黃顙魚XX偽雄魚誘導與全雌種群規模化繁育
2022-12-21 07:24:56魯子怡樊啟學沈志剛
水生生物學報 2022年12期
劉 婭 于 躍 魯子怡 丁 洋 樊啟學,2 沈志剛
(1.華中農業大學水產學院, 農業農村部淡水生物繁育重點實驗室, 教育部長江經濟帶大宗水生生物產業綠色發展工程研究中心, 池塘健康養殖湖北省工程實驗室, 武漢 430070; 2.湖北黃優源漁業發展有限公司, 武漢 430299)
水產養殖是“藍色食品”的主要來源[1], 是人類蛋白質可持續發展的重要組成部分, 也是全世界增長速度最快的食品生產領域[2]。眾多水產養殖動物在生長速度與最終規格、繁殖行為與性腺價值、肉質與口味、形態與觀賞價值等方面表現出明顯的性別二態性(Sexual dimorphism)[3,4]。這些表現是水產動物非常重要的養殖性能, 因此, 水產養殖對性別控制具有重大需求。性別控制育種是水產動物遺傳育種中最重要的育種方式之一, 單性種群養殖能提高魚體規格整齊度, 減少不必要的能量消耗, 降低養殖成本, 滿足人民對于特定性別性腺的需求, 解決親本短缺等問題[5]。全世界淡水養殖品種產值前80位的魚類中, 超過三分之二的種類對單性種群有需求, 包括鮭鱒類、四大家魚、羅非魚(Oreochromis niloticus)、鱖(Siniperca chuatsi)、鯰類、加州鱸(Micropterus salmoides)等[6,7]。在這其中, 部分種類單性種群已經實現規模化生產, 包括全雌虹鱒、全雄羅非魚、全雌鱖、全雄黃顙魚等[8],推動了養殖產業的發展。
黃顙魚(Tachysurus fulvidraco)屬于鯰形目、鲿科、黃顙魚屬, 在我國分布廣泛, 除西部高原及新疆外, 廣布于各個水系, 野外資源豐富。其無肌間刺、肉質鮮嫩、經濟價值高, 養殖地區已覆蓋全國27個省份, 2020年產量達到56.5×107kg, 全國淡水魚類養殖產量排名第10位(2021中國漁業統計年鑒)。在過去20余年的規模化養殖過程中, 2010年認定的 “全雄1號” (黃顙魚T.fulvidraco♀×YY超雄黃顙魚T.fulvidraco♂)和2019年認定的雜交黃顙魚“黃優1號” (黃顙魚T.fulvidraco♀×瓦氏黃顙魚T.vachelli♂)兩個國家審定新品種極大推動了黃顙魚養殖產業的穩步增長, 是過去10年時間內產量增長最快(15.0%)的兩個淡水養殖種類之一(另一個是加州鱸)。但是目前, 一方面黃顙魚雌性親本養殖量極度縮減, 另一方面野生種群不可再作為繁殖親本,第三是在黃顙魚人工繁殖過程中, 雌性親本死亡率一般高達30%。這三方面的因素, 導致我國黃顙魚雌性親本嚴重不足。因此, 黃顙魚母本的短缺, 已成為制約黃顙魚養殖產業發展的最重要因素之一。
當前黃顙魚母本的獲得, 是通過人工繁殖獲得XX雌魚和XY雄魚混合群體, 在養殖達到50 g左右規格時, 將XY雄魚通過生殖突挑出, 留下XX雌魚繼續培育至性成熟。這種生產方式效率低下, 浪費大量池塘、人力和飼料資源, 也不利于新品種選育工作的開展[9,10], 亟須建立黃顙魚全雌種群的生產體系。黃顙魚性別決定方式屬于遺傳-環境共同決定類型, 或者更準確地說, 是遺傳型性別決定[11,12]同時受環境應激影響[6,13]。因此, 進行全雌黃顙魚規模化生產, 首先是要誘導XX遺傳型個體性逆轉,批量獲得功能性的XX偽雄魚。