基于線粒體CO I 和Cyt b 序列的丹江口水庫鳙群體遺傳結構分析
2022-12-27 04:12:32汪鄂洲付昕石義付陳澤濤朱勇夫董薇薇鄭海濤4闕延福李偉濤朱麗婭
漁業致富指南 2022年11期
○汪鄂洲付昕石義付陳澤濤朱勇夫董薇薇鄭海濤,4闕延福李偉濤朱麗婭
(1.水利部水工程生態效應與生態修復重點實驗室,水利部中國科學院水工程生態研究所 湖北武漢 430079;2.南水北調中線水源有限責任公司 湖北丹江口 442700;3.湖北省水產科學研究所 湖北武漢 430071;4.湖北中水長江生態保護研究院有限公司 湖北武漢 430014)
鳙(Aristichthys nobilis)屬鯉形目(Cypriniformes)、鯉科(Cprinidae)、鰱亞科(Hypophthalmichthyinae),是我國傳統的“四大家魚”之一,其自然分布區主要在中國大陸長江和珠江流域,其他水體中也有分布。自二十世紀五十年代我國突破四大家魚人工繁殖技術后,鳙逐漸成為我國乃至世界主要淡水魚類養殖種類。世界上已有20多個國家和地區引進了鰱和鳙,或作為水產養殖對象供人們食用,或用于池塘、水庫等水體浮游生物的生物控制。根據聯合國糧農組織(FAO)的統計資料,2010年鳙為世界第七大水產養殖種類,根據我國2019年漁業年鑒統計數據,鳙的產量達到310.16萬噸,居養殖產量第三位。由于受水利建設,過度捕撈以及水體污染等多種因素影響,我國幾大天然水體中的鳙野生資源量呈急劇下降態勢。
丹江口水庫(110°34′47″-110°47′53″E,32°14′10″-32°58′10″N)位于漢江中上游,在湖北省丹江口市和河南省南陽市之間,水域橫跨鄂、豫兩省,是南水北調中線工程的唯一水源地,也是優化我國水資源配置的重大戰略工程。為進一步改善水庫水質,維護和恢復丹江口水庫水生生物資源,近20年來丹江口庫區多次開展包括“四大家魚”在內的經濟魚類增殖放流活動。僅2019年丹江口市放流鰱、鳙、草魚、鳊等魚苗4250萬尾。