東北碳酸鹽型鹽堿池塘耐(嗜)堿微生物的篩選及功能分析
2023-01-05 04:44:50張瑞羅亮王世會郭坤徐偉趙志剛
水產學雜志 2022年6期
張瑞,羅亮,王世會,郭坤,徐偉,趙志剛
(中國水產科學研究院黑龍江水產研究所,黑龍江省冷水性魚類種質資源及增養殖重點開放實驗室,黑龍江 哈爾濱 150070)
以松嫩平原鹽堿地為代表的東北地區鹽堿地是世界三大著名的碳酸鹽型鹽堿地之一。該地區的鹽堿水域以高pH(≥8.5)、高堿度(20~30 mmol/L HCO3-)、低鹽度(1‰~3‰ Na+)為特點,難以進行水產養殖。高pH 的水質不僅直接影響水產動物健康生長[1],還會導致水體中氨氮的毒性升高,增加對水產動物的危害[2,3]。因此,通過水質調控有效改善鹽堿水質環境,成為開發與利用鹽堿水域,推動鹽堿水漁業綠色發展的主要途徑之一。一直以來,微生物水質調控技術因其高效、綠色、安全的特點被廣泛應用于水產養殖[4-6],而“原位”功能性微生物的篩選和應用,是實現微生物水質調控的重要途徑[7]。
長期以來,國內外學者多以高鹽濃度耐受為關注點,針對海水、鹽湖、鹽堿地等不同鹽堿環境開展了大量耐(嗜)鹽堿菌的多樣性分析工作[8-14],分離鑒定了大量中度嗜鹽菌、輕度嗜鹽菌等具有重要研究價值的高鹽(堿)耐受菌株[8-14],并在2.5%(w/v)甚至更高的鹽(NaCl)濃度下分析菌株的耐堿能力[9,12]。值得注意的是,微生物要在強堿性條件下生長,就必需調節并維持細胞內pH 范圍以滿足其生命活動的需要。而通過細胞膜上的Na+/H+、K+/H+等逆向轉運蛋白,在Na+等的跨膜運輸的同時調節細胞內的pH,是耐(嗜)堿菌最主要的抗逆途徑之一[15]。因此環境中鹽濃度在很大程度上影響著菌株的耐堿能力。
迄今,尚未有以低鹽度、高pH 為篩選條件,針對碳酸鹽型鹽堿水環境特點進行的耐(嗜)堿微生物篩選及多樣性分析的報道,也缺少菌株在低鹽濃度下耐堿能力的分析。本研究對大慶連環湖——典型碳酸鹽型鹽堿地區池塘水體和底泥中的耐(嗜)堿微生物進行篩選分離、鑒定分類,分析其低鹽度下耐堿能力及pH 調節功能,為以“原位”功能微生物為核心的原位水質調控技術提供微生物資源保障,為碳酸鹽型鹽堿漁業的綠色發展提供支撐。
1 材料和方法
1.1 培養基及儀器試劑
