套袋對(duì)富士芽變優(yōu)系果皮色澤及花青苷合成相關(guān)酶活性的影響
2023-01-12 09:14:52蘇婧怡李文勝魯芯志張振軍陳高安
經(jīng)濟(jì)林研究 2022年4期
蘇婧怡,李文勝,魯芯志,張振軍,陳高安
(1. 新疆農(nóng)業(yè)大學(xué) a. 園藝學(xué)院;b. 林學(xué)與風(fēng)景園林學(xué)院,新疆 烏魯木齊 830000;2. 新疆阿克蘇地區(qū)林業(yè)技術(shù)推廣服務(wù)中心,新疆 阿克蘇 842008;)
蘋(píng)果是薔薇科Rosaceae仁果亞科Pomideae蘋(píng)果屬M(fèi)alus.Mill.的落葉喬木果樹(shù)[1]。新疆阿克蘇地區(qū)栽培的蘋(píng)果品種以富士為主,該地區(qū)憑借獨(dú)特的地理氣候條件生產(chǎn)出了具有鮮明區(qū)域特色的“冰糖心”富士蘋(píng)果,果實(shí)汁水豐富,口感脆甜,深受消費(fèi)者的喜愛(ài)。芽變是植物產(chǎn)生新變異的無(wú)限豐富的源泉,既可為雜交育種提供新的種質(zhì)資源,又可從中直接選育出優(yōu)良的新品種;而芽變育種是選育新品種的一種簡(jiǎn)易而有效的方法。宋來(lái)慶等[2]研究發(fā)現(xiàn),在長(zhǎng)期的栽培過(guò)程中,長(zhǎng)富2號(hào)是最容易發(fā)生變異且變異類型最豐富的富士品種。陳學(xué)森等[3]認(rèn)為,富士蘋(píng)果因其質(zhì)優(yōu)、晚熟、耐貯等特點(diǎn)在我國(guó)蘋(píng)果產(chǎn)業(yè)中獨(dú)占鰲頭,推動(dòng)了我國(guó)蘋(píng)果產(chǎn)業(yè)的高速發(fā)展。但是,生產(chǎn)栽培中富士蘋(píng)果時(shí)常表現(xiàn)出著色不佳等問(wèn)題,生產(chǎn)者采用套袋措施解決蘋(píng)果著色不良的問(wèn)題,然而采用這種措施存在生產(chǎn)成本高和果實(shí)風(fēng)味欠佳等問(wèn)題,因而,選育出著色好且風(fēng)味佳的免套袋富士品種是蘋(píng)果育種者們追求的目標(biāo),也是果農(nóng)的迫切期望。為此,本課題組成員針對(duì)阿克蘇富士芽變發(fā)生頻繁的特點(diǎn),開(kāi)展了優(yōu)異富士芽變選種的工作。2016年,本課題組成員在阿克蘇地區(qū)紅旗坡農(nóng)場(chǎng)果園普查時(shí)發(fā)現(xiàn)了長(zhǎng)富2號(hào)的紅色突變枝,果實(shí)由原來(lái)的條紋紅色突變?yōu)闈饧t色,果面呈濃紅色,著色艷麗,特別是在不套袋時(shí)其果實(shí)也表現(xiàn)出著色優(yōu)異的特性,為免袋栽培提供了可能性,也為后續(xù)果實(shí)著色機(jī)理的深入研究提供了試材。
花青苷是蘋(píng)果果皮中存在的一種重要的類黃酮色素,與葉綠素、類胡蘿卜素共同決定果皮的呈色[4]。而富士蘋(píng)果的紅色主要是由花青苷產(chǎn)生的,果實(shí)的著色程度取決于花青苷的含量及其在色素中所占的比例[5]。有關(guān)研究結(jié)果表明,花青苷的合成與苯丙氨酸裂解酶等酶活性密切相關(guān)[6],但是,花青苷的合成與相關(guān)酶活性的關(guān)系十分復(fù)雜。因此,探討富士蘋(píng)果中花青苷的合成與相關(guān)酶活性的關(guān)系,對(duì)于蘋(píng)果果實(shí)著色機(jī)理的研究具有重要意義。前人關(guān)于蘋(píng)果果皮著色的研究?jī)?nèi)容多集中在果實(shí)整個(gè)發(fā)育期果皮色素變化規(guī)律的比較分析和果實(shí)發(fā)育期果皮花青苷合成相關(guān)酶活性的變化等方面。王龍等[7]研究了‘紅香酥梨’果實(shí)發(fā)育過(guò)程中果皮色素的變化規(guī)律;劉曉靜等[8]研究了國(guó)光蘋(píng)果及其紅色芽變果實(shí)發(fā)育期果皮花青苷合成相關(guān)酶活性的變化情況。而有關(guān)套袋、解袋后果皮著色的研究報(bào)道較少,僅查詢到夏靜等[9]、遲馨等[10]關(guān)于解袋后富士蘋(píng)果著色的相關(guān)因子變化情況的研究報(bào)道。為此,本研究以長(zhǎng)富2號(hào)芽變優(yōu)系為試材,測(cè)定并分析了套袋與未套袋果實(shí)果皮的色澤參數(shù)值、葉綠素含量、花青苷含量及其相關(guān)酶活性,探討了富士紅色芽變優(yōu)系著色的生理機(jī)理。
