新疆新源縣野果林3種野生果樹秋播更新特性
2023-01-12 09:14:52劉立強(qiáng)李炎艷尤璐瑤曼蘇爾那斯?fàn)?/span>
經(jīng)濟(jì)林研究 2022年4期
關(guān)鍵詞:生長
程 功,劉立強(qiáng),李炎艷,尤璐瑤,曼蘇爾·那斯?fàn)枺?康
(1. 新疆農(nóng)業(yè)大學(xué) 園藝學(xué)院,新疆 烏魯木齊 830052;2. 邯鄲市農(nóng)業(yè)科學(xué)院,河北 邯鄲 056000)
新疆地處我國西北邊陲,面積廣闊,光熱資源豐富,屬于溫帶大陸性氣候,多樣而特殊的生境孕育著豐富的野生果樹資源。野果林是我國重要的植物資源寶庫,是野蘋果Malus sieversii、野杏Armeniaca vulgaris、野生櫻桃李Prunus divaricata等多種野生果樹的起源地[1-2]。野果林在新疆僅分布于伊犁地區(qū)沿天山山脈,其中蘊(yùn)藏著無數(shù)特異優(yōu)良性狀基因,對(duì)于研究果樹起源及品種改良具有重要意義,野生果樹的持續(xù)生存能力對(duì)維持野果林生態(tài)系統(tǒng)的穩(wěn)定性有重要的意義[3]。
近年來,由于病蟲害嚴(yán)重[4],加上氣候變化以及人為因素[5]的影響,野果林的生態(tài)系統(tǒng)越來越脆弱,伊犁地區(qū)野蘋果的分布面積銳減至原來的1/3[6],自然更新困難[7],嚴(yán)重影響了野果林的健康生長與恢復(fù),許多珍貴物種(如野生歐洲李)已瀕臨滅絕。野果林已經(jīng)被列入中國優(yōu)先保護(hù)生態(tài)系統(tǒng)名錄,野果林種群亟待更新。研究其主要樹種的實(shí)生更新能力和成苗特性對(duì)恢復(fù)野果林生態(tài)系統(tǒng)有重要意義。
人工更新是快速恢復(fù)野果林的重要途徑之一。直播更新造林已成為近幾年的主要造林方式,其生長形成的苗木具有根系完整、對(duì)造林地適應(yīng)性強(qiáng)、施工簡單、節(jié)約資金等優(yōu)點(diǎn),但見效慢,更新受多種因素的影響[8-9]。種子直播更新主要集中用于森林培育與恢復(fù)中,播種更新的影響因素包括樹種選擇、種子處理、整地和播種季節(jié)、播種技術(shù)和撫育管理措施[10]。溫水浸種后播種可促進(jìn)種子萌發(fā)出土,播種時(shí)應(yīng)適當(dāng)增加雜草清除面積,盡量減少對(duì)土體結(jié)構(gòu)的破壞,盡量增大活土層深度,土壤蓄水保墑效果最好。陡坡等宜采用魚鱗坑整地,緩坡處及山頂宜采用穴狀整地,退耕、撂荒梯田宜采用帶狀整地。播種方式和播種季節(jié)對(duì)種子萌發(fā)也有一定的影響。野杏的播種試驗(yàn)結(jié)果表明,氣候因子與野杏的萌發(fā)和幼苗生長有顯著相關(guān)性,在15°~43°的南坡上坡位生長最好,土層厚、礫石多、全磷含量高的土壤環(huán)境有利于種子的萌發(fā)和生長[11]。