富集培養基(3% Sehgal-Gibbons):胰蛋白胨5 g,酵母粉10 g,酸水解酪蛋白5 g,檸檬酸三鈉3 g,KCl2 g,MgSO4·7H2O20 g,NaCl30 g,加水定容至1 000 mL,用NaOH 調pH 至8.0,滅菌備用。
篩選培養基(LB):胰蛋白胨10 g,酵母粉5 g,NaCl 10 g,瓊脂15 g,加水定容至1 000 mL,用NaOH調pH 至8.5,滅菌備用。
耐堿上限測試培養基:液體LB 培養基,用NaOH 調至不同pH(7.0、7.5、8.0、8.5、9.0、9.5、10、11、12),滅菌備用。
pH 調節能力測試培養基:葡萄糖20 g,酵母粉5 g,蛋白胨5 g,NaCl 50 g,配置固體培養基時額外加入瓊脂15 g,加水定容至1 000 mL,用NaOH 調pH 至7.0,滅菌備用。
PCR 儀購自Bio-Rad 公司;凝膠成像分析儀購自Bio-Rad 公司;全溫振蕩培養箱購自滄州萊玻特瑞儀器設備有限公司(ZQPL-200);生化培養箱購自上海博迅實業有限公司(SPX-100B-Z);紫外分光光度計購自上海佑科儀器儀表有限公司(752N);便攜式pH 計購自上海沛瑞儀器有限公司(PH400);通用引物由北京華大基因研究院合成;PCR 預混合溶液購自上海圣生物科技有限公司。
1.2 樣品采集
2020 年6 月,從大慶連環湖地區的9 口養殖池塘中隨機采集水樣(嫩江水水源)與底泥樣(距離水底0~10 cm)共12 份,分別裝入無菌三角瓶中,帶回實驗室立即進行菌株富集與分離。
1.3 菌株富集與分離
將各采集樣品混合均勻,稱取3 g(3 mL)置于200 mL 富集培養基中,30℃150 r·min-1振蕩培養2~3 d 后,將富集的菌懸液在無菌水中逐級稀釋至10-4、10-5、10-6和10-7倍,然后取各稀釋度的菌懸液0.1 mL 分別涂布于pH 8.5 的篩選培養基平板上,在30℃靜置培養。3~5 d 后,挑取單菌落進行多次劃線純化。最后根據菌落形態、顏色以及顯微形態等表型特征鑒定為純培養后,將純培養菌液保藏于-80℃。
1.4 基于16S rRNA 基因序列的系統發育分析