早在2013年,Shen等[14,15]發現, 來曲唑(Letrozole, LZ)處理可以增加黃顙魚群體的雄性比例, 推測有XX遺傳型個體成功性逆轉為XX雄魚; 而17α-甲基睪丸酮(17αmethyltestosterone, MT)處理沒有顯著改變雄性比例, 而是誘導了大量間性個體的產生。但是在這個階段, 黃顙魚性別特異性分子標記還未見報道, 并不能完全確定性逆轉的比例及鑒定出性逆轉的個體。此后, 借助黃顙魚性別特異性分子標記[12], 楊天毅等[16]進一步確認了MT處理導致精巢發育異常,但LZ能誘導XX個體雄性化, 并且部分XX偽雄魚具備繁殖能力。本研究團隊前期發現, 黃顙魚性別分化關鍵時期用33.5℃高溫處理, 能誘導39.1%的XX遺傳型個體雄性化[6]。這些前期工作, 為黃顙魚性別控制奠定了重要基礎, 但全雌黃顙魚種群的規模化繁育及其性腺發育情況還未見報道。
為了滿足黃顙魚養殖產業對母本的大量需求,本研究使用高溫、LZ和MT共同作用誘導黃顙魚性逆轉, 獲得了具有繁殖能力的XX偽雄魚, 將其與正常XX母本進行規模化人工繁殖, 獲得了57萬尾仔魚; 利用性別特異性分子標記, 鑒定該群體全部為XX遺傳型; 組織學結果表明, XX全雌魚群體91.9%個體卵巢發育正常, 8.1%個體可能受到池塘高水溫影響而性逆轉為XX雄魚。研究結果將為黃顙魚產業的健康可持續發展奠定重要基礎。
1 材料與方法
1.1 XX偽雄魚制備
普通黃顙魚雌魚與雄魚來自湖北黃優源漁業發展有限公司選育群體。選取規格一致, 無病無傷且體質健壯的親本進行人工繁殖, 受精卵出膜后帶回華中農業大學水產學院科研基地進行養殖。從3日齡開始用豐年蟲進行投喂, 隨后逐步馴食微粒飼料(山東升索漁用飼料研究中心, 主要成分: 粗蛋白質≥50.0%, 粗脂肪>8.0%, 粗纖維≤3.0%, 灰分≤16.5%, 水分≤12.0%, 鈣≤5.0%, 總磷≥1.0%,賴氨酸≥2.0%)。
為了誘導雌魚雄性化, 基于實驗室前期研究[6,14,15],我們采取LZ、MT和高溫共同處理黃顙魚幼魚。處理條件為300 mg/kg LZ、5 mg/kg MT和33.5℃高溫, 處理時間為12至65日齡, 覆蓋了整個黃顙魚性別分化關鍵時期。高溫處理通過加熱棒實現, 白天每隔6h測定一次水溫, 以保證高溫的維持和穩定。激素飼料的制備是通過將LZ與MT溶于95%乙醇溶液, 再將配制好的酒精均勻地噴灑在微粒飼料表面,放入通風櫥中使酒精自然晾干, 從而將激素有效配制至飼料中; 對照組飼料同樣用乙醇溶液處理, 所有制備好的飼料均置于4℃條件下備用。對照組養殖水溫為(26±2)℃。飼料與乙醇的比例參照實驗室之前的配制比例[14,15], 略有調整, 每100 g飼料使用10—30 mL乙醇溶液。實驗分處理組與對照組兩個組, 每組3個重復, 每個重復500尾魚。
實驗期間魚苗飼養在360 L的圓形養殖桶中,12—35日齡期間每天投喂3—5次, 35日齡后每日投喂2次, 每天清理殘餌糞便并及時換注清水。所有實驗魚65日齡后投喂未經處理的黃顙魚商品飼料,利用加熱棒將水溫維持在(26±2)℃。越冬期間(12月至次年2月)自然降溫, 水溫隨自然水溫波動,之后利用加熱棒將水溫逐步升至(26±2)℃。此后,在300日齡至330日齡期間, 利用黃顙魚性別特異分子標記, 將XX與XY遺傳型個體分開, 轉移至1600 L圓形養殖桶飼養, 直至性成熟。
1.2 XX偽雄魚與XX雌魚人工繁殖及其子代苗種培育