1 材料與方法
1.1 材 料
試驗(yàn)地位于新疆維吾爾自治區(qū)阿克蘇地區(qū)紅旗坡農(nóng)場(chǎng)園林二分場(chǎng)蘋(píng)果園,供試材料為該果園中發(fā)現(xiàn)的富士芽變優(yōu)系,芽變優(yōu)系為長(zhǎng)富2號(hào)母樹(shù)芽變產(chǎn)生,樹(shù)體健壯,生長(zhǎng)勢(shì)一致,其樹(shù)齡均為15 a。該果園的土壤為砂壤土,蘋(píng)果樹(shù)的株行距為5 m×4 m,其栽培管理水平一致。
1.2 套袋處理
2021年6月12日套袋,采收前的第30天即9月20日解袋。芽變優(yōu)系和長(zhǎng)富2號(hào)的套袋處理分別標(biāo)記為T(mén)1與T2,芽變優(yōu)系和長(zhǎng)富2號(hào)的未套袋處理分別標(biāo)記為C1與C2。分別于解袋日后0(即解袋日當(dāng)天)、2、4、6、8、10、15、20、30 d采摘套袋和未套袋的果實(shí)樣品。采樣時(shí)分別從蘋(píng)果樹(shù)的東、西、南、北4個(gè)方位各采摘3~5個(gè)蘋(píng)果果實(shí)樣品,采后立即用裝有冰袋的泡沫箱帶回實(shí)驗(yàn)室,用削皮器刮下果皮,將混合的果皮樣品放入液氮中冷凍,做好標(biāo)記后置于液氮罐中保存待測(cè)。
1.3 果皮色澤參數(shù)的測(cè)定
將果皮表面擦凈,在果皮赤道部位選擇5個(gè)點(diǎn),用NH310全功能便攜式色差儀,按照國(guó)際照明委員會(huì)頒布的CIELAB評(píng)價(jià)體系,測(cè)定果皮L*、a*、b*、C*、h°值。L*值表示亮度,其絕對(duì)值越大,表示亮度越高;a*值表示紅綠色度,若a*值為正值,則代表紅色,若a*值為負(fù)值,則代表綠色,其絕對(duì)值越大,表示顏色越深;b*值表示黃藍(lán)色度,若b*值為正值,則代表黃色,若b*值為負(fù)值,則代表藍(lán)色,絕對(duì)值越大,表示顏色越深;C*值表示色澤飽和度;h°為色調(diào)角,若h°為0°,則表示紅色—紫色,若h°為90°,則表示黃色,若h°為180°,則表示藍(lán)綠色,若h°為270°,則表示藍(lán)色。
1.4 果皮中花青苷和葉綠素含量的測(cè)定
參照Pirie等[11]與王惠聰?shù)萚12]所用方法測(cè)定花青苷含量。稱取0.5 g果皮,用液氮研磨成粉末,轉(zhuǎn)移至10 mL的離心管中,加入5 mL的1%HCL-甲醇溶液,于4 ℃避光條件下浸提24 h,期間晃動(dòng)2次,以果皮碎渣變白為準(zhǔn)。然后于4 ℃的溫度條件下以12 000 r/min的轉(zhuǎn)速離心10 min,以1%的HCL-甲醇作對(duì)照,利用分光光度計(jì),在波長(zhǎng)分別為530、657 nm處測(cè)定吸光度。參照仝月澳等[13]所用方法(略有改動(dòng))測(cè)定葉綠素含量。稱取果皮0.5 g,用80%預(yù)冷的丙酮研磨,轉(zhuǎn)移至10 mL的離心管中,用80%的丙酮反復(fù)沖洗研缽,定容至10 mL。于4 ℃避光條件下浸提24 h,期間多次搖勻。于4 ℃的溫度條件下以12 000 r/min的轉(zhuǎn)速離心20 min,以80%的丙酮作為空白對(duì)照,利用分光光度計(jì)在波長(zhǎng)分別為470、645、663 nm處測(cè)定吸光度。
1.5 果皮中花青苷合成相關(guān)酶活性的測(cè)定
參照王惠聰?shù)萚12]所用方法分別提取苯丙氨酸解氨酶(PAL)與查爾酮異構(gòu)酶(CHI)的酶液,參照Lister等[14]所用方法測(cè)定PAL與CHI的活性;參照Murray等[15]所用方法提取二氫類黃酮還原酶(DFR)、類黃酮3,5-糖苷轉(zhuǎn)移酶(UFGT)的酶液,參照Stafford等[16]報(bào)道的方法測(cè)定DFR的活性,參照Lister等[17]報(bào)道的方法測(cè)定UFGT的活性。
