連環湖河鱸的生長特性及資源評估
2023-01-05 04:44:48趙晨叢艷鋒王樂宋聃都雪張瀾瀾孫佳偉王慧博黃曉麗霍堂斌
水產學雜志 2022年6期
關鍵詞:生長
趙晨,叢艷鋒,王樂,宋聃,都雪,張瀾瀾,孫佳偉,4,王慧博,黃曉麗,霍堂斌
(1.中國水產科學研究院黑龍江水產研究所,農業農村部黑龍江流域漁業生態環境監測中心,寒地水域水生生物保護與生態修復重點實驗室,黑龍江 哈爾濱 150070;2.杜爾伯特蒙古族自治縣水產總站,黑龍江 大慶 166200;3.黑龍江省水產技術推廣總站,黑龍江 哈爾濱 150040;4.大連海洋大學,遼寧 大連 116023)
連環湖(46°30′~47°3′N,123°59′~124°15′E)是由烏裕爾河和雙陽河尾癌河水滯留形成的一組松嫩平原上的大型湖泊群[1],通過構造斷陷形成二八股泡、那什代泡、西葫蘆泡、火燒黑泡、東湖等18 個淺水湖泊[2]。連環湖魚類資源豐富,土著魚類以鯉(Cyprinus carpio)、銀鯽(Carassius auratus gibelio)、紅鰭原鲌(Culterichthys ergthropterus)、花(Hemibarbus maculatus)和烏鱧(Channa argus)等為主[3],為了優化漁業產業結構,人工移植鰱(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、草魚(Ctenopharyngodon idella)、河鱸(Perca fluviatilis)和大銀魚(Protosalanx hyalocranius)等魚類作為增養殖對象。河鱸屬鱸形目(Perciformes)、鱸科(Percidae)、鱸屬(Perca),為我國額爾齊斯河、烏倫古湖、博斯騰湖的主要經濟魚類[4]。由于其生長較快[5],且具有良好的環境溫度適應能力[6]和耐鹽性[7],作為經濟魚類從額爾齊斯河流域引入連環湖。肉食性魚類的引進對小型魚類的種群資源影響尤為明顯[8,9],自連環湖二八股泡首次引入河鱸后,河鱸便以較強的適應能力擴散至整個連環湖水域形成自然種群,但河鱸在地方市場認可度不高,且對連環湖主產經濟魚類—大銀魚存在威脅,如何減少河鱸種群資源量成為連環湖漁業生產中有待解決的難題。