在黃顙魚繁殖季節, 挑選規格較大、腹部不膨大的XX偽雄魚作為后備父本; 解剖后將性腺中有乳白色精巢的個體作為繁殖父本。為了避免雌性親本中Y染色體的摻入, 我們對雌性親本進行逐尾遺傳性別鑒定, 將XY雌魚篩選剔除, 保證雌性親本基因型全部為XX。解剖上述獲得的XX偽雄魚, 取乳白色、發育良好的精巢組織進行碾磨; 將達到效應時間的XX雌魚進行人工擠卵, 按照1.0 g精巢組織與1.5 kg魚卵的比例人工授精, 在0.3%鹽水條件下混合受精, 加滑石粉脫黏, 轉入流水孵化桶進行孵化。受精12h后統計受精率(受精卵數/總卵粒數×100%), 出膜后估算孵化率(孵出仔魚數/受精卵數×100%)。
將出膜后的XX全雌魚水花轉移至水泥池, 暫養3d, 待魚苗平游后轉移至面積為4000 m2、深2.0 m的池塘中進行苗種培育, 下塘7d開始馴食人工配合飼料, 下塘20d時投喂黃顙魚商品飼料, 開展大規格苗種培育及成魚養殖。
1.3 遺傳型性別鑒定
普通黃顙魚雌雄親本及其子代、偽雄魚及與其繁殖的雌性親本均利用黃顙魚性別特異性分子標記進行遺傳型性別鑒定。采用異丙醇沉淀法快速提取背鰭DNA[17]。利用Dan等[12]研發的性別特異分子標記(XY-F: 5′-GATTGTAGAAGCCATCT CCTTAGCGTA-3′; XY-R: 5′-CATGTAGATCACTG TACAATCCCG-3′)鑒定每尾魚的遺傳型性別。PCR反應體系為10 μL: 上下游引物各0.5 μL; 模板DNA 1.0 μL; 2×TaqPCR Master Mix 5.0 μL(康為世紀); ddH2O 3.0 μL。PCR反應程序為: 94℃預變性3min, 34個循環(94℃變性30s, 59℃退火30s, 72℃延伸40s), 72℃延伸5min。擴增完成后, 取PCR產物3 μL, 以DL2000 marker(康為世紀)作為參照, 利用凝膠成像系統依據片段長度鑒別出XX(一條帶,955 bp)與XY(2條帶, 分別為826和955 bp)基因型。
1.4 表型性別鑒定與性腺組織學觀察
取實驗組XX基因型個體繁殖季節(300日齡)性腺組織, 從外觀上區分雌性、雄性與間性(性腺中同時具有精巢和卵巢組織的個體), 雄性與雌性組織分別取精巢、卵巢部分, 間性性腺組織取卵巢、精巢與連接部分; 隨機取池塘養殖XX全雌魚60日齡和120日齡個體性腺組織, 保存于Bouin’s液中固定, 48h后轉入70%乙醇常溫保存。性腺組織學處理及樣品拍攝參照齊飄飄的處理方法[18], 樣品通過梯度乙醇脫水、二甲苯透明和石蠟包埋后, 連續切片(切片厚度5—6 μm), 用蘇木精-伊紅染色(HE染色), 中性樹膠封片, 自然晾干。使用連接成像系統(MShot Image Analysis System)的光學顯微鏡進行性腺組織的觀察與拍攝, 結合性腺外觀, 鑒定個體表型性別, 判斷性腺發育狀態。各發育階段性腺中生殖細胞的鑒別參照本實驗室前期對黃顙魚性別分化及性腺發育的組織學觀察研究[6,9,13—15]。
1.5 XX偽雄魚與XY雄魚精子活力比較分析