1.6 數(shù)據(jù)分析
采用Microsoft Excel軟件進(jìn)行數(shù)據(jù)處理和計(jì)算,采用IBM SPSS statistics 23軟件進(jìn)行顯著性分析,采用Origin與Excel軟件繪圖。
2 結(jié)果與分析
2.1 果皮色澤參數(shù)的變化趨勢(shì)
套袋和未套袋的果皮亮度L*值的變化趨勢(shì)如圖1所示。從圖1中可以看出,以解袋日9月20日當(dāng)天(即解袋后0 d)為觀測(cè)起始日,隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋的果皮L*值整體呈下降趨勢(shì)。套袋的芽變優(yōu)系果皮的L*值在解袋后0~8 d急速下降,10~30 d緩慢下降;套袋的長(zhǎng)富2號(hào)果皮的L*值在解袋后0~2 d呈下降趨勢(shì),解袋后3 d略有回升而后平緩下降;套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的L*值在解袋后0~4 d沒(méi)有顯著差異,但長(zhǎng)富2號(hào)果皮的L*值整體高于芽變優(yōu)系的。未套袋的芽變優(yōu)系果皮的L*值在解袋后10 d降至最低,解袋后15 d快速上升,之后又呈下降趨勢(shì);未套袋的長(zhǎng)富2號(hào)果皮的L*值在解袋后0~4 d呈下降趨勢(shì),解袋后6 d短暫回升,之后又緩慢下降;未套袋的長(zhǎng)富2號(hào)果皮的L*值整體高于未套袋的芽變優(yōu)系的L*值。解袋后0~4 d,套袋果皮的L*值極顯著高于未套袋的(P<0.01),但套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)的果皮之間的L*值無(wú)顯著差異;解袋后30 d,套袋和未套袋的芽變優(yōu)系果皮之間、套袋和未套袋的長(zhǎng)富2號(hào)果皮之間其L*值均不存在顯著差異。
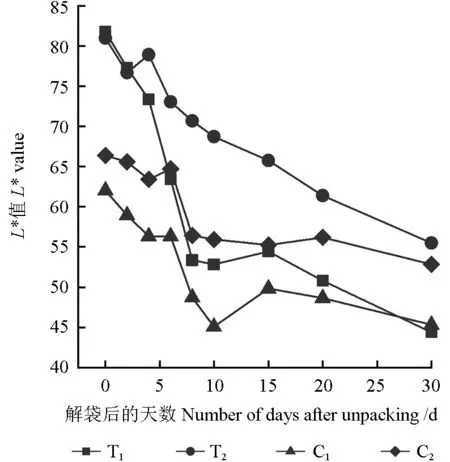
圖1 不同處理的果皮亮度值L*的變化趨勢(shì)Fig. 1 The change trend of peel brightness value L* under different treatments
套袋和未套袋的果皮紅綠色度a*值與黃藍(lán)色度b*值的變化趨勢(shì)如圖2所示。由圖2可知,隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋果皮的a*值總體均呈上升趨勢(shì)。套袋的芽變優(yōu)系果皮的a*值,解袋后0~8 d快速上升,8~10 d無(wú)明顯波動(dòng),10~15 d快速上升,15~30 d緩慢上升;套袋的長(zhǎng)富2號(hào)果皮的a*值,解袋后0~4 d呈上升趨勢(shì),解袋后6 d短暫下降之后又上升,解袋后8~15 d沒(méi)有明顯的變化趨勢(shì),解袋后20~30 d大幅上升;套袋的芽變優(yōu)系果皮的a*值上升最快且整體高于套袋的長(zhǎng)富2號(hào)果皮的a*值。未套袋的芽變優(yōu)系果皮的a*值,解袋后0~4 d呈上升趨勢(shì),解袋后6 d短暫下降,之后又上升;而未套袋的長(zhǎng)富2號(hào)果皮的a*值,解袋后0~8 d呈“W”曲線型的變化趨勢(shì),解袋后10 d開(kāi)始平穩(wěn)上升;未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的a*值,均呈現(xiàn)出平緩上升后短暫而快速下降然后又上升的變化趨勢(shì),解袋后30 d均達(dá)到最高值。解袋后30 d,套袋和未套袋的芽變優(yōu)系果皮之間、套袋與未套袋的長(zhǎng)富2號(hào)果皮之間果皮a*值的差異均極顯著(P<0.01),套袋和未套袋的果皮a*值由低至高依次為C2<C1<T2<T1。