常規的資源評估方法需要準確的種群生物學信息和可靠的漁獲量統計等數據[10],在數據有限情況下的漁業資源評估是國內主要研究的目標[11]。基于體長頻率分布對單一物種生長參數及資源數量變動特征的研究方法被廣泛應用于嘉陵江[12]、長江[13]、西江[14]、洱海[15]、竺山湖[16]、黃河[17]、東海[18]、南海[19]和黃海[20]等不同類型水域。但目前有關連環湖河鱸生物學參數及種群資源量評估的相關研究尚未開展,連環湖河鱸資源的管理和開發缺乏理論依據。本研究以受河鱸入侵問題困擾較為嚴重的二八股泡及那什代泡為例,根據2021 年河鱸生物學調查數據,探究其生長特性及資源利用狀況,可為連環湖河鱸資源的科學管理和有效防控提供理論參考。
1 材料與方法
1.1 樣品采集
2021 年6 月、8 月、10 月在連環湖二八股泡和那什代泡共設置6 個采樣點,具體位置見圖1。利用復合網目刺網(2 m×30 m),由12 種網目類型組成,每種網目的網長為2.5 m,網高2 m,網目規格依次 為8.5 cm、4.0 cm、12.5 cm、2.0 cm、11.0 cm、1.6 cm、2.5 cm、4.8 cm、3.1 cm、1.0 cm、7.5 cm 和6.0 cm,分別采集河鱸223 尾、244 尾,在新鮮狀態下,測量全長、體長、體質量,長度精確到1 mm,體質量精確到0.1 g。
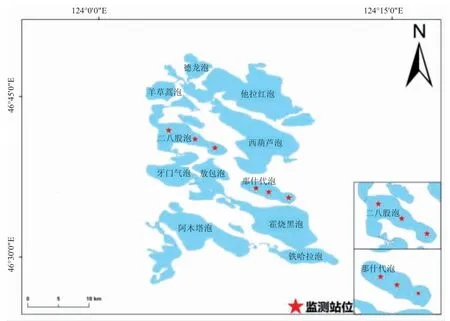
圖1 研究區域及采樣站點位置Fig.1 Location of study area and sampling sites
1.2 生長參數
用冪函數擬合體長—體質量的關系,其表達式為:

其中:W 為體質量(g);L 為體長(mm);a、b 為常數。a 為生長的條件因子,b 為異速生長指數。利用t 檢驗分析異速生長指數(b)和理論值(3)的差異性[21]。
利用Von Bertalanffy 生長方程擬合河鱸的生長特征[22],表達式為:
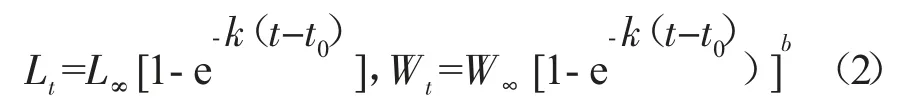
生長速度為生長方程的一階求導:
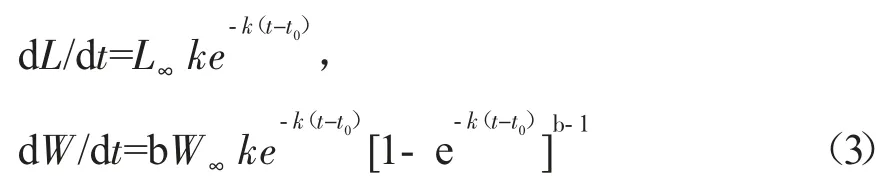
生長加速度為生長方程的二階求導:
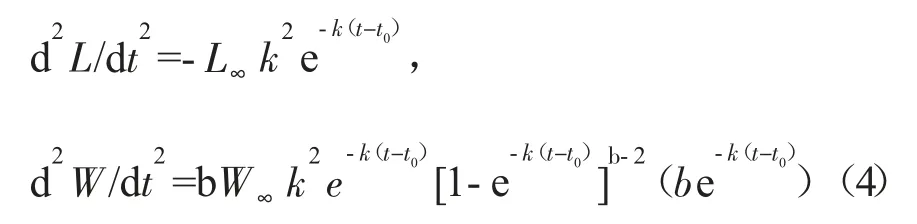
其中:t 為年齡,Lt和Wt分別為t 齡時的理論體長和體質量,L∞和W∞分別為漸近體長和漸近體質量,k 為生長系數,t0為理論初始年齡。
t0采用Pauly 的經驗公式[23]估算:
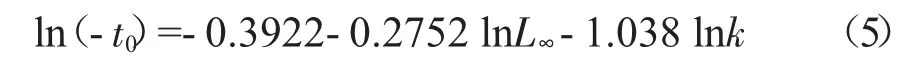
拐點年齡(ttp)、臨界年齡(tc)計算公式[22]為:

生長特征指數(Φ):
Φ=lg k+21g L∞
1.3 死亡系數與開發率
通過體長變換漁獲曲線法求得總死亡系數(Z)[24]:

式中Ntn為各體長組的樣本數占總樣本數比例,△tn為相對體長組下限生長到上限所需的時間。
通過Pauly 經驗公式[23]估算自然死亡系數(M):

式中T 為調查水域年平均水溫(℃),本文取12.5℃。
捕撈死亡系數(F)、資源開發率(E)及種群殘存率(S)表達式分別為:

1.4 資源量評估及利用分析
利用FISAT Ⅱ軟件的體長股分析法[24],評估河鱸的資源現存量,其表達式為:
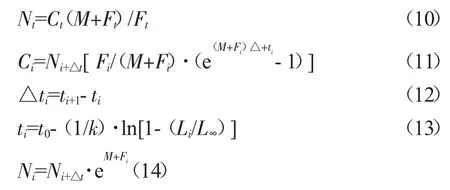
Nt為河鱸最大體長時的群體數量(ind.);Ct為河鱸最大體長時的漁獲量(t);Ft為河鱸最大體長時的捕撈死亡系數;Ci為河鱸第i 齡時的漁獲量(t);Ni、Ni+?t分別為河鱸處于第i 齡及第i+?t 齡時的資源數量(t);Fi為河鱸i 齡時的捕撈死亡系數。
利用Beverton-Holt 動態綜合模型,估算單位補充量漁獲量(Y’/R)和單位補充量生物量(B’/R)[25],對河鱸資源利用現狀進行分析,具體公式參見文獻[26]。
2 結果與分析
2.1 生長參數
2.1.1 體長、體質量組成及關系
連環湖二八股泡和那什代泡河鱸體長和體質量組成見表1。通過冪函數對二八股泡和那什代泡河鱸體長和體質量關系進行冪指數曲線擬合,其表達式分別為W=1×10-5L3.1271(R2=0.9683,n=223)、W=9×10-6L3.1598(R2=0.9919,n=244)(圖2)。異速生長指數b 與數值3 之間存在顯著差異(P<0.05),表明兩個湖泊河鱸均為異速生長。獨立樣本t 檢驗表明,兩個湖泊的河鱸體長與體質量無顯著性差異(P>0.05)。

圖2 二八股泡和那什代泡河鱸體長與體質量關系Fig.2 The length-weight relationships of perch Perca fluviatilis in Erbagu Lake and Nashidai Lake
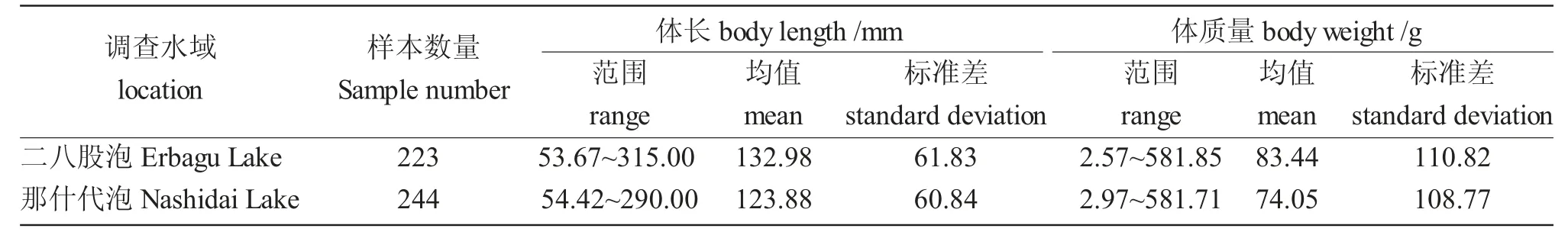
表1 二八股泡和那什代泡河鱸體長與體質量分布Tab.1 The body length and body weigh distribution of perch Perca fluviatilis in Erbagu Lake and Nashidai Lake
2.1.2 生長方程及生長速度
以10 mm 為間距劃分河鱸的長度頻率序列(圖3),用ELEFANⅠ方法求得二八股泡河鱸漸近體長L∞=409.8 mm,生長參數k=0.16,理論初始年齡t0=-0.8646,Von Bertalanffy 生長方程為:

圖3 二八股泡河鱸體長頻率時間序列及應用ELEFAN 估計的生長曲線Fig.3 Size frequency and body length growth curve estimated by ELEFAN in perch Perca fluviatilis in Erbagu Lake
Lt=409.8[1-e-0.16(t+0.8646)]
Wt=1478.34[1-e-0.16(t+0.8646)]3.1271
那什代泡河鱸漸近體長L∞=428.2 mm,生長參數k=0.16,理論初始年齡t0=-0.8542,Von Bertalanffy生長方程為(圖4):
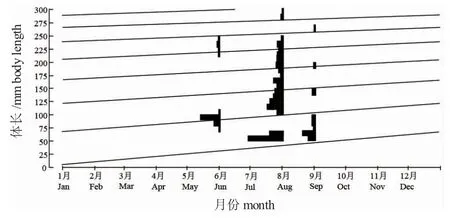
圖4 那什代泡河鱸體長頻率時間序列及應用ELEFAN 估計的生長曲線Fig.4 Size frequency and body length growth curve estimated by ELEFAN in perch Perca fluviatilis in Nashidai Lak
Lt=428.2[1-e-0.16(t+0.8542)]
Wt=1860.89[1-e-0.16(t+0.8542)]3.1598
對兩個湖泊河鱸的生長方程進行一階、二階求導,得到二八股泡河鱸的生長速度及生長加速度方程:
dL/dt=409.8×0.16e-0.16(t+0.8646)
dW/dt=3.1271×1478.34×0.16e-0.16(t+0.8646)[1-e-0.16(t+0.8646)]2.1271
d2L/dt2=-409.8×0.0256e-0.16(t+0.8646)
d2W/dt2=3.1271×1478.34×0.0256e-0.16(t+0.8646)[1-e-0.16(t+0.8646)]1.1271(3.1271e-0.16(t+0.8646)-1)
那什代泡河鱸的生長速度及生長加速度方程:
dL/dt=428.2×0.16e-0.16(t+0.8542)
dW/dt=3.1598×1860.89×0.16e-0.16(t+0.8542)[1-e-0.16(t+0.8542)]2.1598
d2L/dt2=-428.2×0.0256e-0.16(t+0.8542)
d2W/dt2=3.1598×1860.89×0.0256e-0.16(t+0.8542)[1-e-0.16(t+0.8542)]1.1598(3.1598e-0.16(t+0.8542)-1)