麻醉后解剖XX偽雄魚與對照組XY雄魚, 取精巢, 將其剪開后收集精液樣本, 用Hank’s平衡液稀釋。XX偽雄魚與XY雄魚各取3尾, 每尾魚的精液樣本進行3次技術性重復。通過計算機輔助精子分析系統(Computer-Aided Sperm Analysis, CASA)對精子活力進行檢測, 拍攝精子運動過程, 統計第2s、4s、6s、8s、10s和12s時3個常用的精子活力運動參數, 即平均路徑速度(Average Path Velocity,VAP)、直線運動速度(Velocity Straight Line, VSL)和曲線運動速度(Curvilinear Velocity, VCL), 記錄每次運動中不同運動類型精子的比例。
1.6 數據分析
實驗數據采用SPSS (IBM SPSS Statistics 26)統計軟件進行統計學分析, 用單因素方差分析(Oneway ANOVA)進行體指標和精子運動參數的差異顯著性檢驗, 用t檢驗進行受精率和孵化率的差異顯著性檢驗, 分析前進行數據正態性和方差齊性檢驗, 利用Duncan’s多重比較確定組間差異的顯著性,P<0.05時表示差異具有統計學意義。文中的數據均以平均值±標準誤(mean±SE)表示。
2 結果
2.1 LZ、MT與高溫共同處理對XX個體雄性化的影響
通過性別特異分子標記、性腺外部形態與組織學, 我們將實驗處理組XX遺傳型中性別發生逆轉和具備繁殖能力的個體分為三種表型, 即完全逆轉型(性腺完全逆轉為精巢, XX偽雄魚)、多精少卵型(XX間性個體, 精巢面積≥50%)與少精多卵型(XX間性個體, 精巢面積<50%)。XX偽雄魚的性腺指數(GSI)顯著低于XY雄魚(P<0.05), 但三種XX偽雄魚之間GSI差異不顯著(表1)。實驗處理組XY與XX遺傳型比例分別為48.0%和52.0%, 其中完全逆轉型XX比例為8.0%, XX間性個體比例為14.0%。對照組XY與XX遺傳型比例分別為46.0%與54.0%, 沒有觀察到性別發生逆轉或性腺發育異常的個體。
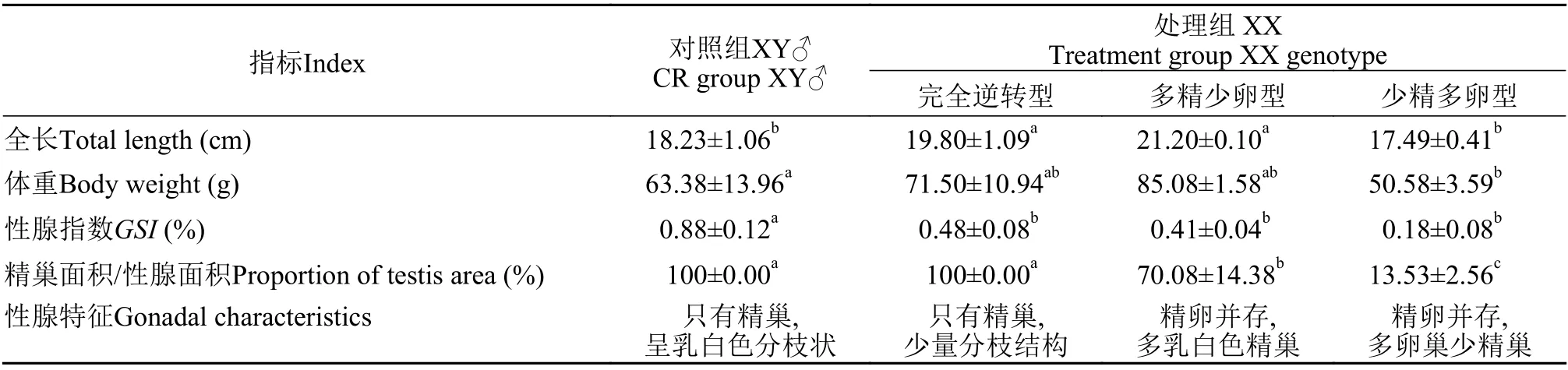
表1 黃顙魚性別逆轉XX個體與XY雄魚體指標比較分析Tab.1 Comparative analysis of body index of XX pseudo-male and XY male
黃顙魚繁殖季節期間, 對照組XY雄魚精巢發育至Ⅳ期(圖1A), 呈乳白色, 具有鯰形目魚類精巢典型的分枝狀結構, 分枝狀結構中精液充盈, 剪開精巢有白色精液流出; 通過組織切片可觀察到大量精小囊結構, 其中存在大量精子細胞(圖1A1和圖1A1′)。完全性逆轉的XX偽雄魚精巢發育稍滯后, 精巢分枝結構較少, 發育至Ⅳ期, 乳白色精巢組織中有精液存在(圖1B), 通過組織學切片觀察到精巢中存在大量成熟精子和不同發育時期的精原細胞, 精小葉結構異常(圖1B1和圖1B1′)。兩種類型XX間性個體中(多精少卵型和少精多卵型), 精巢部分有少量分枝狀結構, 剪開精巢有白色精液流出,通過組織學切片觀察到精巢結構中存在大量成熟精子; 卵巢部分發育正常, 肉眼可見成熟卵粒, 卵母細胞中觀察到大量卵黃顆粒(圖1C和圖1D)。
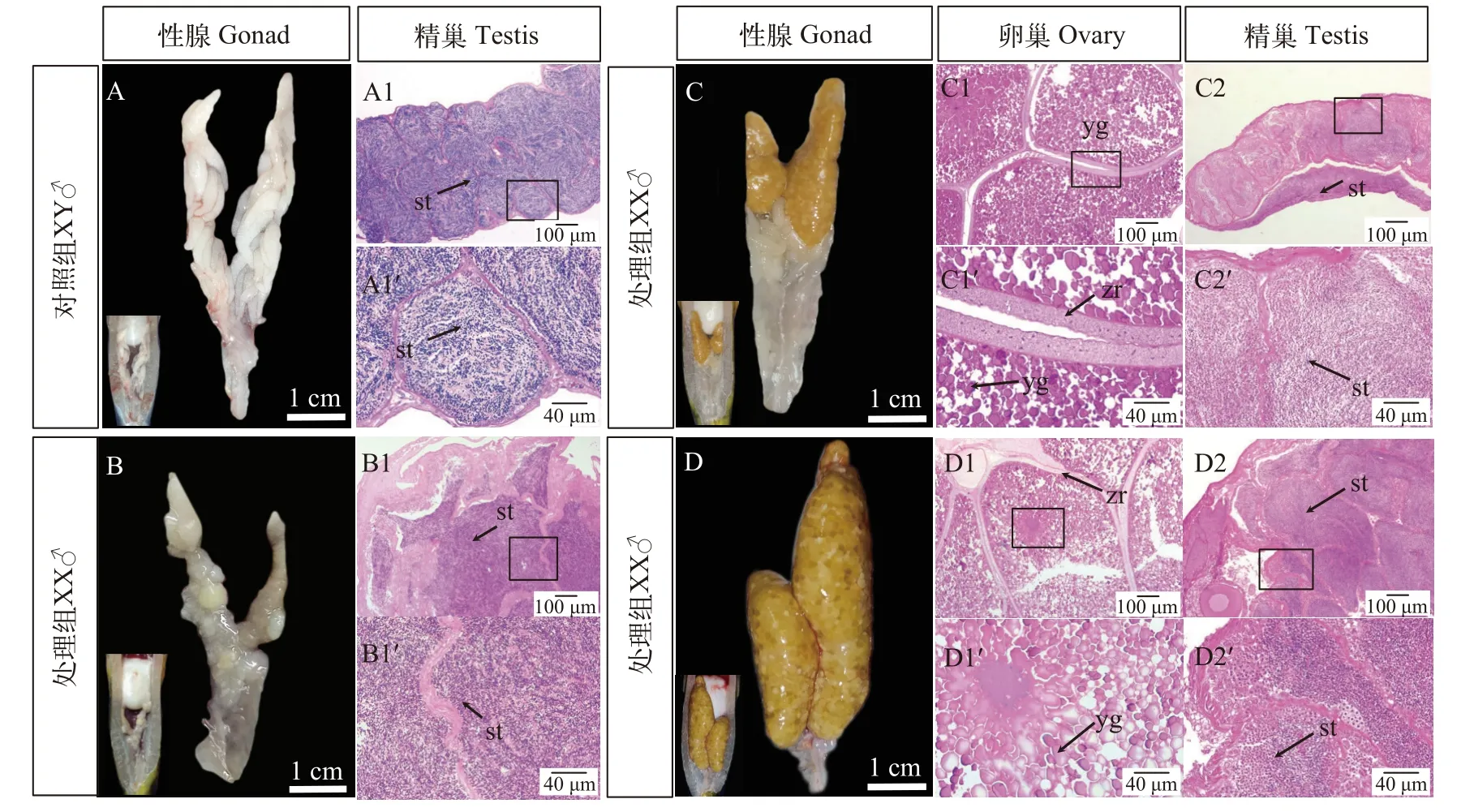
圖1 黃顙魚XX偽雄魚與XY雄魚性腺形態及組織學比較Fig.1 Gonadal morphology and histology of XX male and XY male in yellow catfish
2.2 XX偽雄魚生殖能力鑒定
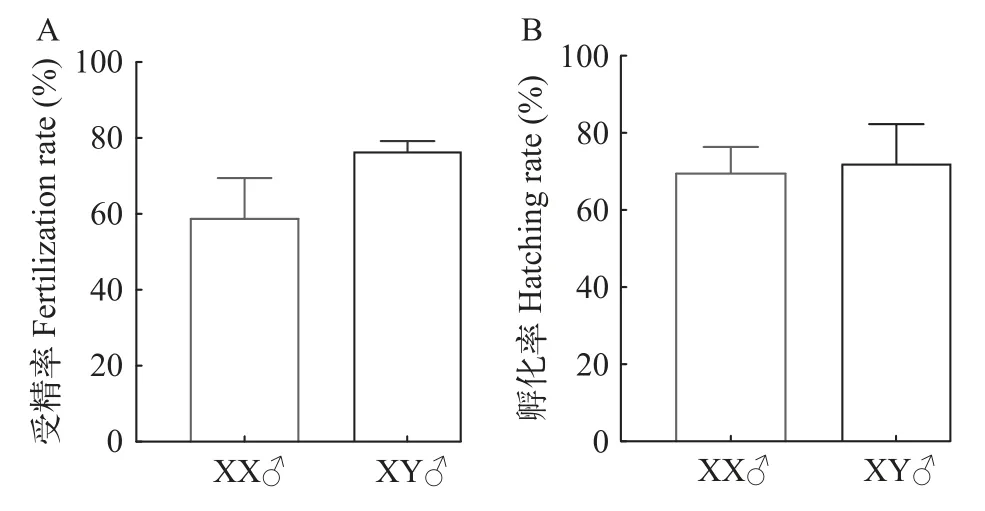