圖2表明,隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋果皮的黃藍(lán)色度b*值均呈下降趨勢(shì)。套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的b*值均呈“臺(tái)階”式下降趨勢(shì);未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)的b*值,解袋后0~4 d均快速下降,其中芽變優(yōu)系的b*值分別在解袋后6和10 d回升,而長(zhǎng)富2號(hào)的b*值僅在解袋后6 d回升,此后緩慢下降。解袋后30 d,套袋和未套袋的芽變優(yōu)系果皮之間、套袋和未套袋的長(zhǎng)富2號(hào)果皮之間b*值的差異均極顯著(P<0.01),套袋和未套袋果皮的b*值由低至高依次為T(mén)1<T2<C1<C2。
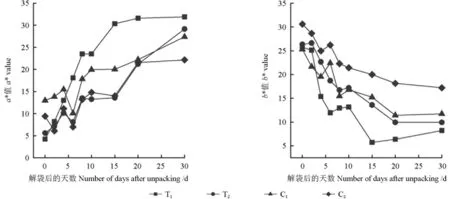
圖2 不同處理的果皮色澤參數(shù)a*、b*值的變化趨勢(shì)Fig. 2 The change trend of peel color parameters a* and b* values under different treatments
不同處理的果皮色澤參數(shù)C*和h°值的變化趨勢(shì)如圖3所示。
由圖3可知,隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋果皮的色澤飽和度C*值均呈先下降后上升的變化趨勢(shì),但其變化轉(zhuǎn)折的時(shí)間有所不同:套袋的芽變優(yōu)系果皮的C*值,解袋后0~4 d呈下降趨勢(shì),解袋后6 d開(kāi)始上升;而套袋的長(zhǎng)富2號(hào)果皮的C*值,解袋后15 d才開(kāi)始上升。未套袋的芽變優(yōu)系果皮的C*值,解袋后10 d開(kāi)始升高;而未套袋的長(zhǎng)富2號(hào)果皮的C*值,解袋后20 d快速下降,解袋后30 d又上升。解袋后30 d,套袋和未套袋的芽變優(yōu)系果皮之間、套袋和未套袋的長(zhǎng)富2號(hào)果皮之間其C*值均無(wú)顯著差異,套袋和未套袋的果皮的C*值由高到低依次為T(mén)1>T2>C1> C2。
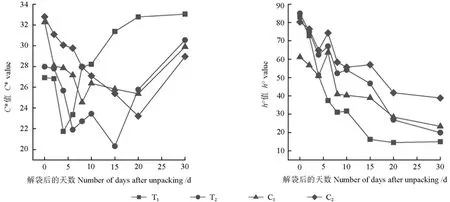
圖3 不同處理的果皮色澤參數(shù)C*、h°值的變化趨勢(shì)Fig. 3 The change trend of peel color parameters C* and h° in different treatments
圖3表明,解袋后0 d,套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的h°值均高于未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的h°值。隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋果皮的色調(diào)角h°值均呈下降趨勢(shì)。套袋的芽變優(yōu)系果皮的h°值,解袋后0~8 d急速下降,解袋后15~30 d變化不大;套袋的長(zhǎng)富2號(hào)果皮的h°值,分別在解袋后6、10 d有所回升后又下降。套袋的芽變優(yōu)系果皮的h°值整體低于套袋的長(zhǎng)富2號(hào)的果皮的h°值,解袋后0~2 d兩者間不存在顯著差異,而解袋后4~30 d其差異極顯著(P<0.01);未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的色調(diào)角h°值均呈下降趨勢(shì),未套袋的芽變優(yōu)系的果皮h°值整體低于未套袋的長(zhǎng)富2號(hào)的果皮h°值。未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)的果皮h°值在解袋后0~4 d均下降,而解袋后6 d均上升至最高值后又開(kāi)始下降。至解袋后30 d,套袋和未套袋的果皮色調(diào)角由高到低的順序?yàn)椋篊2>C1>T2>T1,套袋和未套袋的芽變優(yōu)系果皮之間、套袋和未套袋的長(zhǎng)富2號(hào)果皮之間其h°值差異極顯著(P<0.01)。