生長速度與年齡的關系表明(圖5),河鱸的體長生長速度不具有拐點,隨年齡的增加,生長速度逐漸遞減,體質量生長速度具有拐點;二八股泡河鱸拐點年齡ttp=6.26a,對應的體長、體質量為278.73 mm 和442.93 g;那什代泡河鱸拐點年齡ttp=6.34a,對應的體長、體質量為292.76 mm 和559.67 g。二八股泡、那什代泡河鱸的生長特征指數(Φ)分別為4.429 和4.467。
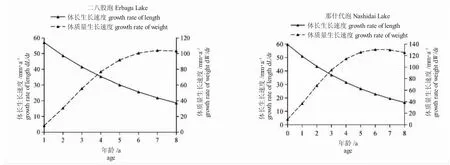
圖5 二八股泡、那什代泡河鱸體長和體質量的生長速度Fig.5 Growth rates of body length and body weight of perch Perca fluviatilis in Erbagu Lake and Nashidai Lake
2.2 死亡系數及臨界年齡
基于體長變換漁獲物曲線法(圖6),求得二八股泡和那什代泡河鱸總死亡系數Z,根據Pauly 經驗公式求得自然死亡系數M,進而求得捕撈死亡系數F 及開發率E(表2)。根據自然死亡系數及生長方程,求得二八股泡河鱸臨界年齡tc=4.71 a,對應的體長、體質量分別為241.84 mm 和284.13 g,那什代泡河鱸臨界年齡tc=4.77 a,對應的體長、體質量分別為254.08 mm 和357.68 g。
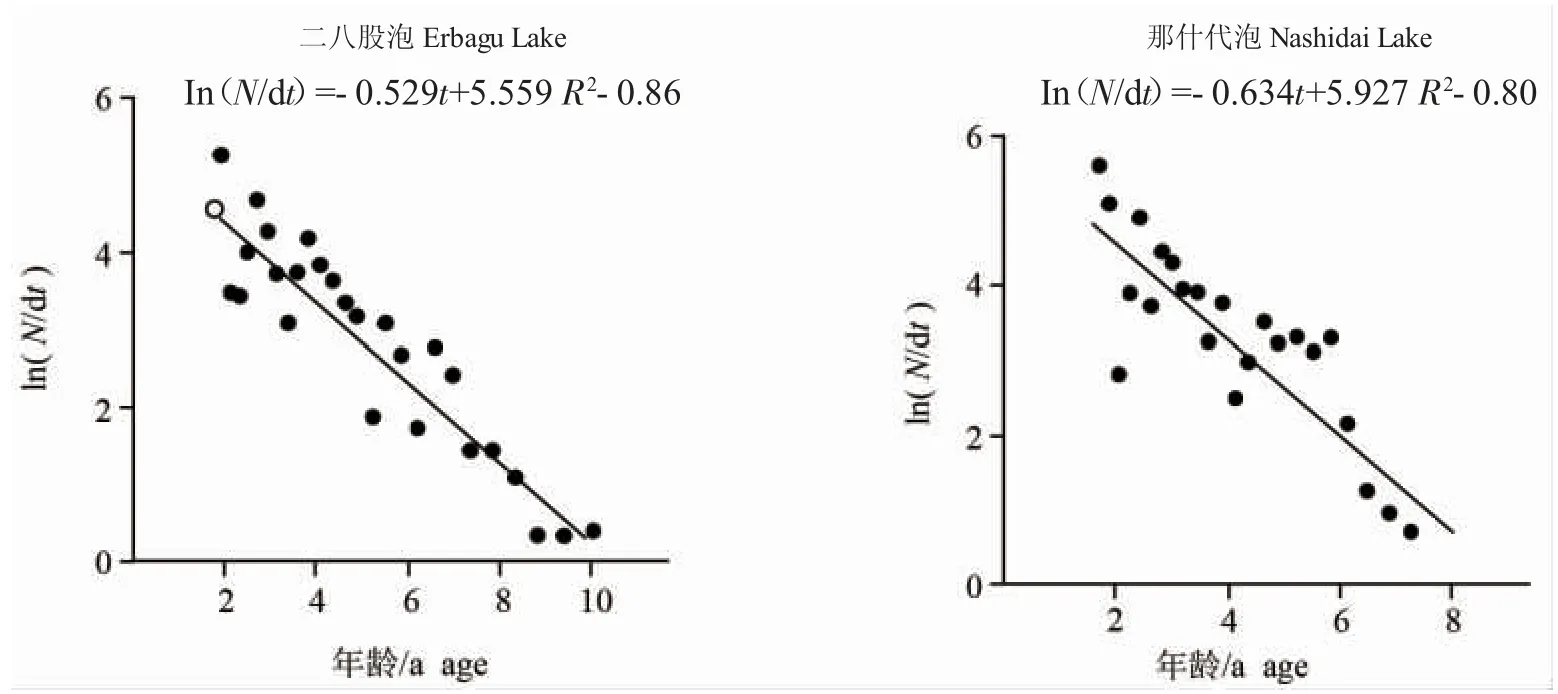
圖6 根據體長變換漁獲曲線估算總死亡系數Fig.6 Estimation of total mortality parameter from length converted catch curve
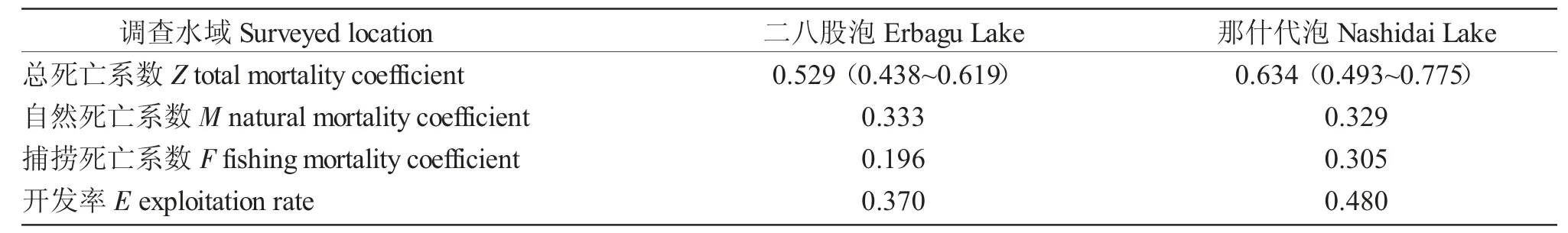
表2 二八股泡和那什代泡河鱸的死亡系數和開發率Tab.2 Mortality and exploitation rate for perch Perca fluviatilis in Erbagu Lake and Nashidai Lake
2.3 資源量利用及評估
2.3.1 資源量利用
本研究以70 mm 以下的河鱸作為群體補充量,根據Beverton-Holt 動態綜合模型估算Emax(最大漁獲量下的開發率)、E0.1(最大漁獲量下減少10%的開發率)、E0.5(生物量下降到原始水平的50%的開發率)值。結果表明,二八股泡河鱸開發率Emax、E0.1、E0.5數值依次為0.429、0.355 和0.265;那什代泡河鱸開發率Emax、E0.1、E0.5數值依次為0.424、0.352 和0.263。