雖然大規模的增殖放流一定程度上對鳙資源的恢復起到作用,但放流苗種來源不一在很大程度上會對天然水域鳙群體帶來影響,甚至會導致物種野生群體遺傳多樣性降低,造成遺傳污染。因此,應加強增殖放流的遺傳管理,在放流前開展遺傳評估。馮曉婷等對2016-2018年長江江蘇段主要參與增殖放流的良種場所采集的鳙親魚和長江江蘇段各采樣點的回捕鳙進行評估,結果顯示增殖放流對長江江蘇段野生群體的貢獻率為6.11%。
遺傳多樣性不僅是物種多樣性和生態系統多樣性的基礎,也是魚類物種生存與進化的基礎,開展魚類遺傳多樣性研究,對漁業資源管理與養護、人工增殖放流等工作具有重要的指導意義。自上世紀九十年代以來,我國科研人員利用同工酶、微衛星、線粒體等不同的標記對長江流域鳙遺傳多樣性進行了研究,而有關丹江口庫區魚類的遺傳多樣性研究尚未見報道。丹江口庫區有關魚類的研究主要集中在庫區魚產力、魚類資源調查及魚類群落特征等方面。
本研究利用線粒體CO I和Cytb序列分析了丹江口水庫鳙增殖放流親本、苗種及庫區自然群體的遺傳多樣性,研究結果不僅是科學的漁業管理、制定保護策略依據,也將為鳙種質資源開發利用和增殖放流遺傳管理提供基礎支撐。
1 材料與方法
1.1 實驗材料
2019-2020年于丹江口增殖放流站隨機采集鳙親本(Yong-QB)和子代(Yong-ZD),收集丹江口庫區自然水體鳙樣品(Yong-KQ),共計收集鳙樣品92尾,具體信息見表1。每個樣品經過形態鑒定后,剪取適量尾鰭置于裝有無水乙醇的EP管中,帶回實驗室凍存于-20℃冰箱保存備用。
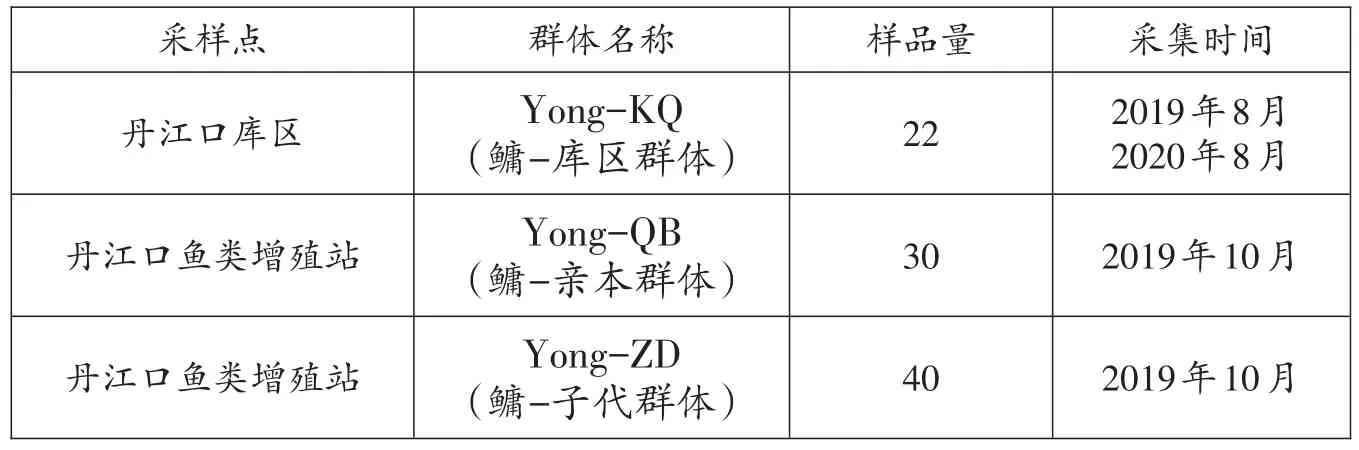
表1 鳙樣本信息
1.2 基因組總DNA提取及PCR擴增
鳙基因組總DNA采用酚/氯仿法提取,溶解于200 μL TE緩沖液中,檢查DNA樣品濃度后,保存于-20℃冰箱備用。DNA工作液稀釋為 50 ng/μL,PCR 反應體系為40 μL,其中Mix 20 μL、正反向引物各 2 μL、雙蒸水 14 μL、DNA模板2 μL。PCR擴增條件如下:94℃預變性 5 min,94℃變性 35 s,58℃退火 35 s,72℃延伸1 min,35個循環;72℃終延伸10 min。PCR產物送天一輝遠生物科技有限公司測序。鳙線粒體PCR擴增引物見表2。