野生櫻桃李的天然更新包括種子繁殖和萌蘗,但其齡級(jí)轉(zhuǎn)化率較低,表現(xiàn)出前期生存能力較弱,中期穩(wěn)定發(fā)展,后期逐漸衰退的特點(diǎn)[12]。關(guān)于野蘋果的調(diào)查結(jié)果顯示,野蘋果以中齡階段個(gè)體較多,缺少幼樹和老樹,隨著新疆野蘋果種群個(gè)體齡級(jí)逐漸增大,種群期望壽命逐漸降低,提高新疆野蘋果幼齡個(gè)體的存活率和加強(qiáng)現(xiàn)存?zhèn)€體的保護(hù)是新疆野蘋果種群可持續(xù)發(fā)展的關(guān)鍵[13]。
野杏、野生櫻桃李和野蘋果是組成野果林最重要的果樹資源,對(duì)這些野生資源的研究主要集中在生物學(xué)特性及遺傳多樣性等方面[14-16]。本試驗(yàn)中選取野杏、野生櫻桃李和野蘋果3種野生果樹資源的種子,秋播于野果林原生境平地、陰坡和陽坡3種生境類型試驗(yàn)地中,研究其出苗規(guī)律和成苗特性,探究3種野生果樹的繁殖能力,旨在為新疆新源野果林的人工播種更新提供參考。
1 材料與方法
1.1 試驗(yàn)地概況
試驗(yàn)地位于新源縣野果林(83°36′4.92″E,43°22′44.28″N),海拔 1 402.6 m,屬于溫帶大陸性氣候。試驗(yàn)地年均氣溫9.37 ℃,極端氣溫分別為-25.42 ℃(1月28日)和32.65 ℃(7月28日),平均光合有效輻射為221.1 W/m2,年降水量556.8 mm,年內(nèi)降水分布不均,其中4—6月降雨占全年降水的49.1%。土壤pH值為7.38,0~50 cm深度土壤年均水分體積分?jǐn)?shù)為0.227,來自自然降水。播種時(shí),平地、陰坡、陽坡土壤表層水分體積分?jǐn)?shù)分別為0.296、0.248、0.229,播種后30 d內(nèi)平均氣溫分別為0.34、0.77、3.71 ℃,降水7次,共計(jì)28.8 mm。播種的平地、陰坡是衛(wèi)生采伐后的野蘋果林地,陽坡以禾本科植被為主。
1.2 試驗(yàn)材料
于秋季野果林果實(shí)成熟期,在試驗(yàn)地原生境采集野杏、野蘋果和野生櫻桃李成熟果實(shí),用水洗凈種子,選擇健康、飽滿、成熟度高的種子用濕紗布包裹后備用。
1.3 試驗(yàn)方法
1.3.1 樣地選擇與播種
于秋季土壤上凍前(11月3日),在試驗(yàn)區(qū)選擇平地(坡度4°)、陰坡(西北向,25°)、陽坡(西南向,35°)3種類型試驗(yàn)地,做1 m×1 m樣方,3次重復(fù),將3種種子分別播種于樣方中,覆土深度為1~2 cm,播種量為野杏種子300粒,野生櫻桃李種子300粒,野蘋果種子100 g(約3 400粒)。
1.3.2 出苗和成苗觀測
播種后第2年春季,樣方中有種子萌發(fā)出土?xí)r,開始記錄出苗數(shù)量,每3 d記錄1次出苗時(shí)間和成苗植株數(shù)量,待無幼苗出土?xí)r,計(jì)算出苗率(出苗數(shù)量占播種數(shù)量的比例)和成苗率(生長至4葉1心幼苗的植株數(shù)量占播種數(shù)量的比例)。生長季結(jié)束后統(tǒng)計(jì)各樣方中苗木數(shù)量作為保存數(shù)量,計(jì)算保存率(保存數(shù)量占成苗數(shù)量的比例)。