通過分析單位補充量漁獲量Y’/R、單位補充量生物量B’/R 與開發率E 的關系可得,隨著開發率E 的增大,單位補充量漁獲量Y’/R 呈現先增大后減少的趨勢,單位補充量生物量B’/R 呈現逐漸減少并趨于0 的趨勢(圖7)。二八股河鱸在現開發率保持不變的情況下,約35%的生物量資源得到保持,那什代泡在現開發率保持不變的情況下,約20%的生物量資源得到保持。根據殘存率計算公式,可得二八股泡河鱸種群數量捕撈死亡占15%、自然死亡占26%、殘存率為59%;那什代泡河鱸種群數量捕撈死亡占23%、自然死亡占24%、殘存率為53%。
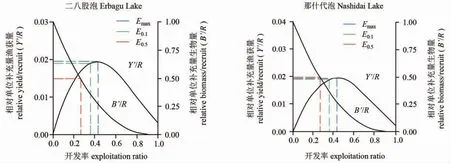
圖7 河鱸的Y'/R 和B'/R 二維分析Fig.7 Two-dimensional analysis for Y'/R and B'/R of perch Perca fluviatilis
2.3.2 資源量評估
依據大慶市連環湖漁業有限公司2021 年二八股泡、那什代泡河鱸捕撈產量數據,在最大體長河鱸的捕撈死亡系數不變的情況下,利用FISAT II 軟件推算出二八股泡河鱸的現有資源數量為265.59萬尾,資源量348.66 t(圖8);那什代泡河鱸的現有資源數量193.91 萬尾,資源量204.93 t(圖9)。二八股泡、那什代泡水域面積分別為22.67 km2、12 km2,河鱸種群密度分別為117154.83 尾/km2和161591.67尾/km2。

圖8 二八股泡河鱸資源量估算Fig.8 Estimation of population number of perch Perca fluviatilis in Erbagu Lake

圖9 那什代泡河鱸資源量估算Fig.9 Estimation of population number of perch Perca fluviatilis in Nashidai Lake
3 討論
3.1 生長特征
受內源和外源因子的影響,魚類的生長特征不同,通常利用生長系數k 和表觀生長指數(Φ)來判定魚類的生長速度[26]。受水域自然環境影響,同種魚類在不同水體環境條件下生長特征存在一定差異。蔣朝明[28]研究表明,嘉陵江不同江段的蛇(Saurogobio dabryi)生長特征明顯不同,上游水域蛇的生長系數及生長率明顯高于中、下游。比較不同水域河鱸的生長系數k(表3),根據Branstetter 對生長系數的分類,連環湖河鱸k 值為0.1~0.2 之間,屬于勻速生長型魚類[29]。