表2 鳙CO I和Cytb序列擴增引物信息
1.3 數據處理
利用軟件FinchTV對測序得到的CO I和Cytb序列進行拼接,輔以人工校對,并與NCBI數據庫中已發表的鳙mtDNA序列進行對比,確保擴增CO I和Cytb序列的準確性。使用DnaSP 5.0軟件計算單倍型數目(N)、變異位點(V)、單倍型多樣性(Hd)、核苷酸多樣性(Pi)、平均核苷酸差異數(K)、基因流(Nm),以及單倍型類型等參數。使用Arlequin 3.01軟件,根據pairwise difference模型計算種群間的分化指數值(Fst)和分子方差分析(AMOVA)。使用MEGA V6.0計算堿基含量和轉換/顛換比率,根據Kimura雙參數模型計算單倍型間遺傳距離并進行NJ聚類。
2 結果
2.1 堿基組成
測序后經序列比對,共獲得3個鳙群體92條有效的線粒體CO I和92條有效的Cytb序列,長度分別為653 bp和1120 bp。基于CO I基因序列分析結果表明,堿基的平均含量分別為A=25.5%,T=29.3%,C=27.3%,G=17.9%;其中A+T的平均含量為54.8%,G+C為45.2%。92條鳙線粒體CO I序列中檢測到多態位點3個,其中簡約信息位點3個,單一變異位點0個,定義了4個單倍型,其中1個單倍型為3個群體共有單倍型,2個單倍型為2個群體共有,1個單倍型僅在親本群體中檢測到(表3)。
基于Cytb基因序列分析結果表明,堿基的平均含量分別為A=29.6%,T=27.2%,C=29.0%,G=14.2%,轉換顛換比為6.4;其中A+T的平均含量為56.8%,G+C為43.2%。92條鳙線粒體Cytb序列檢測到多態位點9個,其中簡約信息位點6個,單一變異位點3個,定義了8個單倍型,其中1個單倍型為3個群體共有單倍型,3個單倍型為2個群體共有,4個單倍型僅在單一群體中檢測到(表3)。