2.2 果皮中葉綠素與花青苷含量的變化趨勢(shì)
不同處理的果皮中葉綠素、花青苷含量的變化趨勢(shì)如圖4所示。
由圖4可知,隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋的果皮中花青苷的含量整體均呈上升趨勢(shì),其花青苷含量由高到低依次為T(mén)1>C1>T2>C2。套袋和未套袋的芽變優(yōu)系果皮中花青苷的含量均顯著高于套袋和未套袋的長(zhǎng)富2號(hào)的。套袋的芽變優(yōu)系果皮中花青苷的含量為0.389 mg/g,而套袋的長(zhǎng)富2號(hào)果皮中花青苷的含量?jī)H為0.165 mg/g。套袋和未套袋的花青苷含量均在形成第1個(gè)峰值后短暫下降然后又急速上升,但其峰值出現(xiàn)的時(shí)間不同,套袋的芽變優(yōu)系果皮中花青苷的含量在解袋后15 d才形成第1個(gè)峰值,而套袋的長(zhǎng)富2號(hào)果皮中花青苷的含量峰值則出現(xiàn)較早,在解袋后6 d就出現(xiàn)了第1個(gè)峰值;未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中花青苷的含量峰值均出現(xiàn)在解袋后6 d之時(shí),套袋和未套袋果皮的花青苷含量均在解袋后30 d達(dá)到最高值。
套袋和未套袋的果皮中葉綠素的含量均較低且其變化趨勢(shì)基本一致,套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)的葉綠素含量極顯著低于未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)的(P<0.01)。解袋后0~30 d,套袋的芽變優(yōu)系與套袋的長(zhǎng)富2號(hào)果皮之間其葉綠素含量不存在顯著差異,而未套袋的芽變優(yōu)系與未套袋的長(zhǎng)富2號(hào)果皮之間其葉綠素含量卻存在極顯著差異(P<0.01)。套袋和未套袋的果皮中葉綠素的含量均較低且波動(dòng)不大,葉綠素含量下降的時(shí)間與花青苷大量合成的時(shí)間相近。
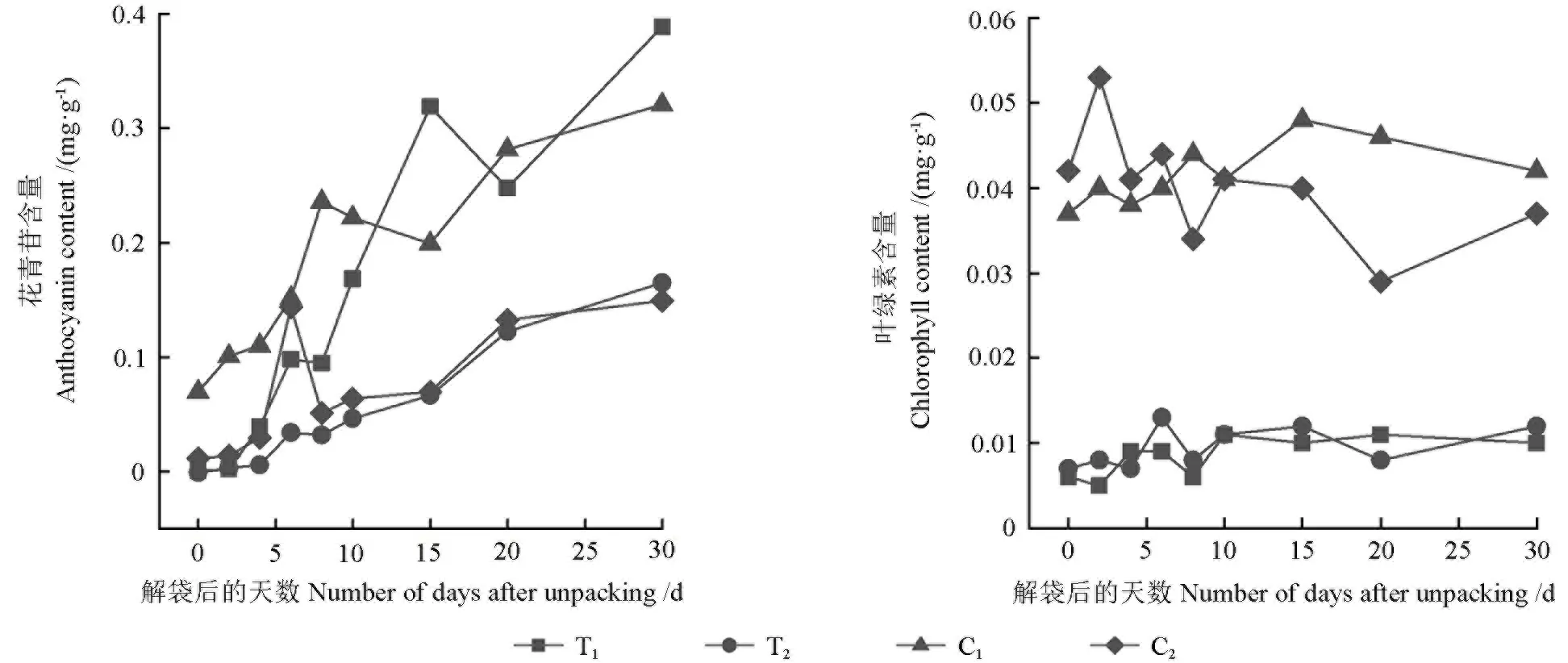
圖4 不同處理的果皮中花青苷和葉綠素的含量變化趨勢(shì)Fig. 4 The change trend of anthocyanin and chlorophyll content in peel of different treatments
2.3 果皮中花青苷合成相關(guān)酶活性的變化趨勢(shì)
不同處理的果皮中的花青苷合成相關(guān)酶活性的變化趨勢(shì)如圖5所示。
由圖5A可知,套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中苯丙氨酸解氨酶(PAL)活性的波動(dòng)范圍(7~10 U/g)均較小,解袋后30 d其PAL活性較解袋后0 d的均略有上升,其與未套袋的芽變優(yōu)系及長(zhǎng)富2號(hào)果皮PAL活性的變化趨勢(shì)均相似。
由圖5B可知,果皮中查爾酮異構(gòu)酶(CHI)與PAL活性的變化趨勢(shì)不同。套袋的芽變優(yōu)系果皮中CHI的活性,解袋后緩慢上升,然后短暫下降,最后又上升,這一變化趨勢(shì)與其花青苷的積累表現(xiàn)一致;而套袋的長(zhǎng)富2號(hào)果皮中CHI的活性,解袋后0~8 d呈現(xiàn)出“M”曲線型的變化趨勢(shì),解袋后10~30 d呈平緩上升趨勢(shì)。未套袋的芽變優(yōu)系果皮中CHI的活性,解袋后8 d即達(dá)到最高值,此后略有降低但仍高于解袋后0 d的;未套袋的長(zhǎng)富2號(hào)果皮中CHI的活性,解袋后0~10 d呈現(xiàn)出“W”曲線型的變化趨勢(shì),這與套袋的長(zhǎng)富2號(hào)果皮中CHI活性的變化趨勢(shì)相反。套袋和未套袋果皮中CHI的活性,解袋后0~30 d總體均呈上升趨勢(shì)。套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中CHI的活性均明顯低于未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中CHI的活性。