采用菌液PCR 法擴增純培養細菌的16S rRNA基因序列。擴增引物采用擴增細菌16S rRNA 的通用引物Eubac27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和Eubac1492R(5’-GGTTACCTTGTTACGACT T-3’)。PCR 以過夜培養菌液為模板,反應體系參照2×Hieff PCR Master Mix 使用說明書配置。反應程序:95℃10 min;94℃l min,55℃30 s,72℃1.5 min,30 個循環;72℃10 min。PCR 產物經電泳檢測后送北京華大基因研究院測序,測序引物與擴增引物相同。
測序所得序列提交GenBank 數據庫,獲取序列號(Accession number)。采用Nucleotide Blast 在線分析,找出各序列在NCBI 數據庫中同源性最高的已鑒定菌株序列,用MEGA 6.0 中的Clustal X 程序進行多序列比對,然后用Neighbor-Joining 方法選擇Bootstrap 為1 000 個重復構建系統發育學進化樹。
1.5 菌株耐堿上限測試
所有待試菌株經LB(pH 7.0)培養基過夜活化后,分別按1%接種量接種于不同pH 培養基中,在30℃150 r·min-1震蕩培養24 h 后觀察生長情況。
1.6 菌株pH 調節能力驗證
所有待試菌株經活化后,分別按1%接種量接種于200 mL 培養基中驗證pH 調節能力,培養液于30℃150 r·min-1震蕩培養,每隔4 h 取20 mL 培養菌液測定pH 與OD600吸光度,直至24 h 結束。
2 結果與分析
2.1 菌株的篩選與分離
從本次采集的12 份樣品中共篩選出純培養菌株49 株,經16S rRNA 基因序列鑒定,最終共確定菌株24 株。它們分屬于氣單胞菌屬(Aeromonas)、芽孢桿菌屬(Bacillus)、腸桿菌屬(Enterobacter)、腸球菌屬(Enterococcus)、微小桿菌屬(Exiguobacterium)、鹽單胞菌屬(Halomonas)、賴氨酸芽孢桿菌屬(Lysinibacillus)、動性球菌屬(Planococcus)以及假單胞菌屬(Pseudomonas)共9 個菌屬。其中芽孢桿菌屬與微小桿菌屬為優勢菌屬,分別為7 株與6 株,占總菌株量的29.17%和25.00%(圖1)。其次是動性球菌屬(Planococcus)3 株,占總菌株量的12.50%;腸球菌屬與腸桿菌屬各2 株,分別占8.33%;氣單胞菌屬、鹽單胞菌屬、賴氨酸芽孢桿菌屬以及假單胞菌屬各1 株,分別占菌株總量的4.17%。