1.3.3 幼苗生長指標(biāo)和生物量測定
待無幼苗出土?xí)r,開始測量株高和基徑,此后每20 d測量1次。幼苗生長結(jié)束后于9月20日挖回整株,測定幼苗的主根長、側(cè)根長、側(cè)根數(shù)量、根鮮質(zhì)量、根干質(zhì)量,莖鮮質(zhì)量、莖干質(zhì)量,葉鮮質(zhì)量、葉干質(zhì)量。
1.4 數(shù)據(jù)處理
使用Excel 2013和SPSS 19.0軟件處理分析試驗(yàn)數(shù)據(jù)。
2 結(jié)果與分析
2.1 3種野生果樹的播種出苗規(guī)律
播種后3種試驗(yàn)地野杏出苗率的變化如圖1A所示。由圖1A可見,野杏于次年4月10日首先在陰坡出苗,平地和陽坡出苗晚3 d,陽坡出苗量稀少,平地和陰坡出苗集中在5月10日前,在此期間出苗數(shù)量分別占總出苗數(shù)量的90.2%和99.0%。
播種后3種試驗(yàn)地野生櫻桃李出苗率的變化如圖1B所示。由圖1B可見,野生櫻桃李在4月10日開始出苗,4月13日—5月4日的出苗數(shù)量占總出苗數(shù)量的80.6%,平地和陰坡出苗較多,陽坡出苗少。

圖1 播種后3種試驗(yàn)地野生果樹出苗率的變化Fig. 1 Variation of seed emergence rate after seeding in three test plots
播種后3種試驗(yàn)地野蘋果出苗率的變化如圖1C所示。由圖1C可見,野蘋果在4月10日開始出苗,在3種試驗(yàn)地的出苗率均小于1%。3種試驗(yàn)地的主要出苗期略有差異,在平地和陽坡集中在4月10—25日,在陰坡則延長至5月10日,此后出苗減緩。
不同類型的試驗(yàn)地適合不同類型種子的出苗。截至5月31日,平地和陽坡野生櫻桃李的出苗率最高,陰坡為野杏出苗率最高,平地和陰坡出苗率最低的為野蘋果,陽坡出苗率最低的為野杏,可能與種子質(zhì)量有關(guān),同時(shí)種子越冬期間易受動(dòng)物覓食搬運(yùn)及多種因素的影響。
2.2 3種野生果樹的播種成苗和保存情況
3種試驗(yàn)地野生果樹的種子成苗及其保存情況見表1。由表1可見,平地和陽坡野生櫻桃李的成苗率最高,分別為26.2%和4.2%,在平地的保存率最高,為87.6%,陽坡的保存率為52%。在陰坡,野杏的成苗率最高,其保存率也最高,分別為16.2%和82.1%。在3種試驗(yàn)地野蘋果的成苗率均低于1%,陽坡試驗(yàn)地野蘋果幼苗的保存率相對(duì)較高。同時(shí),在陽坡試驗(yàn)地成苗率最低的是野杏,其保存率也較低。
2.3 3種野生果樹實(shí)生苗的年生長動(dòng)態(tài)