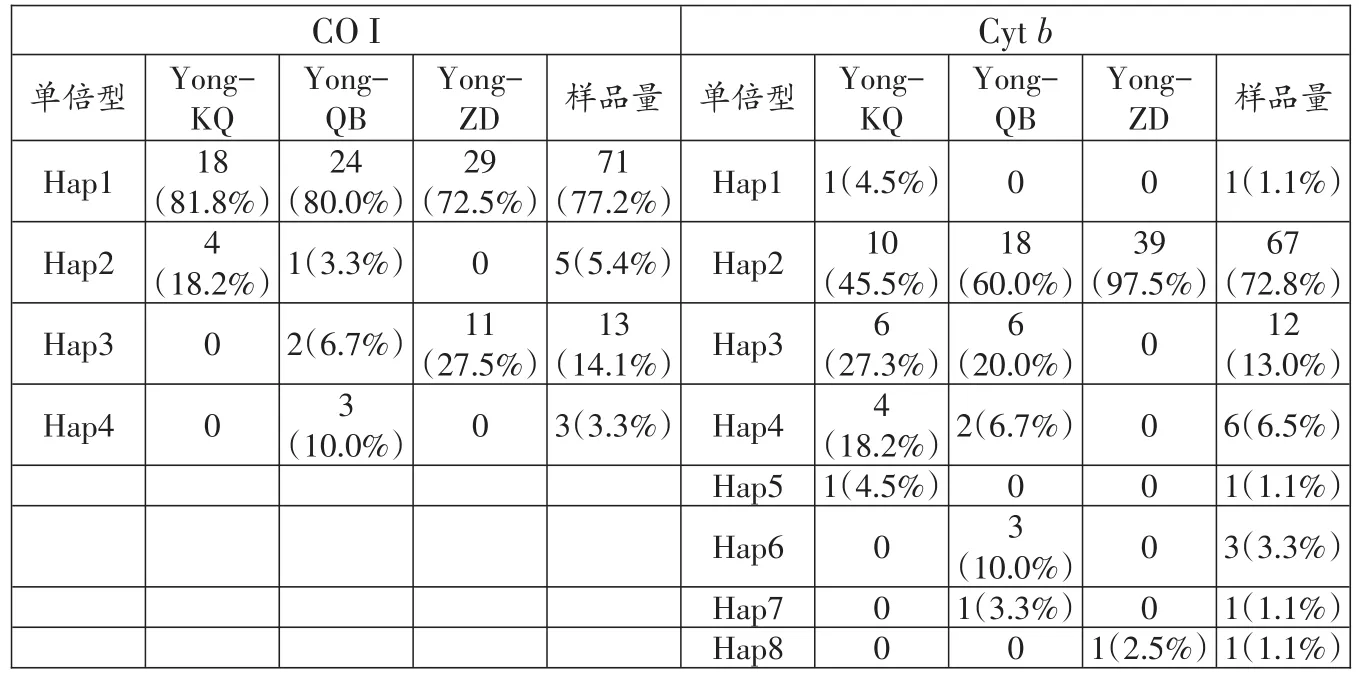
表3 鳙3個群體單倍型數目、類型、頻率及分布
2.2 群體遺傳多樣性
線粒體CO I序列分析結果顯示,3個鳙群體共定義了4個單倍型,其中Yong-KQ群體共22尾,定義了2個單倍型,其單倍型多樣性最低,僅為(0.312±0.106);Yong-QB群體共30尾,定義了4個單倍型,其單倍型多樣性為(0.356±0.106);Yong-ZD群體共40尾,定義了2個單倍型,其單倍型多樣性最高,為(0.409±0.065)。核苷酸多樣性和平均核苷酸差異分析結果顯示,Yong-ZD群體最高,分別為(0.00063±0.00011)、(0.40897);Yong-QB 群體次之,分別為(0.00059±0.00019)、(0.38161);Yong-KQ 群體最低,分別為(0.00048±0.00016)、(0.31169)。
線粒體Cytb序列分析結果顯示,3個鳙群體共定義了8個單倍型,其中Yong-KQ群體共22尾,定義了5個單倍型,其單倍型多樣性最高,為(0.714±0.065);Yong-QB群體共30尾,定義了5個單倍型,其單倍型多樣性為(0.605±0.016);Yong-ZD群體共40尾,定義了2個單倍型,其單倍型多樣性最低,僅有(0.050±0.047),此群體中有1個特有單倍型,可能是在丹江口增殖站運行初期,引進了部分苗種所導致。核苷酸多樣性和平均核苷酸差異分析結果顯示,Yong-ZD群體最低,分別為(0.00004±0.00004)、(0.05000);Yong-QB群體次之,分別 為(0.00158± 0.00027)、(1.76782);Yong-KQ群體最高,分別為(0.00159± 0.00022)、(1.78355)(表4)。
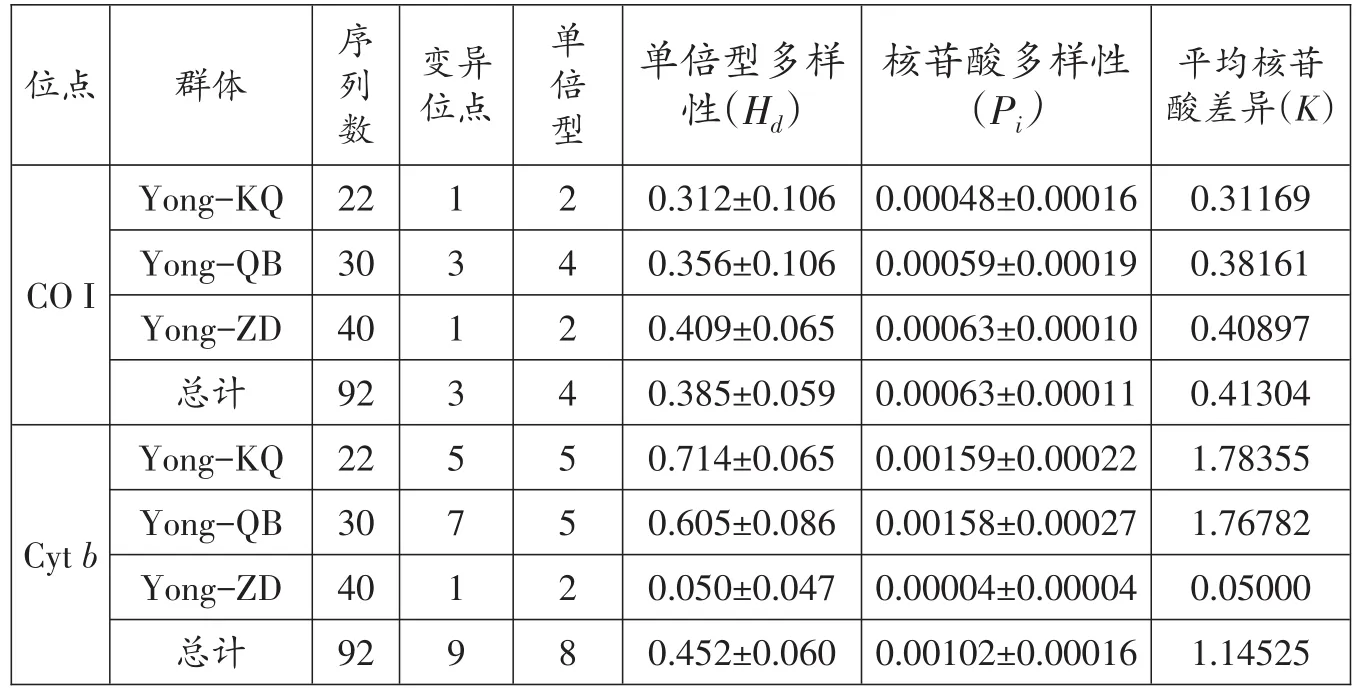