2.2 基于16S rRNA 基因序列的系統發育學分析
用DNAMAN6.0 軟件分別比對分析上述24 株菌的16S rRNA 基因序列及其已鑒定的最近緣種菌株的同源性。結果表明:CT-WN-B17 與已鑒定的最近緣種菌株同源性為96.35%,CT-MH6-7 與已鑒定的最近緣種菌株同源性為96.84%,而其余同源性均在99.58%~100%之間。碳酸鹽型池塘耐(嗜)堿菌菌群中所有菌株與已鑒定近緣菌株在系統發育進化樹上也穩定聚簇(圖2)。上述結果支持將除CTWN-B17、CT-MH6-7 以外的22 株菌株認定為各自相應的已鑒定最近緣種,而菌株CT-WN-B17 和CT-MH6-7 的16S rRNA 序列與已鑒定最近緣種同源性較低,系統進化關系較遠,具有新種潛質,需進行進一步鑒定,因而暫將它們分別命名為Lysinibacillus sp.與Enterococcus sp.(表1)。
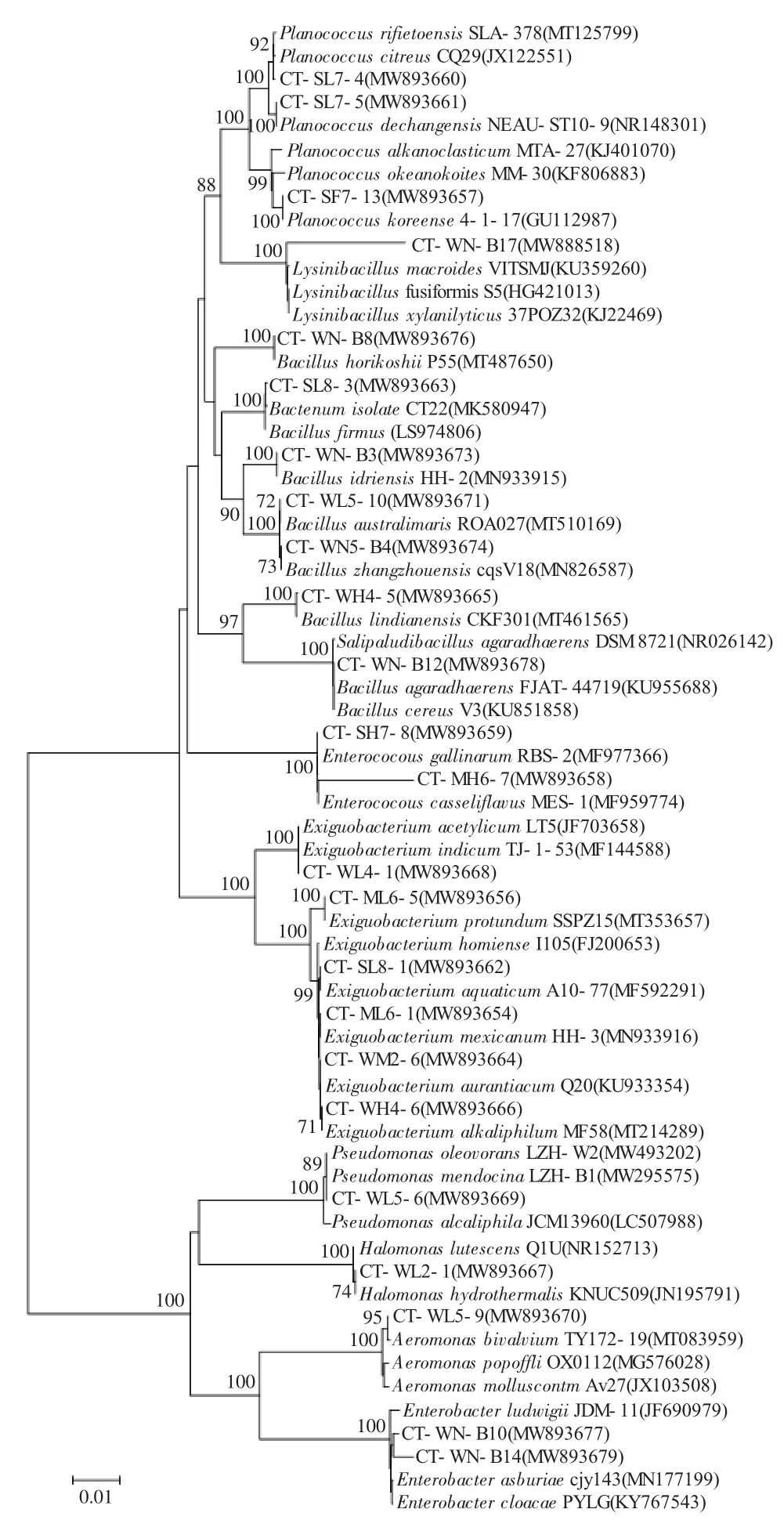
圖2 東北地區碳酸鹽型鹽堿池塘中耐(嗜)堿細菌群落的系統發育樹Fig.2 Neighbor -Joining tree of alkalitolerant(alkaliphilic)bacteria in carbonate saline-alkali ponds in northeast China
2.3 耐堿上限測定
以上24 株菌株分屬于細菌域的2 個門——變形菌門(Proteobacteria)和厚壁菌門(Firmicutes);9個屬——氣單胞菌屬、芽孢桿菌屬、腸桿菌屬、腸球菌屬、微小桿菌屬、鹽單胞菌屬、賴氨酸芽孢桿菌屬、動性球菌屬以及假單胞菌屬。其中厚壁菌門為優勢類群,菌株占比達79.17%;芽孢桿菌屬與微小桿菌屬為優勢菌屬,菌株占比分別為29.17%和25.00%。