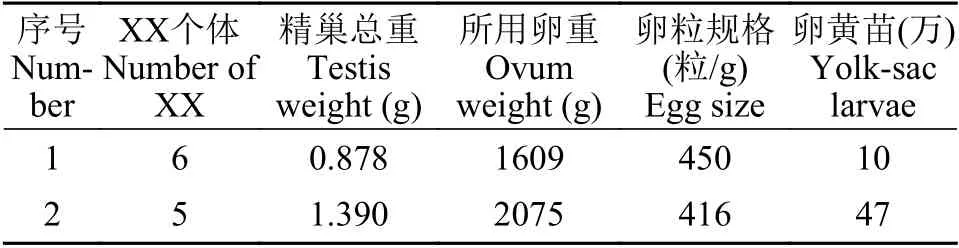
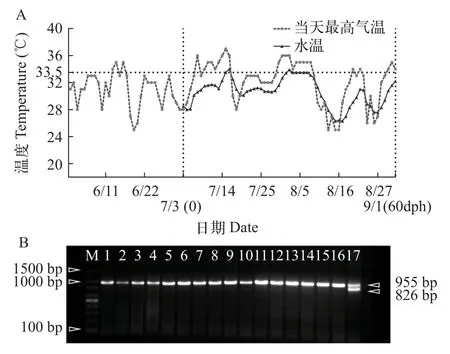
為了評估XX雄魚的生殖能力, 我們對發生逆轉的XX個體與XY雄魚的精子活力進行了比較分析(圖2)。結果顯示, 完全逆轉型的XX雄魚與XY雄魚在不同精子運動類型、VAP(平均路徑速度, Average Path Velocity)、VSL(直線運動速度, Straight Line Velocity)和VCL(曲線運動速度, Curvilinear Velocity)方面均無顯著性差異(P>0.05)。XY雄魚與完全逆轉型XX雄魚的運動型精子比例顯著高于少精多卵型XX個體 (P<0.05)。前進型精子[19](Progressive, 精子運動參數符合VAP>50 μm/s, 前進型ASL/VAP>80%)中, 完全逆轉型XX雄魚顯著高于少精多卵型XX個體, 而慢速型運動精子[19](Slow, 精子運動參數符合0 圖2 黃顙魚XX偽雄魚與XY雄魚精子活力比較分析Fig.2 Comparative analysis of sperm motility between XX pseudo-male and XY male in yellow catfish 利用上述具有繁殖能力的XX偽雄魚共開展了兩個批次的全雌黃顙魚規模化繁育工作, 共孵出57萬尾黃顙魚仔魚(表2)。以XX偽雄魚為父本的人工繁殖, 其受精率和孵化率平均分別為58.8%與69.6%, 與正常XY雄魚為父本的人工繁殖相比, 受精率與孵化率在統計學上均無顯著性差異 (圖3A和圖3B)。 圖3 黃顙魚XX偽雄魚和XY雄魚作為父本的人工繁殖受精率與孵化率比較Fig.3 Comparison of fertilization rate and hatching rate between XX pseudo-male and XY male as male parents in yellow catfish 表2 全雌黃顙魚規模化繁殖明細Tab.2 Breeding details of the all-female yellow catfish population 為觀察夏季自然高溫條件下全雌黃顙魚種群性別分化和性腺發育情況, 我們記錄了培育池塘6—9月份每日最高氣溫, 及幼魚0至60日齡期間池塘水深1m處的水溫。圖4A顯示, 在黃顙魚0—60日齡期間, 池塘平均水溫為30.6℃, 有8d時間水溫超過33.5℃。在幼魚60日齡和120日齡時, 我們對隨機采樣所獲的共計172尾幼魚進行了遺傳型性別鑒定, 部分結果如圖4B所示, 所有幼魚基因型性別均為XX(圖4B)。 圖4 池塘養殖溫度記錄及全雌黃顙魚種群基因型性別鑒定Fig.4 Pond temperature records and genotypic sex identification of the all-female yellow catfish population 對全雌黃顙魚幼魚60日齡和120日齡分別進行了取樣觀察, 發現在60日齡時, 97.2%的個體性腺發育正常, 呈透明棒狀, 組織切片觀察可發現大量初級卵母細胞, 細胞核與核仁明顯(圖5)。剩余2.8%的XX個體發生了性逆轉, 精巢組織中可見精原細胞和初級精母細胞, 也能觀察到精小囊結構。