由圖5C可知,果皮中二氫類黃酮還原酶(DFR)活性的變化趨勢(shì)與CHI活性的變化趨勢(shì)基本一致。套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中DFR的活性,解袋后6 d均達(dá)到峰值,然后均下降,之后又都緩慢上升;未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中DFR的活性,分別在解袋后6和4 d達(dá)到峰值,然后均開(kāi)始下降。套袋和未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中DFR活性的變化趨勢(shì)不一致,套袋有可能使其DFR活性變?nèi)酢?/p>
由圖5D可知,套袋的芽變優(yōu)系果皮中類黃酮3,5-糖苷轉(zhuǎn)移酶(UFGT)的活性呈上升趨勢(shì),而套袋的長(zhǎng)富2號(hào)果皮中UFGT活性的變化平緩,此二者的UFGT活性均在解袋后15 d時(shí)達(dá)到最高值;未套袋的芽變優(yōu)系果皮中UFGT的活性呈平緩上升趨勢(shì),而未套袋的長(zhǎng)富2號(hào)果皮中UFGT的活性卻呈下降趨勢(shì)。套袋和未套袋的芽變優(yōu)系果皮中UFGT的活性均呈上升趨勢(shì),而套袋和未套袋的長(zhǎng)富2號(hào)果皮中UFGT的活性變化均較小。由此推測(cè),芽變優(yōu)系果皮著色優(yōu)異或許是由其UFGT活性較高所致。
2.4 果皮色澤參數(shù)值、色素含量和花青苷合成相關(guān)酶活性之間的相關(guān)性分析
果皮色澤參數(shù)值、色素含量和花青苷合成相關(guān)酶活性之間的相關(guān)性分析結(jié)果見(jiàn)表1。由表1可知,芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的L*值與a*值、花青苷含量、葉綠素含量、PAL活性、CHI活性之間均呈極顯著負(fù)相關(guān)(P<0.01),而與b*值、h°值之間均呈極顯著正相關(guān)(P<0.01);a*值與b*、h°值之間均呈極顯著負(fù)相關(guān)(P<0.01),而與花青苷含量呈極顯著正相關(guān)(P<0.01),與PAL、CHI、UFGT活性之間均呈顯著正相關(guān)(P<0.05);b*值與h°值之間呈極顯著正相關(guān)(P<0.01),其相關(guān)系數(shù)最高,達(dá)到0.952,而與花青苷含量呈極顯著負(fù)相關(guān)(P<0.01),與UFGT活性呈顯著負(fù)相關(guān)(P<0.05);h°值與花青苷含量呈極顯著負(fù)相關(guān)(P<0.01),而與CHI活性呈顯著負(fù)相關(guān)(P<0.05);花青苷含量與CHI、UFGT活性之間均呈極顯著正相關(guān)(P<0.01);葉綠素含量與CHI活性呈顯著正相關(guān)(P<0.05)。
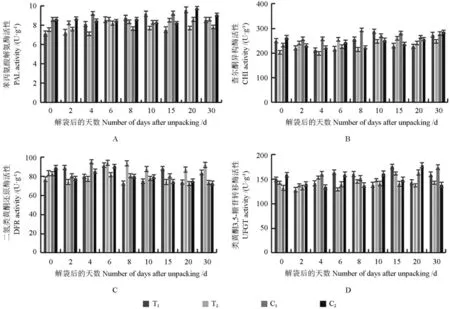
圖5 不同處理的果皮中花青苷合成相關(guān)酶活性的變化趨勢(shì)Fig. 5 The change trend of enzyme activity related to anthocyanin synthesis in peels of different treatments
3 討 論
不同品種蘋(píng)果解袋后其果皮色澤參數(shù)的變化趨勢(shì)不同。閆玖英[18]研究了澳洲青蘋(píng)、金冠、新紅星蘋(píng)果果實(shí)解袋后果皮色澤參數(shù)的變化情況,結(jié)果發(fā)現(xiàn):3個(gè)品種蘋(píng)果果實(shí)解袋后果皮的L*、b*、h°值均呈下降趨勢(shì),但其下降的快慢不同,新紅星的下降速度最快,澳洲青蘋(píng)的次之,金冠的最慢;而其a*值均呈上升趨勢(shì),新紅星的上升最快,澳洲青蘋(píng)的次之,金冠的最慢;解袋后澳洲青蘋(píng)和金冠的C*值變化都不大,而新紅星的C*值卻呈先下降后上升的變化趨勢(shì)。