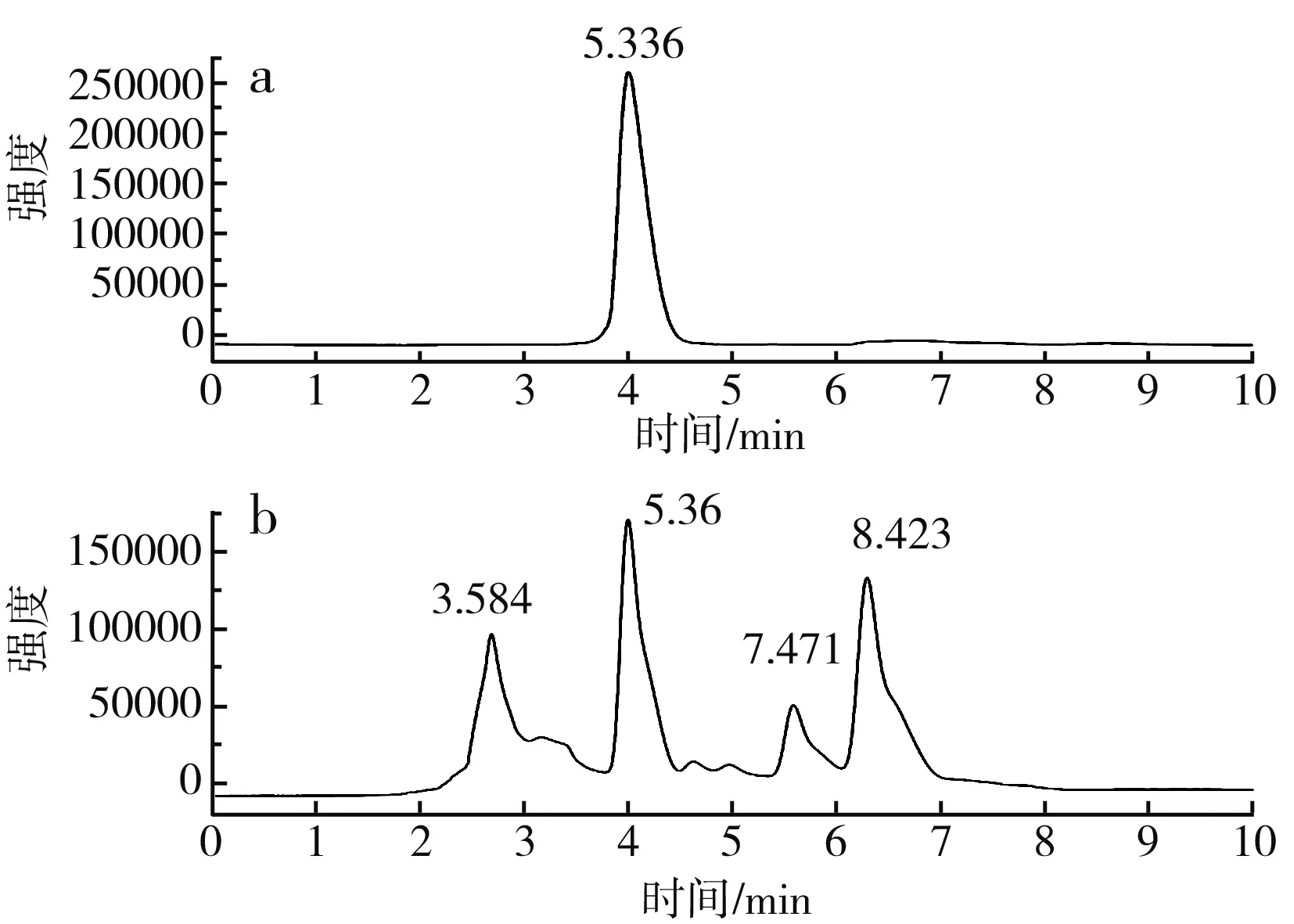
圖7 蒜氨酸高效液相色譜圖Fig.7 HPLC of alliin
由圖7可知,蒜氨酸標品的出峰時間為5.336 min,細胞破碎液在5.36 min出峰,與標品出峰時間相近,證明有產物蒜氨酸的生成。
經方差分析得到的最優組合為A2B1C2D3E2F2,經試驗驗證最優組合的S-烯丙基-L-半胱氨酸干重產量為(136.01±0.88) mg/g DCW,正交試驗中最佳組合A2B1C2D3E3F2的試驗結果為(128.29±0.67) mg/g DCW,經H2O2氧化后得蒜氨酸的干重產量為(68.281±0.93) mg/g DCW。綜合k值和正交試驗方差分析結果可知,利用釀酒酵母產蒜氨酸的最優發酵條件為A2B1C2D3E2F2,即培養溫度為30 ℃,初始pH為5,搖瓶轉速為180 r/min,接種量為10%,裝液量為50 mL/250 mL,烯丙基硫醇添加量為1.5 mL。
3 結論
蒜氨酸作為一種天然活性物質,性質穩定、生物活性高且安全無毒,其受熱分解能產生多種具有特殊風味和生物活性的含硫化合物[33]。酵母作為國際上公認的安全的食品級微生物,其抽提物也是一種純天然和營養的食品輔料[34-35]。因此,利用釀酒酵母發酵制備蒜氨酸在天然植物防腐保鮮劑、食品調味劑、保健品等方面具有極大的應用潛力。
本研究以烯丙基硫醇為底物,從19種酵母中篩選出能產生S-烯丙基-L-半胱氨酸的菌株,并通過H2O2的氧化作用,產生蒜氨酸,通過ESI-MS和HPLC法驗證了產物的生成。通過單因素和正交試驗對產S-烯丙基-L-半胱氨酸的發酵條件進行了優化,得最優發酵條件為發酵時間19 h,發酵溫度30 ℃,接種量10%,初始pH 5,搖瓶轉速180 r/min,裝液量50 mL/250 mL,烯丙基硫醇添加量1.5 mL,在此條件下,經H2O2的氧化作用,蒜氨酸的干重產量為(68.281±0.93) mg/g DCW,比優化前提高了1.96倍。本研究利用蒜氨酸在植物體內的生物合成途徑,以烯丙基硫醇為底物,作用于釀酒酵母CCCG的相關代謝途徑中,實現了蒜氨酸的生物合成,為生物法合成蒜氨酸的研究提供了數據參考。后續研究將從代謝途徑的調控入手,通過補料分批發酵提高底物的利用率以及由底物烯丙基硫醇直接發酵合成蒜氨酸;尋找替代外加H2O2氧化的方法,以期提高S-烯丙基-L-半胱氨酸轉化為蒜氨酸的轉化率,進一步優化生物法合成蒜氨酸的工藝。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