至120日齡時, 100尾幼魚中88.0%的個體卵巢發育正常, 部分個體可觀察到黃色卵粒, 組織學結果顯示,卵母細胞中充滿卵黃顆粒, 處在卵黃積累期。剩余12.0%的XX個體發生了性逆轉, 其中少部分個體精巢呈典型的分枝狀, 組織學觀察到大量的精小囊結構, 精巢中觀察到精原細胞、精母細胞及非常少量稀疏的精子細胞, 精巢發育異常; 大部分個體精巢呈透明狀、無分枝, 組織學觀察到精小囊結構異常,并且存在少量卵母細胞。黃顙魚幼魚60日齡時,XX雌魚全長與體重顯著低于XX雄魚, 但養殖至120日齡時, 兩者差異不顯著(圖5)。 為了解決全國黃顙魚雌性親本短缺問題, 我們利用XX雄魚與XX雌魚進行全雌黃顙魚規模化繁育研究, 獲得了57萬尾基因型全部為XX的黃顙魚仔魚, 并將其成功培育成大規格魚種, 研究結果同時將為雜交黃顙魚的遺傳育種提供豐富的選育材料。 魚類的性別具有很強的可塑性, 性別分化關鍵時期易受到外源激素, 包括雌激素、雄激素、性激素合成酶抑制劑、性類固醇受體拮抗劑及具有類似功能的物質的影響, 也易受到環境應激條件, 包括應激性溫度(包括高溫與低溫)、高密度、低溶氧和環境背景色等的影響[3,4,6,20]。據不完全統計, 為了直接或間接獲得單性種群, 全世界有超過27種性逆轉藥物在超過100種魚類中使用, 其中在多種經濟魚類中都成功獲得了單性種群[4]。黃顙魚的性別決定方式主要是遺傳型性別決定, 性別決定類型為雄性個體異形, 即XX/XY[21], 同時明顯受到高溫等極端環境因素的影響[6]。因此, 生產全雌黃顙魚的關鍵在于成功誘導XX個體雄性化, 以及性逆轉XX雄魚具備正常繁殖能力。 早在2013年, Shen等[14]首次報道了一種芳香化酶抑制劑LZ(Letrozole, 來曲唑)在黃顙魚性別分化關鍵時期(10—59日齡)能顯著提高群體雄性率, 并且雄性率與LZ濃度呈現正相關關系。隨后, Shen等[15]又報道了人工合成的雄激素MT(17α-methyltestosterone, 17α-甲基睪丸酮)并不能顯著提高黃顙魚雄性率, 但是誘導了大量間性個體的產生, 精巢呈現大量空腔結構, 生殖細胞稀疏。在這兩項研究開展過程中, 黃顙魚的性別特異分子標記還未開發, 只能通過性別比例和組織學初步判斷性逆轉的效果。此后, 楊天毅等[16]也嘗試了用MT和LZ對黃顙魚幼魚進行處理, 得到了相似的結論。結合黃顙魚性別特異分子標記[12], 楊天毅等[16]發現, 各濃度MT處理的XX個體性腺為空腔狀精小囊結構, 沒有精子生成, 而LZ能將XX個體誘導為正常XX雄魚,具有較好繁殖能力。此外, Yu等[6]發現, 黃顙魚性別分化關鍵時期經3.5周的高溫處理(33.5℃)能誘導39.1%的XX個體雄性化, 但大部分XX雄魚精巢發育異常, 未觀察到精小囊結構, 推測大部分個體不具備生殖能力。但是溫度處理作為一種環境友好型的性逆轉方法, 為魚類性別控制提供了新的思路,有望建立環境友好型性別控制方法。 因此, 基于以上前期研究結果, 本實驗利用三種雄性化因素(包括MT、LZ和高溫)進行聯合處理, 獲得了完全逆轉型XX和XX間性個體, 其中完全逆轉型XX占XX基因型的8.0%, 比例偏低, 但其精巢發育較好, 組織學切片能觀察到大量精子細胞, 推測其具有繁殖能力。到目前為止, 所有研究表明, 用激素單獨處理或聯合處理誘導黃顙魚XX個體性逆轉并產生具有正常繁殖能力的XX雄魚比例仍然偏低, 這可能與黃顙魚遺傳型性別決定機制較穩定有關。