表4 鳙群體遺傳多樣性參數
2.3 群體遺傳差異
根據線粒體CO I和Cytb序列,采用Kimura雙參數(K2-P)模型分析鳙群體內和群體間遺傳距離,結果顯示群體內和群體間的遺傳距離差異不明顯(0.00048-0.00070,CO I;0.00004-0.00160,Cytb),群體內和群體間遺傳距離小(表5)。遺傳分化指數Fst和基因流Nm結果顯示,鳙群體間的Fst在 0.06785~0.20484,Yong-ZD和Yong-QB群體間P值小于0.05,Yong-KQ和Yong-ZD群體間P值小于 0.001,Nm在 4.29~30.26(基于CO I序列);Cytb分析結果顯示,Fst在 -0.00851~0.31930,Yong-ZD 和 Yong-KQ、Yong-QB群體間P值小于0.001,Nm在 0.89~186.66(表 6)。CO I分子方差分析(AMOVA)結果顯示,群體間的變異貢獻率占整個變異的12.60%,而群體內的變異貢獻率占87.40%;Cytb分析結果顯示,群體間的變異貢獻率占整個變異的15.94%,而群體內的變異貢獻率占84.06%(表7)。

表5 鳙群體內和群體間的遺傳距離

表6 鳙群體間的遺傳分化系數和基因流
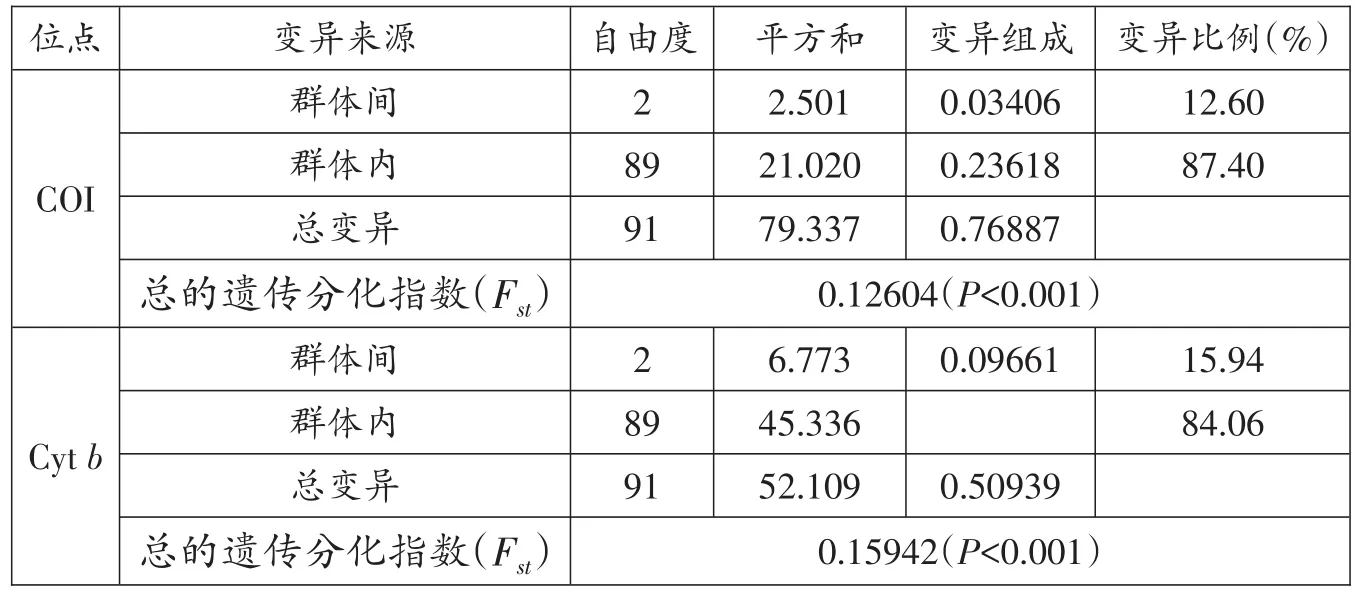
表7 鳙群體的分子方差分析
2.4 系統進化關系
基于 Kimura(K2-P)雙參數模型構建鳙群體CO I和Cytb單倍型序列系統發生NJ樹(圖1),系統發育樹各分支的置信度(bootstrap)均進行1000次重復檢驗。從CO I單倍型序列NJ樹可以看出,CO I-Yong-Hap-4單倍型是Yong-KQ群體特有單倍型。4個鳙單倍型譜系分為2支,Yong-KQ群體特有CO I-Yong-Hap-4單獨為一支,其余3個單倍型聚為另一支。
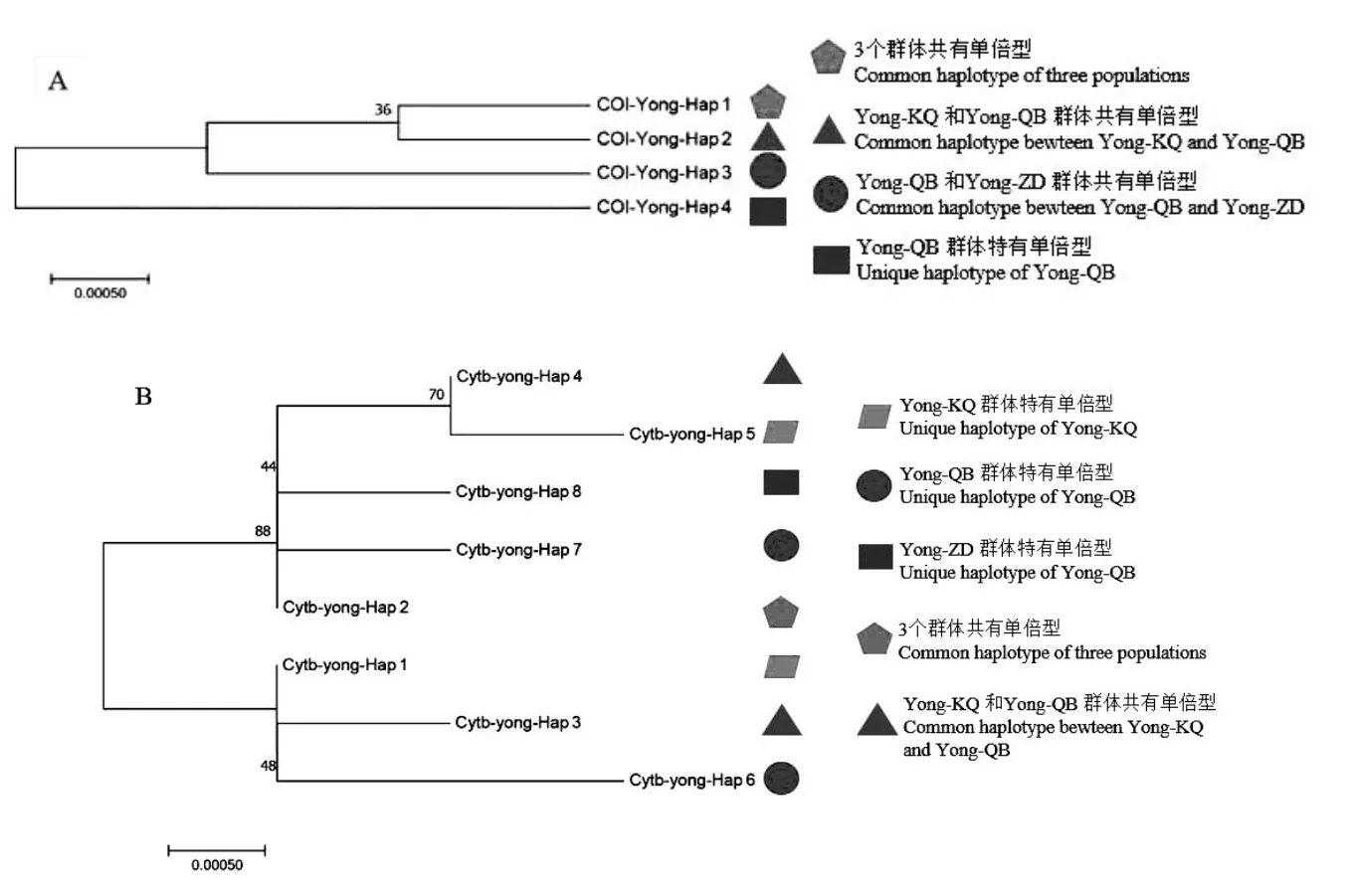
圖1 基于CO I(A)和Cytb(B)序列單倍型系統發育樹