在3種試驗(yàn)地野杏播種苗的生長動(dòng)態(tài)如圖2所示。由圖2可見:野杏出苗后,在陽坡的生長速度最慢,其生長主要集中在5月1日—6月10日,在此期間株高的日增長量為0.24 cm,隨后株高不再變化;平地和陰坡的株高在整個(gè)生長季均不斷增大,株高日增長量分別為0.50、0.37 cm,但在8月10日后,陰坡的株高變化減緩,平地的播種苗仍快速生長,兩者差距變大。在5月1日—6月30日,3種試驗(yàn)地野杏播種苗的基徑無明顯差異,此后陽坡播種苗的基徑增長變緩,與平地和陰坡播種苗基徑的差異變大;在8月10日后,平地與陰坡播種苗基徑的差異變大,在平地、陰坡、陽坡基徑的日增長量分別為0.040、0.036、0.018 mm。陰坡野杏播種苗的節(jié)間長在7月20日前快速增長,后保持不變,平地和陽坡野杏播種苗節(jié)間長變化的差異較小。截至9月20日,野杏的株高和基徑均為平地最高,陰坡次之,陽坡最小。
在3種試驗(yàn)地野生櫻桃李播種苗的生長動(dòng)態(tài)如圖3所示。由圖3可見,野生櫻桃李的生長主要集中在6月10日—8月30日,在此期間平地、陰坡、陽坡野生櫻桃李的平均日生長量分別為0.43、0.43、0.10 cm,至8月10日陰坡試驗(yàn)地野生櫻桃李播種苗停止生長,至8月30日之后,平地試驗(yàn)地野生櫻桃李播種苗停止生長。平地和陰坡試驗(yàn)地野生櫻桃李播種苗的基徑變化集中在5月20日—8月30日,平地的增長量最大,陽坡最小。3種試驗(yàn)地野生櫻桃李播種苗節(jié)間長的變化趨勢相似,變化較小。整個(gè)生長季中,野生櫻桃李的株高、基徑和節(jié)間長均為平地最高,陰坡次之,陽坡最小。
在3種試驗(yàn)地野蘋果播種苗的生長動(dòng)態(tài)如圖4所示。由圖4可見:在陰坡和陽坡試驗(yàn)地,野蘋果的株高在整個(gè)生長季僅增加了4.36、6.17 cm;平地的野蘋果苗在6月10日后快速生長,至8月30日結(jié)束生長,在此期間平均日生長量為0.25 cm。在3種試驗(yàn)地野蘋果的基徑變化在6月10日后出現(xiàn)差異,平地上快速增長,陰坡上基徑生長緩慢,為3種試驗(yàn)地中最小。野蘋果苗的節(jié)間長變化較小,截至9月20日,平地的節(jié)間長最大,陰坡次之,陽坡最小。

圖2 3種試驗(yàn)地野杏播種苗的生長動(dòng)態(tài)Fig. 2 Growth dynamics of wild apricot seedling in three experimental plots

圖3 3種試驗(yàn)地野生櫻桃李播種苗的生長動(dòng)態(tài)Fig. 3 Growth dynamics of wild cherry plum seedling in three experimental plots
2.4 3種野生果樹實(shí)生苗的年生物量

圖4 3種試驗(yàn)地野蘋果播種苗的生長動(dòng)態(tài)Fig. 4 Growth dynamics of wild apple seedling in three experimental plots
