瀕危植物輻花苣苔與同屬2種植物光合生理特性的比較
2023-01-19 07:45:08歐明燭安明態(tài)任啟飛湯升虎劉芳馬菁華陳云飛
華中農業(yè)大學學報(自然科學版) 2023年1期
關鍵詞:植物
歐明燭,安明態(tài),任啟飛,湯升虎,劉芳,馬菁華,陳云飛
1.貴州大學林學院,貴陽 550025; 2.貴州省植物園,貴陽 550004
珍稀植物的瀕危機制及保護策略研究是植物多樣性保護的重點工作之一。植物瀕危的進程是該物種的競爭力及其對立地條件適應力等互作的生態(tài)過程[1],然而植物的競爭力和適應力與其本身的生物學特性緊密相關[2]。光合作用是植物有機物質形成的基礎,對植物生長發(fā)育起著至關重要的作用[3],光合生理能反映植物的生長策略和對資源的獲取與利用能力,是植物與立地環(huán)境互作的體現(xiàn)。植物光合日變化與響應曲線是衡量植物光合能力的重要指標,葉綠素熒光參數(shù)能反映植物光合作用的“內部”特征,揭示植物葉片的光合特性[4]。
苦苣苔科(Gesneriaceae)植物多為多年生草本,我國苦苣苔科植物在喀斯特地貌地區(qū)分布較多,是特化適應石灰?guī)r地貌的一個重要類群,具有重要的觀賞價值[5]。該科植物多分布于熱帶及亞熱帶蔭蔽潮濕區(qū)域,多數(shù)植株或生長在巖石壁上,或見于林下斜坡,具有極強的耐蔭性[6],開展該科植物的光合特征研究有助于了解其光照需求,是解析其光環(huán)境適應機制的重要途徑[7]。輻花苣苔(Oreocharis esquiro?lii)是多年生草本,花冠紫色或藍色,花期8月,因其花輻射對稱被收錄在單型屬輻花苣苔屬(Thamno?charis)中[8],2011年被并入廣義馬鈴苣苔屬(Oreo?charis)[9-10],為國家一級重點保護野生植物,極狹域分布在海拔1 500~1 600 m的林下巖石上,它的存在對于研究貴州喀斯特環(huán)境演變與生物多樣性起源及進化具有重要作用。近年來,輻花苣苔居群及棲息地面積有明顯的下降趨勢,這說明了它對環(huán)境要求的嚴格程度及自身對環(huán)境適應的脆弱性。我們前期的調查表明,輻花苣苔生長在郁閉度極高的林下巖石表面[11],生境的光限制或其自身的光合能力或許是影響因素之一。都勻馬鈴苣苔(O. duyunensis)和紫花粗筒苣苔(O. elegantissima)與輻花苣苔同系廣義馬鈴苣苔屬植物,其中都勻馬鈴苣苔為特產(chǎn)于貴州的苦苣苔科植物新種,野外分布在海拔580~1 100 m的山坡林下巖石上,花冠鐘狀細筒形、藍紫色,花期8月[12]。紫花粗筒苣苔產(chǎn)于貴州、重慶,分布于海拔約600 m的潮濕懸崖上,花冠粗筒狀,紫色,花期8月[8],在開展苦苣苔科植物保育的工作中,我們發(fā)現(xiàn)都勻馬鈴苣苔和紫花粗筒苣苔在保育地具有較好的適應能力,且它們的生長物候特征與輻花苣苔較為一致。
在相同的生態(tài)條件下瀕危植物通常表現(xiàn)出生存力及適應力較差的共同特征,而多數(shù)瀕危植物的生理代謝速率,如光合速率、呼吸速率和蒸騰速率亦低于非瀕危近緣種[13]。開展瀕危植物與同屬非瀕危植物光合生理生態(tài)特性差異的研究,以瀕危植物所表現(xiàn)出的獨特生理生態(tài)特征,找出其可能存在與周圍環(huán)境不相適應的特性,有助于闡明其瀕危的內在機制,從而為制定保護方案提供科學依據(jù)。為研究瀕危物種輻花苣苔光合作用能力,進一步闡釋其瀕危機制,本研究以其同屬非瀕危植物紫花粗筒苣苔和都勻馬鈴苣苔為對照,在同等栽培環(huán)境中開展植物光合生理特性的比較,以期為揭示輻花苣苔的瀕危機制及進一步制定保護策略提供科學依據(jù)。
1 材料與方法
1.1 研究材料概況