Cytb單倍型序列NJ樹結果顯示,3個鳙群體同樣存在2個明顯的單倍型譜系分支,單倍型Cytb-Yong-Hap-2、Cytb-Yong-Hap-4、Cytb-Yong-Hap-5、Cytb-Yong-Hap-7和Cytb-Yong-Hap-8五個單倍型聚為一個分支,其余單倍型聚為另外一個分支,單倍型聚類關系中各群體單倍型散亂分布,3個鳙群體間并未形成明顯的地理譜系。
3 討論
3.1 群體遺傳多樣性
遺傳多樣性是生物多樣性的基礎和重要組成部分,遺傳多樣性每丟失10%,就會對生物繁育能力、存活率、生長等重要性狀產生很大的負面影響。單倍型多樣性(Hd)、核苷酸多樣性(Pi)和平均核苷酸差異數(K)是衡量種群遺傳變異的重要參數。本研究利用線粒體Cytb和CO I基因序列對丹江口增殖放流站鳙親本群體、子代群體和丹江口庫區自然群體進行了遺傳多樣性比較分析。基于線粒體Cytb基因序列研究結果顯示3個鳙群體的平均單倍型多樣性分別為0.452±0.060,平均核苷酸多樣性分別為0.00102±0.00016,平均核苷酸差異分別為1.14525,低于其他流域鳙群體遺傳多樣性。吳偉軍等利用Cytb基因序列研究紅水河鳙3個野生群體發現單倍型多樣性、核苷酸多樣性和平均核苷酸變異差異分別介于0.667-0.8929、0.0014-0.0027和1.5556-2.9286。基于線粒體CO I基因序列研究結果顯示3個鳙群體的平均單倍型多樣性分別為0.385±0.059,平均核苷酸多樣性分別為0.00063±0.00011,平均核苷酸差異分別為0.41304,低于其他增殖放流站鳙群體遺傳多樣性。劉慧芬等分析了河南省11個養殖場鳙群體的線粒體CO I基因序列,165個樣品共檢測出14個單倍型,平均單倍型多樣性和核苷酸多樣性分別為 0.86237、0.00223;Li等等通過線粒體D-Loop區和16S rRNA對比分析了長江(樣品采集于漢江和長江石首段)、珠江和黑龍江鳙群體的遺傳多樣性,發現長江鳙群體(Hd=0.787,Pi=0.0020)低于珠江群體和黑龍江群體;不同流域和增殖放流站鳙群體顯示出高單倍型多樣性和低核苷酸多樣性。