在3種試驗(yàn)地野杏播種苗的年生物量見表2。由表2可知,3種試驗(yàn)地1年生野杏苗高分別為77.70、58.83、23.99 cm,僅平地與陽坡的苗高有顯著差異。葉片鮮質(zhì)量和干質(zhì)量均為陰坡最大,分別為3.45、1.19 g,與陽坡差異顯著。平地的莖鮮質(zhì)量和干質(zhì)量最高,分別為12.54、6.60 g,平地與陽坡間有顯著差異。陰坡的根鮮質(zhì)量和干質(zhì)量最大,分別為9.18、4.99 g,與陽坡有顯著差異。平地的主根長最大,為29.74 cm,3種試驗(yàn)地間無顯著差異。側(cè)根長和側(cè)根數(shù)量在3種試驗(yàn)地之間無顯著差異。總鮮質(zhì)量和總干質(zhì)量均為平地最大,分別為26.65、12.60 g,陰坡次之,陽坡最少,3種試驗(yàn)地間無顯著差異。
在3種試驗(yàn)地野生櫻桃李播種苗的年生物量見表3。由表3可知,在3種試驗(yàn)地,野生櫻桃李的株高分別為78.99、46.25、19.90 cm,平地最高,陽坡最低,3種試驗(yàn)地間有顯著差異。平地的葉鮮質(zhì)量和葉干質(zhì)量最大,分別為2.10、0.74 g,平地與陰坡的葉干質(zhì)量間差異不顯著,與陽坡差異顯著。平地的莖鮮質(zhì)量和干質(zhì)量均最大,分別為13.25、7.05 g,與陰坡和陽坡有顯著差異。平地的主根長、根鮮質(zhì)量、側(cè)根長、根干質(zhì)量均最大,為6.43 g,平地的根干質(zhì)量顯著高于陰坡和陽坡,陰坡的側(cè)根數(shù)量最大且與陽坡差異顯著。平地的整株生物量均最大,總鮮質(zhì)量和總干質(zhì)量分別為25.17、11.25 g,與陰坡和陽坡有顯著差異,陰坡與陽坡的總干質(zhì)量間也存在顯著差異。
在3種試驗(yàn)地野蘋果播種苗的年生物量見表4。由表4可知,在平地、陰坡、陽坡3種試驗(yàn)地野蘋果的株高分別為26.33、9.60、9.73 cm,平地最高,陰坡最低,平地與陰坡間有顯著差異。平地的葉片鮮質(zhì)量和干質(zhì)量最大,分別為2.88、1.16 g,3種試驗(yàn)地間差異顯著。平地的莖鮮質(zhì)量和干質(zhì)量最大,分別為1.49、0.90 g,與陰坡有顯著差異。陽坡的主根長最大,為20.38 cm,與陰坡有顯著差異。平地的根鮮質(zhì)量和干質(zhì)量最大,與陰坡有顯著差異。陽坡的側(cè)根長最大,為10.82 mm,與平地差異顯著。平地的總鮮質(zhì)量和總干質(zhì)量均最大,分別為6.89、2.26 g,但3種試驗(yàn)地的干質(zhì)量間無顯著差異,平地的鮮質(zhì)量與陰坡有顯著差異。
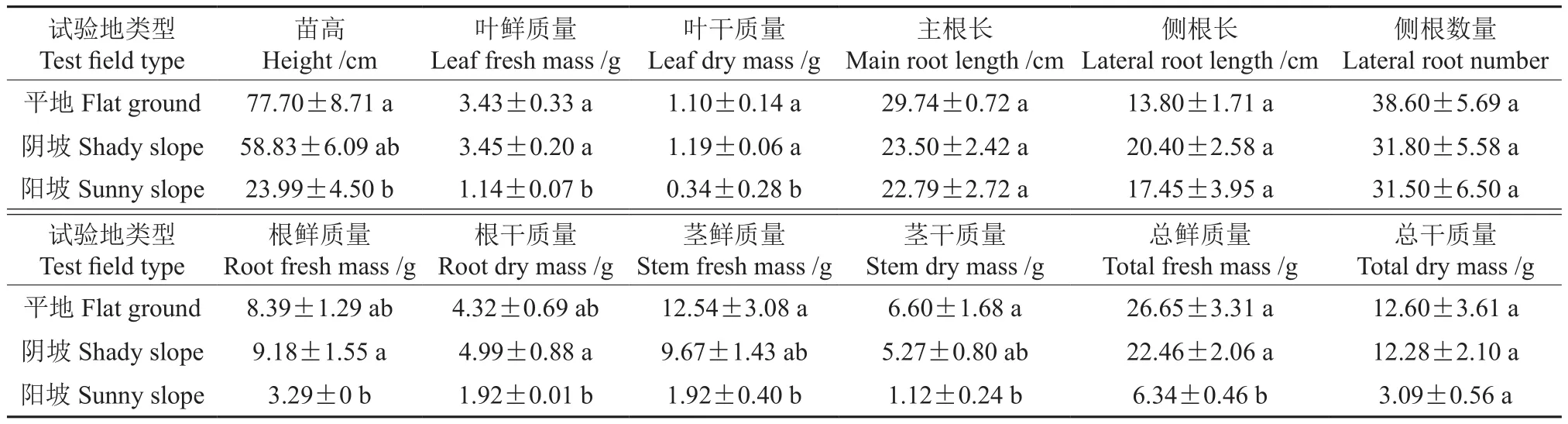
表2 3種試驗(yàn)地野杏播種苗的年生物量?Table 2 Annual biomass of wild apricot seedlings in three experimental plots
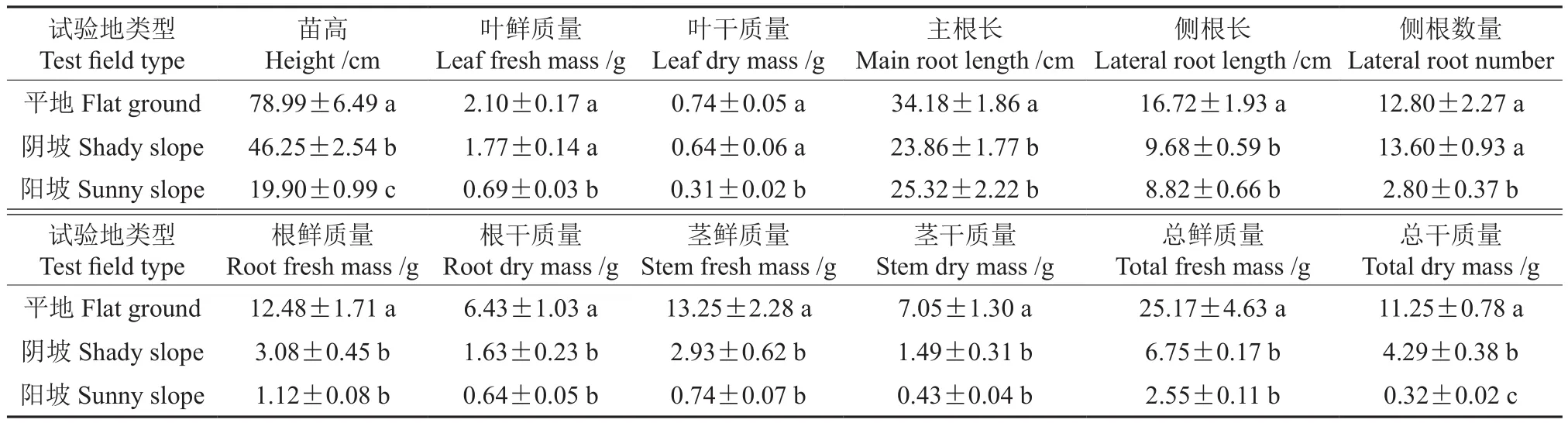
表3 3種試驗(yàn)地野生櫻桃李播種苗的年生物量?Table 3 Annual biomass of wild cherry plum seedlings in three experimental plots