以上24 株菌株在1%NaCl 濃度下的耐堿上限在pH 9.0~10.0 之間,其中7 株能耐受pH9.0,芽孢桿菌屬為最優勢菌屬(4/7);10 株能耐受pH9.5,其中芽孢桿菌屬為最優勢菌屬(3/10);7 株能耐受pH10.0,其中微小桿菌屬為最優勢菌屬(4/7)(表1)。上述所有菌株并未表現出對堿性pH 的生長依賴性,均屬于耐堿菌。
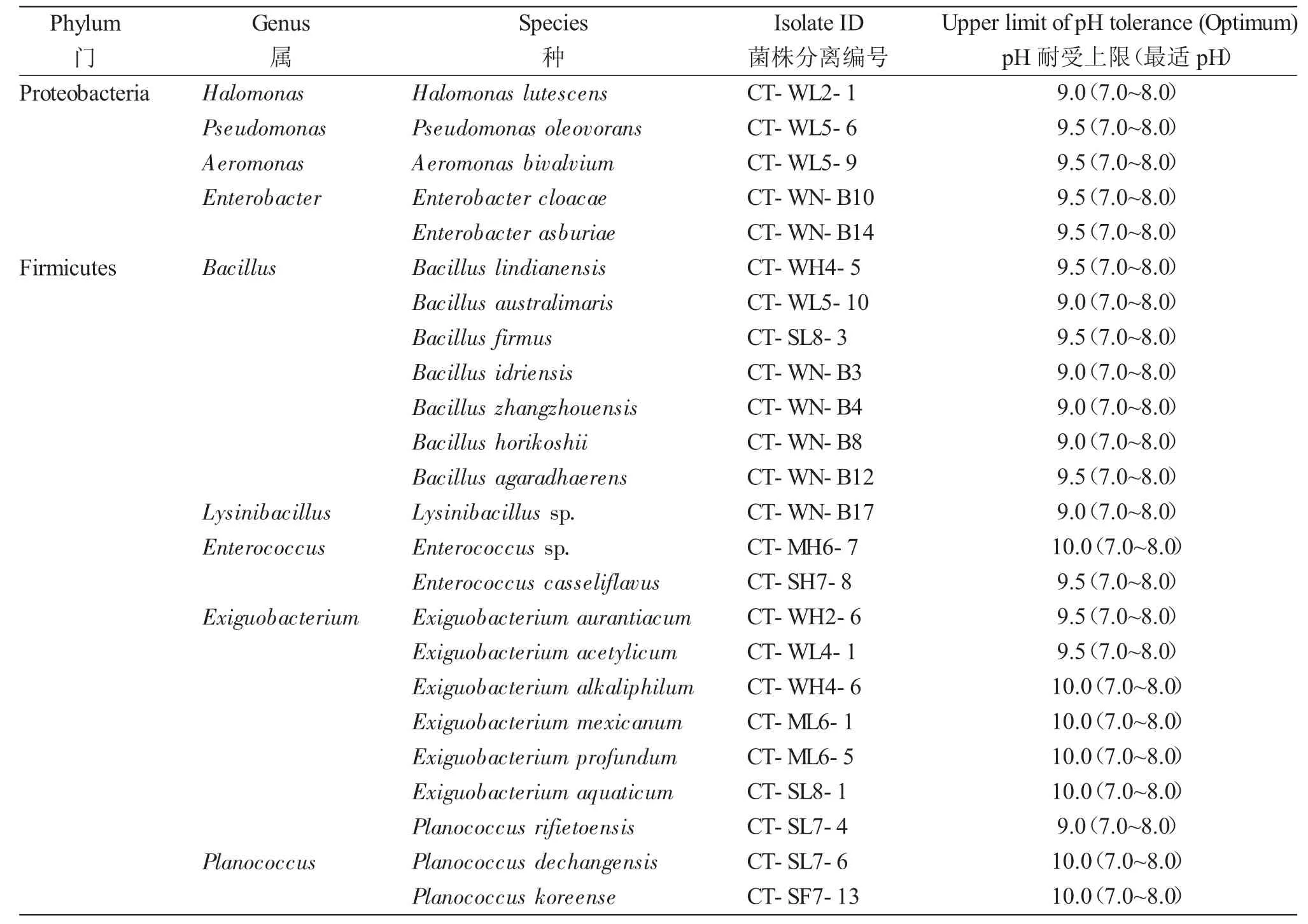
表1 東北地區碳酸鹽型鹽堿池塘中的耐(嗜)堿細菌及其耐堿能力分析Tab.1 Alkalitolerant(alkaliphilic)bacteria and their alkaline-tolerance capacity in carbonate saline-alkali ponds in northeast China
2.4 pH 調節能力測定
碳酸鹽型鹽堿池塘耐堿細菌群落中有兩大優勢菌屬,分別為芽孢桿菌屬和微小桿菌屬。后者的功能之一就是可以利用葡萄糖等碳源產酸[16],導致培養環境的pH 下調。為探究鹽堿池塘中優勢菌群菌株,即芽孢桿菌屬與微小桿菌屬菌株的pH 調節能力,以葡萄糖為補充碳源培養菌株,測定隨菌株繁殖的培養基pH 的變化。
隨24 h 內微小桿菌屬菌株的不斷生長繁殖,菌株CT-WL4-1 將培養基pH 下調了2.5 個單位(圖3-A);盡管菌株CT-WH4-6 最終的繁殖水平與CT-WL4-1 相差無幾,但僅將培養基pH 下調了0.6個單位(圖3-B);菌株CT-SL8-1、CT-WH2-6、CT-ML6-1 和CT-ML6-5 繁殖不十分旺盛(OD600≤1.0),24 h 內培養基pH 下調了1.3~1.5 個單位(圖3-C~F)。
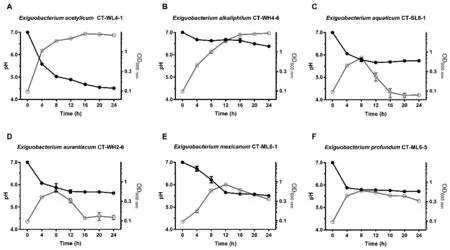
圖3 東北地區碳酸鹽型鹽堿池塘微小桿菌屬細菌的pH 調節能力Fig.3 pH regulation of Exiguobacterium strains in carbonate saline-alkali ponds in northeast China