連環湖二八股泡和那什代泡河鱸k 值均為0.16,與額爾齊斯河流域烏倫古湖和塘巴湖的河鱸無明顯差異,但與鴨綠江流域水豐水庫的河鱸差異較大。譚博真[30]研究表明,捕獲樣本的年齡結構差異是導致哲古錯高原裸鯉k 值異于其他高原魚類的原因之一,這可能也是水豐水庫河鱸生長系數與本研究結果不同的原因,故通過生長指數(Φ)來描述魚類生長性能更為系統[31]。連環湖二八股泡、那什代泡河鱸生長指數(Φ)相差極小,且與額爾齊斯河流域的烏倫古湖、塘巴湖和鴨綠江流域的水豐水庫也無明顯差異,表明生存環境的變化對河鱸的生長速度未產生明顯影響。異速生長指數(b)可反映魚類生長發育的不均勻性,b>3 為正異速生長魚類,且體質量生長速度快于體長[21]。連環湖二八股泡、那什代泡河鱸b 值均大于3,為典型的正異速生長魚類。比較不同水域河鱸的拐點年齡(ttp),連環湖略低于其他水域[33-35],這可能與采集樣本中高齡個體樣本數據較少及不同水域河鱸種群密度差異較大有關。
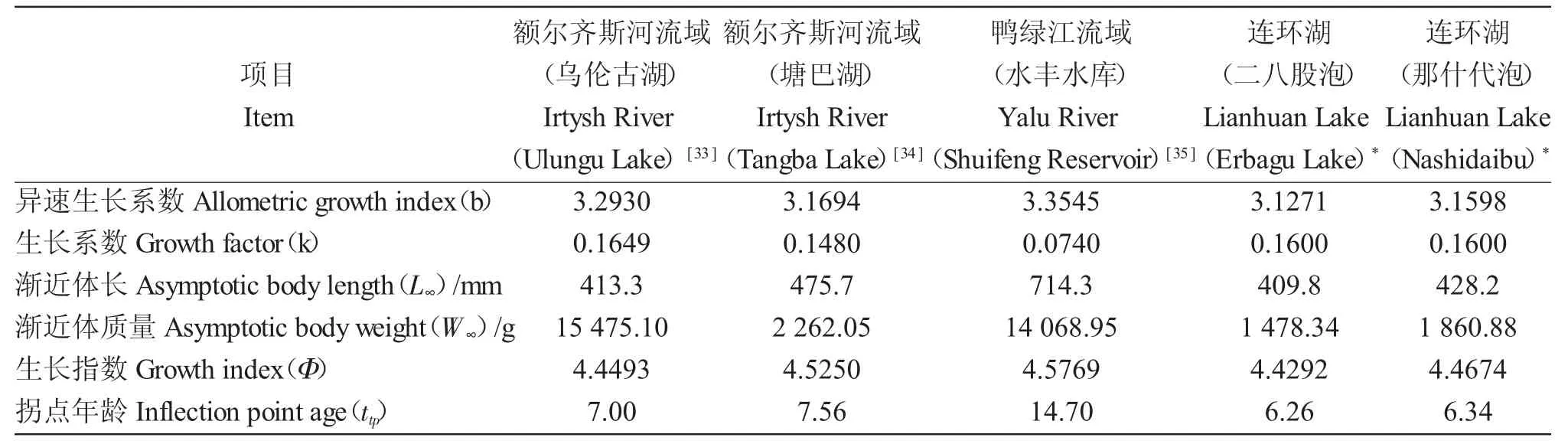
表3 不同流域河鱸的生長參數比較Tab.3 Comparison of growth parameters of perch Perca fluviatilis in different watersheds
3.2 資源利用現狀
水產養殖的隨意引進是外來物種入侵最主要的途徑之一,對入侵水域的魚類群落結構及土著魚類的生存空間造成一定影響[36]。河鱸為典型的肉食性魚類,幼魚階段主要以浮游動物為食,性成熟后以蝦類及小型魚類為主要食物來源[37],其食性特點與連環湖主要增養殖對象—大銀魚存在一定的相似性[38]。對連環湖河鱸消化道解剖發現,大銀魚是其食物組成之一。河鱸與大銀魚存在食物競爭及獵食關系,近幾年,連環湖漁業公司為了保障大銀魚的生長及產量,雖通過增加對河鱸的捕撈量來控制其種群數量,但效果并不理想。本研究結果顯示,以70 mm 作為河鱸的開捕體長,連環湖二八股泡、那什代泡河鱸的開發率分別為0.37、0.48。根據Gulland 提出的開發率評價標準,目前開發率在0~0.5范圍內,表明資源處于輕度開發狀態[39]。Mehanna[40]認為,只要當前開發率低于最大漁獲量下的開發率(Emax)則為安全狀態,二八股泡開發率低于Emax,那什代泡也僅略高于Emax,表明連環湖對河鱸資源的開發和利用不足,以目前的捕撈強度,連環湖河鱸可以長期保持較高的種群資源量,這主要由于河鱸在當地市場認可度不高,且未制定相應捕撈措施導致。種群殘存率分析顯示,目前兩個湖泊河鱸的種群殘存率均處于較高水平,捕撈死亡占比較少,不存在過度捕撈及種群資源崩潰的風險,故以目前的捕撈強度不足以明顯減少河鱸的種群資源量。