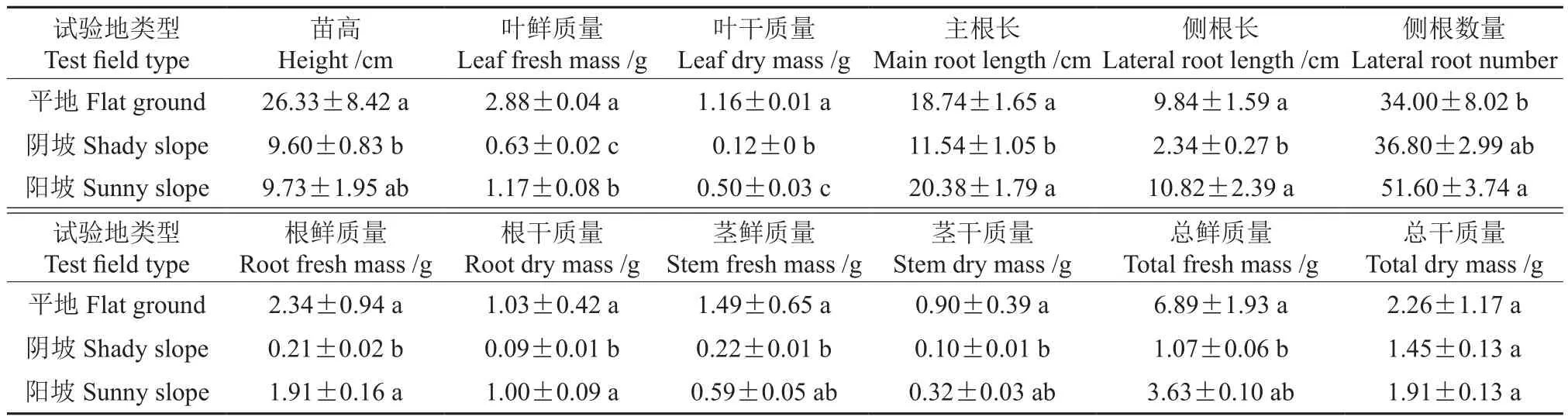
表4 3種試驗(yàn)地野蘋果播種苗的年生物量?Table 4 Annual biomass of wild apple seedlings in three experimental plots
3 結(jié)論與討論
野杏、野生櫻桃李和野蘋果的種子播種后,出苗時(shí)間集中在次年的4月上旬—5月上旬,出苗時(shí)期較一致,但出苗數(shù)量較少。野杏、野生櫻桃李和野蘋果的成苗率分別為1.0%~48.5%、4.2%~26.2%、0.5%~0.9%,保存率分別為50.0%~82.1%、52.0%~87.6%、19.4%~81.8%。野杏和野生櫻桃李的成苗率均為在陰坡試驗(yàn)地最高,3種野生果樹的生長速率和生長量均為在平地試驗(yàn)地最好。原生境中,野杏和野生櫻桃李適宜在平地和陰坡秋播更新,野蘋果不適宜秋播更新。
3.1 3種野生果樹的原生境出苗條件
人工播種是植物更新的初始階段,是種群建立和更新的重要環(huán)節(jié)。秋季播種可使種子在原生境通過休眠期,完成播種前的催芽,減少了人工種子貯藏和催芽,幼苗出土整齊,幼苗健壯,抗寒能力強(qiáng)。但野生果樹種子秋季播種后,多種因素影響種子萌發(fā)出苗,包括生物因素和非生物因素。其中,生物因素包括母樹、種子庫和其他生物。對(duì)野果林母樹的調(diào)查結(jié)果表明:野果林中野杏和野生櫻桃李樹齡較大,細(xì)菌性穿孔病危害嚴(yán)重,種源數(shù)量急劇減少,種源質(zhì)量明顯下降[17];野蘋果病蟲害嚴(yán)重,大量死亡,結(jié)實(shí)率低,種源數(shù)量受到限制,天然更新困難。因此,在原生境采種后應(yīng)進(jìn)行人工篩選,這是由采種數(shù)量和質(zhì)量決定的,人工選種方式較為簡單,會(huì)在一定程度上影響種子的出苗率。
動(dòng)物覓食是影響種子安全越冬萌發(fā)的重要因素。野果林中野生動(dòng)物種類豐富,數(shù)量較多,冬季取食較為困難,野杏和野生櫻桃李種仁較大,營養(yǎng)豐富,是野生動(dòng)物的重要食物來源。春季對(duì)播種樣方的調(diào)查結(jié)果表明,3種野生果樹的播種樣方均有不同程度的破壞,對(duì)陽坡野杏播種樣方的破壞最為嚴(yán)重,與張淵的研究結(jié)果一致[18]。另外,地表枯落物覆蓋厚度和覆土厚度是影響種子出土的重要因素[19],會(huì)顯著推遲造林樹種的萌發(fā)時(shí)間和阻礙幼苗生長,但凋落物是土壤養(yǎng)分的重要來源[20]。野果林植被豐富,生長量大,植物凋落分解后枯落物較厚,影響野蘋果幼苗的出土,對(duì)野杏和野生櫻桃李幼苗的影響較小。因此,原生境人工播種更新應(yīng)采取一定措施加強(qiáng)播種后樣方管理,以減少動(dòng)物和地表枯落物對(duì)種子萌發(fā)的影響。同時(shí),采種后可以在室內(nèi)進(jìn)行人工層積,然后在春季播種,或可減少越冬期外界環(huán)境的影響。