沙航等利用微衛星標記分析了長江中游石首、監利和長沙3個鳙群體遺傳多樣性,結果顯示3個鳙群體具有較高的遺傳多樣性(PIC>0.5,He>0.6);馮曉婷等基于微衛星標記分析長江下游7個原良種場鳙親本和后備親本遺傳多樣性,發現鳙親本具有較高遺傳多樣性水平(PIC>0.789,He=0.889)。魚類養殖群體的遺傳多樣性低于野生群體,在鰱、鯉、草魚等養殖魚類中均有報道,在鳙養殖群體也有遺傳多樣性下降的報道。丹江口水庫3個鳙群體的線粒體DNA數據顯示了相對偏低單倍型多樣性和核苷酸多樣性,可能是以下因素有關。首先,養殖場親本在封閉的養殖條件下經過人工選擇和淘汰,這可能導致親本遺傳多樣性降低。其次,在采樣過程中,特別是鳙子代樣品,可能來自少數幾個親本的后代,表現出偏低的遺傳多樣性水平。再次,丹江口水庫建成之后,近20多年進行了多次人工放流,放流的苗種來源多樣,質量不一;如:有些繁殖場存在近親繁殖苗種的不科學操作,或往往幾條雌魚的產量就足以滿足當年魚苗的需求量,或在進行人工繁殖時所用的親本組合數較少,這就導致了后代群體的遺傳多樣性降低甚至出現生長性狀退化、抗逆性降低等不利于水產養殖業發展的現象。
3.2 群體遺傳結構
遺傳距離、遺傳分化指數(Fst)和基因流(Nm)是研究遺傳結構的重要參數。本研究中根據線粒體序列數據計算出的鳙種群內和種群間遺傳距離均<0.005,群體內和群體間遺傳距離差異不顯著,表明群體之間親緣關系很近。AMOVA分析顯示,有12.60%(CO I)、15.94%(Cytb)的變異來自于群體間,群體內的變異依然是主要來源;綜合線粒體序列數據分析發現群體間差異顯著(Fst=0.12604,Fst=0.15942,P<0.001)。一般認為,Fst值小于0.05時,表示群體間遺傳分化極小;當Fst值在0.05-0.15之間時,說明群體間有遺傳分化;當Fst值在0.15-0.25之間時,說明群體間遺傳分化較大;當Fst值大于0.25時,表明群體間有極大的遺傳分化。