芽孢桿菌屬菌株也具有不同程度調節pH 的能力。如圖4 所示,菌株CT-WN-B4、CT-WL5-10 在24 h 內繁殖旺盛,分別使培養基pH 下調了1.1 和1.3 個單位(圖4-A、B);菌株CT-WH4-5、CT-WN-B12 在實驗條件下生長較為微弱,均僅將培養基pH 下調了不足1 個單位(圖4-C、D);菌株CT-SL8-3、CT-WN-B3 和CT-WN-B8 的繁殖并不十分旺盛(OD600≤1.1),在24 h 內將培養基pH 分別下調了1.8、1.9 和1.2 個單位(圖4-E~G)。上述結果顯示,在本實驗條件下,已篩選鑒定的6 株微小桿菌屬菌株與7 株芽孢桿菌屬菌株中,微小桿菌屬菌株CT-WL4-1 具有最強的pH 調節能力,芽孢桿菌屬菌株CT-SL8-3 和CT-WN-B3 次之。
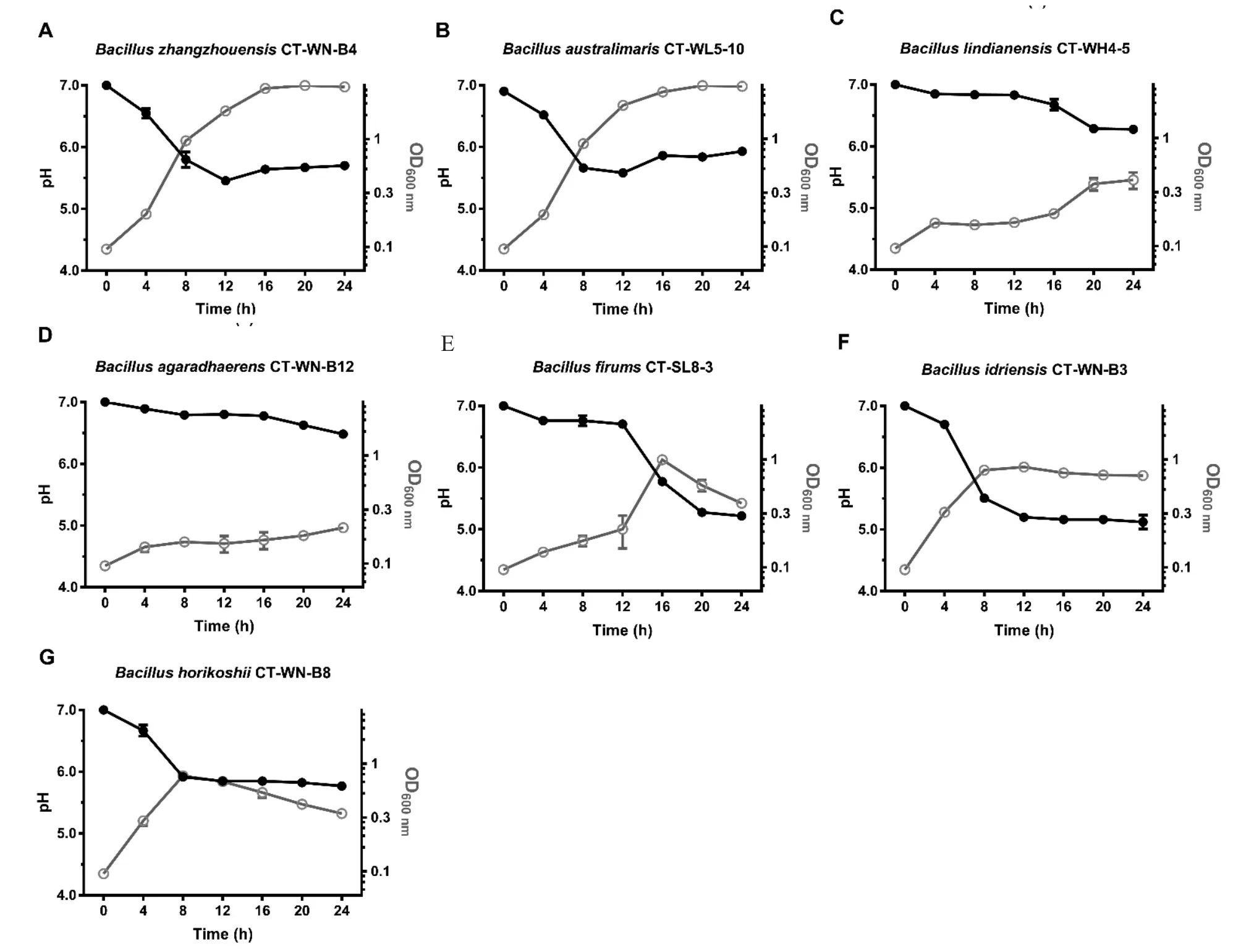
圖4 東北地區碳酸鹽型鹽堿池塘中芽孢桿菌屬細菌的pH 調節能力Fig.4 pH regulation of Bacillus strains in carbonate saline-alkali ponds in northeast China
3 討論
本研究以高pH、低鹽度為條件從典型碳酸鹽型鹽堿池塘水體和底泥樣品中定向富集、篩選耐(嗜)堿菌,通過16S rRNA 基因同源性比對和系統發育學分析進行菌株鑒定分類,獲取耐(嗜)堿菌多樣性信息。結果表明,東北地區碳酸鹽型鹽堿池塘中耐堿菌種群豐富,共分離到的49 個菌株,分屬于細菌域中2 個門(Firmicutes、Proteobacteria)、9 個屬、24 個種。其中多數菌株屬于厚壁菌門(Firmicutes),兩個優勢屬分別為芽孢桿菌屬(7 株,占總菌株的29.17%)和微小桿菌屬(6 株,占總菌株的25%)。絕大多數菌株的16S rRNA 基因序列與已鑒定的最近緣種序列同源性在99.58%~100%之間。僅腸球菌屬菌株CT-MH6-7 與賴氨酸芽孢桿菌屬菌株CT-WN-B17 的16S rRNA 序列與已鑒定的最近緣種菌株同源性較低,分別為96.35%、96.84%,且系統進化關系較遠,認為其具有新種潛質。
多年來,科研工作者圍繞多個鹽堿環境開展了耐(嗜)鹽菌的多樣性研究,已在碳酸鹽型鹽堿土壤中篩選鑒定出34 株耐(嗜)鹽菌。這些菌株大多具有5%~10%的耐鹽能力和不同程度的耐堿能力[9]。與之相比,本研究獲得的碳酸鹽型鹽堿池塘耐堿菌多樣性并不十分豐富。