溫度和水分是影響種子萌發(fā)的主要?dú)庀笠蛩亍R靶印⒁疤O果、野生櫻桃李3種野生果樹種子的萌發(fā)試驗(yàn)結(jié)果表明,種子萌發(fā)過程中均有吸水特性,但適宜的層積條件不同:低溫層積100 d時(shí)野杏種子萌發(fā)率可達(dá)到90%,但層積時(shí)長不宜超過130 d[21];野蘋果種子的最佳層積條件為低溫層積75 d,發(fā)芽率可達(dá)到90%[22];野生櫻桃李的最佳萌發(fā)條件為低溫層積100 d,其萌發(fā)率可達(dá)到72%[23]。對(duì)原生境的氣象監(jiān)測結(jié)果表明:種子越冬期平均氣溫是-0.24 ℃,3種試驗(yàn)地溫差為2.73 ℃;淺層土壤溫度為2.81 ℃,3種試驗(yàn)地溫差為0.96 ℃。另外,淺層土壤中水分的體積分?jǐn)?shù)為0.27,3種試驗(yàn)地差值小于0.09。種子越冬期的氣溫、淺層土壤溫度及水分含量均可滿足種子萌發(fā)的要求,至4月初開始升溫,地溫上升稍有延遲,為種子萌發(fā)提供了適宜的條件。本試驗(yàn)中,播種苗的出土?xí)r間集中在4月10日—5月10日,時(shí)間跨度小,長勢較好。
野果林在新疆的分布較為廣泛,生境類型豐富且差異較大。種子萌發(fā)及幼苗生長受氣候等多種因素影響,本試驗(yàn)中選取新源野果林進(jìn)行人工播種,在其他地點(diǎn)的播種更新效果有待進(jìn)一步研究。同時(shí),野果林伴生種野山楂、小檗、薔薇等的人工更新也有待研究。
3.2 3種野生果樹的出苗與成苗特性
種子播種后萌發(fā)至幼苗定居是植物人工更新過程中較為敏感的階段之一,多種因素影響種子的發(fā)芽和幼苗的定居,主要有2個(gè)方面的因素:一是生理?xiàng)l件,即種子本身的內(nèi)部因素;二是環(huán)境,即種子萌發(fā)的外部因素。在伊犁大西溝,野杏的萌芽期在4月11—25日,萌芽速率最大值為5.67%~6.13%,較高的空氣溫度、土壤溫度有利于野杏種子的萌發(fā),空氣濕度與野杏的死亡率為正相關(guān)關(guān)系[24]。本試驗(yàn)中,3種野生果樹種子秋播后,次年不同試驗(yàn)地和不同類型種子的成苗率和保存率均不同。其中,平地和陽坡成苗率較高的均為野生櫻桃李,成苗率分別為26.2%和4.2%,保存率分別為97.6%和52.0%,陰坡出苗率較高的為野杏,其成苗率和保存率分別為16.2%和82.1%,野蘋果在3種試驗(yàn)地的出苗率低于1%,這可能是因?yàn)橐靶雍鸵吧鷻烟依罹哂休^大的種仁,種仁能為種子萌發(fā)和幼苗生長提供重要的營養(yǎng)物質(zhì)。野杏的種仁大于野生櫻桃李,但其成苗率和保存率低于前者,可能是因?yàn)榉N子越冬期間動(dòng)物覓食破壞嚴(yán)重,種子數(shù)量減少,成苗率降低。野蘋果種子室內(nèi)萌芽育苗的成苗率為38.1%,與秋播成苗率差異較大,野外環(huán)境播種不利于野蘋果成苗,而室內(nèi)容器育苗后移栽可不受季節(jié)限制,同時(shí)可避免原生境倒春寒等不良天氣影響,能在一定程度上提升更新效果,因此可嘗試容器苗移栽更新[25]。