3.3 資源量評估
通常,河鱸2 齡性成熟[41],在二八股泡和那什代泡體長分別為150.67 mm 和156.98 mm,依據FISAT II 軟件,結合同一世代魚類種群的體長組成對現存資源量的評估結果顯示[42],二八股泡、那什代泡性成熟河鱸個體數量分別約為56.6 萬尾和38.2 萬尾,僅占種群總量的21.31%和19.70%,種群結構中低齡河鱸個體數量占比較大,如何減少低齡河鱸個體種群數量是遏制其種群資源量進一步增加的關鍵。結合不同體長組的捕撈死亡系數結果分析,在當前漁業形勢下,體長超過245.00 mm 的大型個體被捕撈的幾率明顯增加,這可能是在捕撈鰱、鳙等大型魚類的同時兼捕導致。假設資源現存量為可獲取的最大可持續產量(即1/2B0),根據MSY=0.5MB0,可得二八股泡、那什代泡河鱸年最大可持續產量分別為58.05 t 和33.71 t,目前兩處水域的年捕撈量均低于年最大可持續產量,此結果可為制定科學合理的河鱸資源管理策略提供參考。
3.4 建議
河鱸一般4—5 月自然繁殖[41],建議在連環湖冰層融化后立即開展河鱸的捕撈作業,減少繁殖群體數量,遏制河鱸的種群資源量進一步增加。目前連環湖主要作業方式為網箔,為了捕撈四大家魚,取魚部即后墻網的網目規格為6~8 cm,不利于對河鱸的捕撈作業,建議將后墻網的網目規格減少到3~5 cm,再對已誘入網箔的魚類進行挑選,減輕網目規格調整后對其他非目標魚類資源量的影響。地籠在捕撈小型魚類及保證魚類存活率方面優勢更為明顯[43],故建議于8—9 月在連環湖1 m、1.5 m、2 m 等不同深處放置網目規格為20 mm 地籠(1 km2水域不低于15 個)對河鱸幼魚進行集中捕撈。陳曉龍等[44]設計的大水面可調控捕撈裝置可滿足不同體長規格魚類的選擇性捕撈,已研究證實其在捕撈漁獲量及魚類存活率方面優于傳統作業方式,可滿足針對不同魚類的調控捕撈要求,但連環湖為冬季冰封性湖泊,該裝置捕撈平臺應改進為可移動式或以帶有平臺的運輸船只替代。如何更好地解決連環湖河鱸入侵問題,需進一步開展河鱸的生態學研究,探究其在生態系統中的地位及作用[45],突破北方寒地水域冬季冰封期魚類活動能力受限導致樣本采集困難問題,進一步補充冬季樣本數據,尋找控制河鱸入侵的有效方法。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