通過群體間的逐對比較進一步檢測鳙群體間可能的遺傳分化,Yong-ZD與Yong-KQ、Yong-QB之間Fst分別為 0.20484(P<0.001)、0.09214(P<0.05)(CO I)和 0.31930、0.22203(P<0.001)(Cytb),表明Yong-ZD與Yong-KQ群體之間有極大遺傳分化,與Yong-QB群體之間有顯著的遺傳分化;而Yong-KQ和Yong-QB群體之間遺傳分化不顯著。根據對鳙群體間基因流(Nm)的檢測結果,Yong-ZD與Yong-KQ群體之間有一定基因交流,而與Yong-QB群體之間有頻繁的基因交流(Nm>30),這也反應出親本與子代之間密切關系。從鳙群體間單倍型聚類關系分析可知,3個群體間均有共享單倍型,沒有聚集成不同群體,另外其他2個群體相比,Yong-ZD群體只有1個特有的單倍型,這也反應了Yong-ZD群體與其他群體間的遺傳分化。
3.3 鳙資源保護和利用
增殖放流是目前恢復漁業資源的主要手段之一,而放流后會不會導致魚類遺傳結構發生變化也是人們重點關注問題。本研究的結果表明,丹江口水庫3個鳙群體遺傳多樣性處于較低水平,子代群體與丹江口水庫自然群體有顯著的遺傳分化;親本群體與丹江口水庫自然群體之間有頻繁的基因交流,遺傳分化不顯著。漢江是長江主要支流,在丹江口水庫建成之前,分布于長江、漢江兩個水系的鳙能夠進行交流,丹江口水庫建成之后,阻斷與長江鳙交流通道。丹江口水庫大壩加高蓄水后,回水區上延、水流變緩,對分布于水庫上游至安康大壩江段的鳙及其他產漂流性卵的魚類有較大影響。近年來,為庫區魚類資源的保護和可持續發展,采取了包括禁漁期、增殖放流、打擊偷捕濫捕等措施。增殖放流活動在增加鳙資源量的同時,也可能改變了丹江口水庫鳙群體遺傳多樣性水平,因此,為了改善丹江口水庫天然種群的遺傳結構,建議加強增殖放流遺傳管理,選取來源于較高遺傳多樣性水平的長江鳙進行放流。
參考文獻略。