3.3 3種野生果樹1年生苗的生長特性
幼苗出土后,外界環(huán)境因素成為影響幼苗健康生長的最主要因素。新源野果林位于伊犁河谷最東端,年內(nèi)降雨主要集中在4—6月[26],這有利于幼苗的生長,但該地區(qū)也易受倒春寒的影響,幼苗受凍害后死亡。所以提高幼苗抗性是播種更新的關(guān)鍵。在大西溝野杏幼苗的生長速率分3個(gè)階段,4—6月是其快速生長期,8月開始生長緩慢并停滯;野生櫻桃李的自然更新包括種子繁殖和萌蘗,但幼苗跨越齡級(jí)受到環(huán)境篩選作用的強(qiáng)烈過濾[12];野蘋果的適應(yīng)性較強(qiáng),但土壤含水量較低時(shí)能影響野蘋果幼苗的光合特性[27]。本試驗(yàn)中:3種野生果樹幼苗的生長集中在6月10日—8月30日,因?yàn)檫@個(gè)時(shí)期野果林溫度上升并趨于穩(wěn)定,豐富的降水為幼苗的快速生長提供了保障;截至生長期結(jié)束,3種果樹幼苗均為平地生長量最大,是因?yàn)槠降啬軆?chǔ)存降水,土壤含水量高,土質(zhì)肥沃,光照充足,為幼苗生長提供了適宜的環(huán)境。陽坡3種果樹幼苗的年生長量最小,是因?yàn)殛柶陆涤旰蟠嫠щy,光照充足,蒸發(fā)量大,土質(zhì)養(yǎng)分少,限制了3種果樹幼苗的生長。根據(jù)幼苗年生物量可知,3種果樹均適宜在平地和陰坡播種成苗,在陽坡則生長明顯受限。1年生苗的生長特性與試驗(yàn)地的物理環(huán)境因子、地形因子、植被因子等密切相關(guān),因此,可記錄生境微氣候,找出限制其更新的因素,為摸清幼苗成苗規(guī)律,維持其生長發(fā)育提供參考。
3.4 3種野生果樹的存活特性
3種試驗(yàn)地環(huán)境的差異導(dǎo)致生長季結(jié)束后幼苗的保存率不同。野杏和野生櫻桃李在平地和陰坡的保存率在75.7%~87.6%,兩者在陽坡的保存率僅為50.0%和52.0%。這是因?yàn)?—6月幼苗的快速生長期是試驗(yàn)地的主要降雨時(shí)期[26],土壤含水量是限制野杏和野生櫻桃李幼苗生長的主要因素,平地和陰坡利于雨水的收集,可提高土壤含水量,保證幼苗正常生長,而陽坡的日照時(shí)間長,輻射總量大,氣溫高,地表蒸發(fā)量大,致使陽坡土壤含水量減少,不能維持幼苗的正常生長,野杏和野生櫻桃李幼苗出土后生長速度快,不受雜草影響,所以平地和陰坡適合野杏和野生櫻桃李播種更新。野蘋果的成苗率小于1%,說明野蘋果播種后在原生境不能成苗是影響其人工更新的主要原因,野蘋果幼苗生長緩慢,在平地和陰坡易受周圍雜草形成的高濕度環(huán)境及動(dòng)物影響,存活率較低,在陽坡則相反,野蘋果地上部分的生長不受周圍雜草影響,所以在陽坡的保存率較高,為81.8%。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