遲馨等[10]研究發(fā)現(xiàn),解袋后富士蘋(píng)果果皮的L*和a*值均呈上升趨勢(shì),而其b*和C*值的變化均較不穩(wěn)定,其h°值卻呈穩(wěn)步下降趨勢(shì)。試驗(yàn)結(jié)果顯示,隨著解袋時(shí)間的延長(zhǎng),套袋和未套袋的芽變優(yōu)系與長(zhǎng)富2號(hào)果皮的L*、b*和h°值均表現(xiàn)出下降趨勢(shì),而其a*值卻均呈上升趨勢(shì),其C*值在解袋后0~6 d表現(xiàn)出下降趨勢(shì),解袋后8~30 d卻均呈上升趨勢(shì)。這一結(jié)果與閆玖英[18]的研究結(jié)果基本一致。芽變優(yōu)系與長(zhǎng)富2號(hào)果皮的L*、b*和h°值,套袋的均低于未套袋的;而芽變優(yōu)系與長(zhǎng)富2號(hào)果皮的a*值,未套袋的均低于套袋的。套袋的芽變優(yōu)系果皮的a*值上升最快,且整體高于套袋的長(zhǎng)富2號(hào)的。解袋后30 d,套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮的a*、b*、h°值與未套袋的果皮a*、b*、h°值之間其差異均極顯著(P<0.01),而與其L*、C*值之間均無(wú)顯著差異,這表明套袋對(duì)果皮紅綠色度、黃藍(lán)色度、色調(diào)角均有顯著影響,但對(duì)果皮的亮度、色澤飽和度的影響都不大。相關(guān)性分析結(jié)果表明,花青苷含量與L*、b*和h°值之間均呈極顯著負(fù)相關(guān)(P<0.01),而與a*值呈極顯著正相關(guān)(P<0.01)。這一結(jié)果說(shuō)明,套袋能顯著提高果面的光潔度。
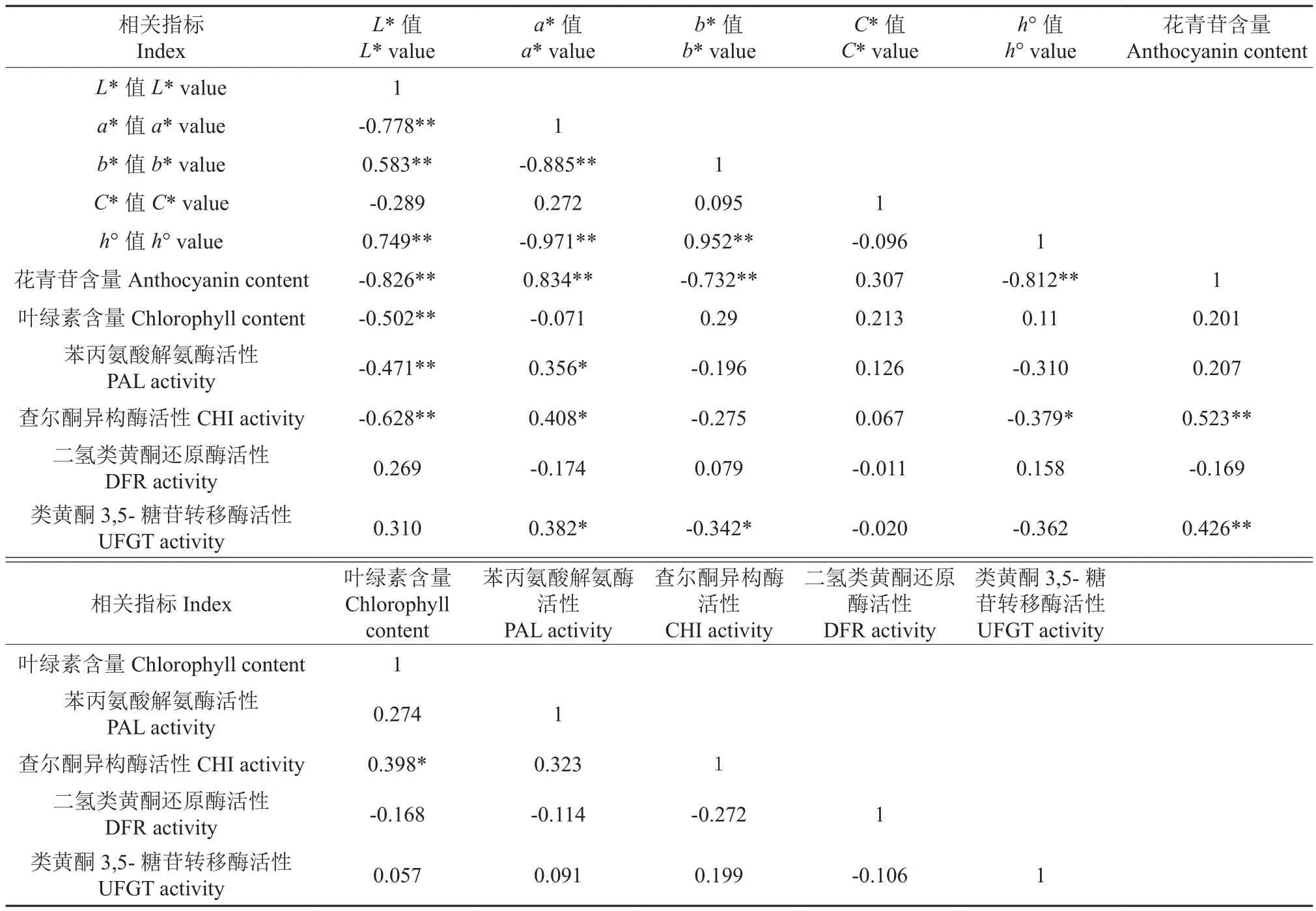
表1 果皮色澤參數(shù)值、色素含量和花青苷合成相關(guān)酶活性之間的相關(guān)性分析結(jié)果?Table 1 Results of correlation analysis among peel color parameters, pigment content and enzyme activities related to anthocyanin synthesis
套袋為果實(shí)的生長(zhǎng)發(fā)育提供了一個(gè)微域環(huán)境。套袋后光極度減弱,花青苷合成受阻,葉綠素和簡(jiǎn)單酚類的合成也受到抑制,導(dǎo)致果皮黃化,從而改變果皮對(duì)光的敏感性。解袋后的黃化果實(shí)受光后果皮中花青苷的含量迅速增加[19]。王甜元等[20]的研究結(jié)果表明,解袋5 d后果皮中花青苷的含量升高。研究中發(fā)現(xiàn),濃紅的芽變優(yōu)系果皮中花青苷的含量在解袋后15 d時(shí)達(dá)到了第1個(gè)峰值,短暫下降后又回升,解袋后30 d達(dá)到最高值;而長(zhǎng)富2號(hào)果皮中花青苷的含量,自解袋后0 d就開(kāi)始平穩(wěn)增加,解袋后30 d時(shí)也達(dá)到了最高值。程存剛等[21]的研究結(jié)果表明,套袋能明顯地影響蘋(píng)果果皮中花青苷的含量,采收時(shí)套袋蘋(píng)果果皮中花青苷的含量為對(duì)照的2.6倍。試驗(yàn)結(jié)果表明,采收時(shí)套袋的芽變優(yōu)系果皮中花青苷的含量,與長(zhǎng)富2號(hào)果皮中花青苷的含量相比,增加了21.2%,說(shuō)明套袋能促進(jìn)其對(duì)花青苷的積累。而未套袋的芽變優(yōu)系果皮的著色依然優(yōu)異,這為免袋栽培提供了可能。芽變優(yōu)系果皮中花青苷的含量始終高于長(zhǎng)富2號(hào)的,而其葉綠素的含量差異不大,這說(shuō)明紅色的呈現(xiàn)是花青苷含量明顯升高所致,而與葉綠素含量無(wú)關(guān),這與劉曉靜等[8]的研究結(jié)果一致。