楊天毅等[16]利用LZ處理黃顙魚幼魚, 發現1000 mg/kg處理組正常發育精巢比例為35.7%, 這個處理劑量對于魚類性逆轉的研究來說是非常高的處理濃度[8]。此外, 上述提到, Yu等[6]利用高溫誘導了39.1%的XX個體雄性化, 但性成熟階段時, 絕大部分XX雄魚精巢發育異常。這些結果表明, 黃顙魚雌性性別決定基因非常強大, 大部分個體都對雄性化因素處理有抵抗作用, 即便本研究將3種雄性化因素聯合進行處理, 完全性逆轉的比例也不高。需要特別指出的是, MT作為一種人工合成的雄激素, 可能不適合誘導黃顙魚XX個體雄性化。這在多數魚類中都有相似報道, 包括大西洋鮭(Salmo salar)[22]、石斑魚(Epinephelus coioides)[23]和雌核發育的鯽[24]等, 可能是因為MT在魚體內代謝過程中被芳構化, 從而產生雌激素和雌性化效應[25,26], 也可能是因為外源雄激素的應用導致內源雄激素合成減少[25,27]。因此應當將MT替換成不能被芳構化的雄激素, 比如17α-methyldihydrotestosterone。后續研究需要同時考慮降低雌性分化相關基因表達或雌激素水平, 同時提高雄性分化相關基因表達或雄激素水平, 從而提高XX完全雄性化效率。 黃顙魚性別特異分子標記未報道時, Shen等[14,15]發現LZ處理可以增加黃顙魚群體雄性比例, 推測有XX遺傳型個體成功性逆轉為XX偽雄魚; 此后, 齊飄飄等[13]借助性別特異性分子標記, 發現黃顙魚在高溫或皮質醇處理后, 部分XX偽雄魚表現出雄性第二性征, 性腺出現典型精小葉結構, 少數能觀察到精子細胞。楊天毅等[16]利用高濃度LZ投喂黃顙魚可誘導XX個體雄性化, 部分XX偽雄魚具備繁殖能力。以上研究為黃顙魚的雄性化奠定了重要基礎, 但未曾對性逆轉所得XX偽雄魚的繁殖能力做出評估。 精子運動速度、持續時間和運動類型是判斷精液質量和受精率高低的主要參數[28], 運動精子和前進型運動精子百分比可以說明精子樣本質量總體概況[29]。Senior等[30]對激素誘導的偽雄魚(基因型雌魚)和正常雄魚之間精子特征進行了系統綜述,經大量數據分析后發現, 偽雄魚與正常雄魚在排精量、運動精子百分比或持續時間等方面無顯著差異, 推測在不同性別決定類型魚類中, 偽雄魚可能具備與遺傳型雄魚相當的繁殖能力, 這對于全雌種群生產至關重要。例如, 七帶石斑魚(Epinephelus septemfasciatus)和橙點石斑魚(Epinephelus Coioides)在MT誘導性逆轉后, 運動精子百分比與對照雄魚無異, 精子濃度相似, 最終受精率與孵化率也無統計學差異[31—34]。本實驗結果與前人描述相似,完全性逆轉的XX偽雄魚與XY雄魚不同精子運動類型間無統計學差異(P>0.05), XX偽雄魚的完全逆轉型中, 前進型精子運動能力甚至略高于XY雄魚(圖2), 說明黃顙魚偽雄魚可能與遺傳型雄魚的生殖性能相當[30]。需要特別指出的是, 本實驗中,激素誘導的性逆轉個體中存在大量XX間性個體,這表明誘導效率未達到理想狀態, 但是這些個體的精巢部分也具備授精能力。總之, 開展全雌黃顙魚規模化繁育的核心是如何高效獲得完全性逆轉的,具備授精能力的XX偽雄魚。 黃顙魚2020年全國產量達56.5×107kg, 屬于小型魚類, 產量的持續增長對親本數量的需求不斷增長。