比較二者菌群結構,發現其均包含芽孢桿菌屬、微小桿菌屬、鹽單胞菌屬、腸桿菌屬、假單胞菌屬和動性球菌屬(Planococcus),這表明上述6 個菌屬的菌株在碳酸鹽型鹽堿土壤中和水體中均有分布。不同的是,土壤中的最優勢菌屬葡萄球菌屬(Staphylococcus)并不包含在池塘樣品中,而鹽堿池塘中的優勢菌屬微小桿菌屬(Exiguobacterium),在土壤僅分離到1 株。這似乎并不是由菌株篩選條件所致。根據現有研究背景,微小桿菌屬菌株可以耐受0%~10%濃度的NaCl[16],即采用耐(嗜)鹽菌的篩選條件也可將其分離出,相似的耐(嗜)堿菌的篩選條件也可將葡萄球菌屬菌株分離出來。因此,可以認為兩群落結構的差異是由不同菌屬微生物對生存環境的偏好性選擇導致。微小桿菌屬曾因并未在其他鹽堿環境中被篩選得到而被認為是屬于東北地區松嫩平原鹽堿土壤的獨特耐鹽菌群類[9],因此建議,微小桿菌屬可作為東北地區碳酸鹽型鹽堿水體環境的代表性耐鹽堿菌屬。
一直以來對于耐堿菌與嗜堿菌的定義并不明確和統一。本研究根據眾多科研工作者的分類習慣,將能在高pH 條件下生長,但最適值并不在堿性pH 范圍內的歸類為耐堿菌;而最適生長pH 在9.0以上,pH 耐受高達10~12 的則稱之為嗜堿微生物[17]。本研究獲得的所有菌株并未表現出堿性條件的生長依賴性,pH 耐受上限介于9.0~10.0 之間,屬于耐堿菌。本結果顯然低于一些現有的研究背景[9,18-20],這是因為本實驗條件中更低的鹽濃度影響了菌株的耐堿能力上限。但本研究的結果更接近菌源環境的實際條件,更有菌株的原位應用指導意義。
菌群中優勢的、且更強耐堿能力的微小桿菌屬菌株,其另一重要功能就是可以利用葡萄糖等碳源產酸[16]。本研究以葡萄糖為補充碳源,以pH 調節能力為目標,在兩大優勢菌屬中篩選出3 株具有較強pH 調節能力的耐堿菌(包括1 株微小桿菌屬菌株,2 株芽孢桿菌屬菌株),能夠在24 h 內將培養基pH下調1.8~2.5 個單位,具有微生物水質調控應用潛力。然而這些菌株是否通過利用葡萄糖合成有機酸來調節pH;有些菌株是否也可以利用其他碳源產酸來調節pH,這些問題均需進一步驗證與探究。
本研究填補了我國東北地區碳酸鹽型鹽堿水土環境——鹽堿池塘的耐堿菌多樣性的研究空白,在優勢菌屬中篩選出了具有較強pH 調節能力的耐堿菌株,為碳酸鹽型鹽堿養殖水質調控技術的研發提供支撐。