花青苷的合成與PAL、CHI、DFR、UFGT等酶的活性都有一定的關(guān)系。華星等[22]的研究結(jié)果表明,藍(lán)莓PAL酶活性的變化趨勢(shì)與花青苷的積累相反,果實(shí)發(fā)育前期酶活性較高,但此時(shí)花青苷的積累較少,而當(dāng)果實(shí)發(fā)育至成熟期花青苷大量積累時(shí),酶活性卻逐漸下降。PAL酶不是藍(lán)莓花青苷合成的關(guān)鍵酶。但是,靳韋等[23]認(rèn)為,隨著果皮中花青苷含量的增加,果皮中PAL的活性也增強(qiáng),PAL活性與花青苷含量間呈正相關(guān)。PAL是否為花青苷合成的關(guān)鍵酶一直存在爭(zhēng)議[24]。張黨權(quán)等[25]認(rèn)為,CHI的表達(dá)量會(huì)直接影響花青苷的合成。有研究者指出,CHI活性的上升趨勢(shì)與花青苷含量的增加趨勢(shì)一致[17];但也有研究者證明,CHI活性與果皮中花青苷的合成的關(guān)系不密切[12,26]。Murray等[15]認(rèn)為,常春藤成年組織中DFR活性的缺乏是其花青苷積累的限制因素。蘋(píng)果[27]、草莓[28]和荔枝[12]中花青苷的含量與UFGT的活性均呈明顯的正相關(guān),說(shuō)明其二者間均有密切關(guān)系。從不穩(wěn)定的花青素轉(zhuǎn)變?yōu)榉€(wěn)定的花青苷,可能是花青苷合成的極其關(guān)鍵的步驟之一。研究中發(fā)現(xiàn),套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中PAL的活性在解袋后0~30 d的波動(dòng)均較小,與未套袋的變化趨勢(shì)基本一致;套袋和未套袋的芽變優(yōu)系與長(zhǎng)富2號(hào)果皮中CHI的活性均呈上升趨勢(shì);芽變優(yōu)系與長(zhǎng)富2號(hào)果皮中CHI的活性,套袋的均明顯低于未套袋的。套袋和未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中DFR活性的變化趨勢(shì)與其CHI活性的變化趨勢(shì)均相似,但套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中DFR的活性均呈上升趨勢(shì),而未套袋的芽變優(yōu)系和長(zhǎng)富2號(hào)果皮中DFR的活性均呈下降趨勢(shì);套袋和未套袋的芽變優(yōu)系果皮中UFGT的活性均呈上升趨勢(shì),而套袋和未套袋的長(zhǎng)富2號(hào)果皮中UFGT的活性均平緩波動(dòng)。相關(guān)性分析結(jié)果表明,花青苷含量與CHI、UFGT活性之間均呈極顯著相關(guān)。因此認(rèn)為,CHI、UFGT都是調(diào)控花青苷合成的關(guān)鍵酶。由此推測(cè),紅色芽變著色優(yōu)異的原因可能是花青苷含量明顯升高和UFGT活性較強(qiáng)。
套袋措施是生產(chǎn)中常用的改善果實(shí)著色的技術(shù)之一,但在實(shí)際生產(chǎn)中存在用工多、生產(chǎn)成本高、生態(tài)環(huán)境污染等一系列的問(wèn)題,單純的套袋并不能顯著改善果實(shí)著色和風(fēng)味問(wèn)題。因此,選育出可免袋栽培的優(yōu)良品種,開(kāi)展無(wú)袋化栽培,這是富士蘋(píng)果生產(chǎn)的迫切需要,也是蘋(píng)果栽培的發(fā)展趨勢(shì)。本研究?jī)H僅分析了長(zhǎng)富2號(hào)及其芽變優(yōu)系蘋(píng)果果實(shí)解袋后果皮的著色規(guī)律、花青苷合成相關(guān)酶活性的差異情況,而對(duì)其果實(shí)品質(zhì)未進(jìn)行全面評(píng)價(jià),今后仍需從其品質(zhì)與商品價(jià)值兩方面綜合評(píng)判該芽變優(yōu)系是否可以用作免袋栽培品種。
4 結(jié) 論
研究結(jié)果表明,套袋可顯著提高長(zhǎng)富2號(hào)及其芽變優(yōu)系蘋(píng)果果面的光潔度,增加果皮中花青苷的含量,降低其葉綠素含量。PAL并非花青苷合成的關(guān)鍵酶,而CHI和UFGT的活性與果實(shí)中花青苷合成的關(guān)系均密切,芽變優(yōu)系和長(zhǎng)富2號(hào)均通過(guò)增強(qiáng)CHI與UFGT的活性來(lái)促進(jìn)花青苷的合成。果皮紅色產(chǎn)生的主要原因可能是花青苷含量明顯升高與UFGT活性較高,而與葉綠素含量不相關(guān)。