黃顙魚屬于分批產卵魚類, 為了提高產卵率和產卵量, 通常催產劑的劑量較其他魚類高, 且腹部擠壓力度較大, 對雌性親本損傷大, 死亡率達50%以上, 即便是操作規范的繁殖場, 母本催產后死亡率也通常高達30%。加上全雄黃顙魚和雜交黃顙魚養殖的普及, 雌性親本嚴重缺乏。按照當前黃顙魚產量和苗種存活率估算, 全國每年黃顙魚母本需求量達1×106kg。因此, 開展全雌黃顙魚規模化繁育對黃顙魚產業健康可持續發展意義深遠, 也對黃顙魚新品種選育至關重要。 本研究首次利用MT、LZ和高溫誘導的XX偽雄魚為父本開展全雌黃顙魚規模化繁育, 其受精率和孵化率分別為58.8%與69.6%, 獲得了基因型為XX的全雌黃顙魚仔魚57萬尾。全雌黃顙魚苗種在池塘中生長情況良好, 對60日齡和120日齡共計172尾魚隨機取樣分析發現, 92%XX雌魚性腺發育正常。120日齡時, 卵巢已發育至卵母細胞成熟期,卵巢呈黃棕色, 肉眼可見大量顆粒狀卵粒(圖5C)。組織學結果顯示, XX雌魚的卵粒中卵黃物質持續增加, 卵黃顆粒相互融合。這一卵巢發育水平比齊飄飄等[13]在黃顙魚雌魚120日齡時觀察到的初級卵母細胞發育更快, 與王凌宇等[9]報道的300日齡黃顙魚卵巢呈現的結果相似, 預計在良好池塘培育條件下, 該批次全雌黃顙魚中較大個體在來年繁殖季節能作為母本開展繁殖和選育工作。以上結果表明,通過性逆轉的XX偽雄魚與正常XX雌魚進行人工繁殖, 可實現全雌黃顙魚的規模化生產。本研究結果為全雌黃顙魚繁育體系的建立提供了重要基礎。 在本研究中, 由于XX偽雄魚在室內養殖系統中養殖, 生長速度較池塘慢, 導致全雌黃顙魚繁育工作延后, 水花下塘時正值湖北地區高溫期, 全雌黃顙魚性別分化期間(12—45日齡)水溫較高。在60日齡和120日齡時取樣發現, 分別有2.8%和12.0%的個體發生了不同程度的雄性化, 與此前Yu等[6]報道的33.5℃高溫誘導XX個體性逆轉相似,性逆轉個體的組織學結構也與Yu等[6]報道的極為相似。在全雌黃顙魚0—60日齡期間, 池塘平均水溫為30.6℃, 期間有8d時間水溫超過33.5℃, 推測黃顙魚幼魚受高溫影響而發生性逆轉。因此, 要獲得基因型和遺傳型為雌魚的全雌黃顙魚群體, 應當避免池塘高溫期間開展苗種培育。另一方面, 也可能是XX偽雄魚后代性逆轉的臨界溫度閾值降低, 從而在池塘高溫條件下發生了雄性化。在羅非魚和半滑舌鰨(Cynoglossus semilaevis)中, 與遺傳雄性的后代相比, 由溫度反轉偽雄魚繁殖的后代在高溫下表現出更高的轉雄率, 其后代在常溫狀態下也可能會發生自然性逆轉[35,36]。當然, 也可能是兩方面共同作用的結果。因此需要更深入研究XX偽雄魚子代發生性逆轉的遺傳學與表觀遺傳學機制, 從而更有效地開展全雌黃顙魚繁育工作。 從另一方面來講, 溫度誘導的雄性化, 為環境友好型性別控制技術體系的建立提供了基礎。全世界重要經濟魚類的單性種群生產體系中, 絕大部分要使用性類固醇激素或類似物, 其殘留或代謝產物對自然環境存在潛在的負面影響, 這種生產方式不符合水產養殖綠色發展的大方向。因此, 環保型單性種群的生產方法亟待建立。后續我們將進一步研究池塘高溫培育條件下產生的XX偽雄魚, 分析其繁殖能力, 為黃顙魚環境友好型性別控制技術體系的建立提供依據。
2.3 全雌黃顙魚規模化繁育
3 討論
3.1 黃顙魚XX遺傳型個體雄性化
3.2 XX偽雄魚個體的繁殖能力
3.3 XX全雌黃顙魚規模化繁育
3.4 環境應激對黃顙魚全雌種群性別分化的影響