試驗地為中國苦苣苔科植物保育中心(貴州),位于貴州省貴陽市,貴州省植物園內(106°42′E,26°34′N),海拔約1 300 m,年均氣溫15.3 ℃,相對濕度約為74%,年平均總降水量為1 174.7 mm。輻花苣苔、紫花粗筒苣苔和都勻馬鈴苣苔均為中國苦苣苔科植物保育中心繁殖栽培的成熟植株。試驗時3種植物分別選擇長勢良好一致的植株3~4株,標記健康成熟的葉片作為測試葉,測定時保持葉片自然生長角度不變,每株植物重復測定3次。
1.2 研究方法
1) 葉綠素含量測定。摘取健康功能葉片用于葉綠素含量測定,測定及計算參考Lichtenthaler等[14]的方法。
2) 葉綠素熒光參數(shù)測定。測定時間在2021年8月,選擇晴朗天氣的上午(08:30-11:30), 使用OP?TI-sciences公司的OS-5p+便攜式脈沖調制葉綠素熒光儀測定,隨機選擇生長狀況良好且相對一致植株,每株選取3~4片功能葉,每個功能葉重復測定4次,記錄各項數(shù)據(jù)。在自然光下適應20 min后測定葉綠體光系統(tǒng)Ⅱ實際光化學量子產(chǎn)量Y(Ⅱ)、表觀電子傳遞速率(electron transportation rate,ETR)。
3)光合日進程觀測。選擇晴朗天氣的08:00-18:00時,在保育基地自然環(huán)境條件下,采用美國LI-COR公司的Li-6800便攜式光合儀,測定3種植物葉片的光合日變化。每種植物選取長勢一致的3~4片功能葉,1 h測1次。測定的主要參數(shù)包括凈光合速率(Pn)、蒸騰速率(E)、胞間CO2濃度(Ci)、氣孔導度(Gs)、飽和水汽壓虧缺(vapor pres?sure deficit,VPD)等。
4)光響應曲線測定。選擇晴朗天氣的09:00-11:30測定,測定前對待測葉片進行充分光誘導,設定葉室溫度28 ℃,葉室相對濕度60%,氣體流速為500 μmol/s,CO2濃度為400 μmol/mol(用CO2小鋼瓶控制濃度),設置0~1 500 μmol/(m2·s)的14個光合有效輻射強度(photosynthetically active radiatio,PAR)梯度進行測定,各處理穩(wěn)定2~3 min后記錄數(shù)據(jù),重復3次,取平均值。根據(jù)R語言軟件(R version 4.1.1,新西蘭)采用非直角雙曲線模型對光合光響應曲線進行擬合[15]。
5)CO2響應曲線測定。與測定光響應曲線的條件相同,測定前控制光強為600 μmol/(m2·s),CO2濃度為400 μmol/mol,對待測葉片進行誘導20 min,設置10~2 100 μmol/mol 的9個CO2濃度梯度,測定葉片凈光合速率Pn對CO2濃度的響應,采用FvCB模型對光合CO2響應曲線進行擬合,并計算最大羧化速率(vcmax)和最大光合電子傳遞速率(Jmax)[16]。
6)葉片解剖結構。將測定光合作用的葉片摘下,制作石蠟切片[17],并用Nikon Eclipes E100光學顯微鏡(日本)觀察和拍照,用Image J軟件(1.8.0,美國)測定葉片各項指標。
1.3 數(shù)據(jù)處理
試驗數(shù)據(jù)均采用Excel 2007和SPSS 13.0分析處理,LSD法進行差異顯著性檢驗,采用Origin Pro(2017C SR2b.4.2.380)進行相關圖表制作。
2 結果與分析
2.1 光合色素含量及熒光特性比較
1)光合色素含量。3種植物的光合色素含量如表1所示,輻花苣苔的葉綠素a及(葉綠素a+葉綠素b)顯著低于紫花粗筒苣苔(P<0.05),與都勻馬鈴苣苔差異不顯著;3種植物的葉綠素b及類胡蘿卜差異不顯著;輻花苣苔的葉綠素a/葉綠素b顯著低于紫花粗筒苣苔和都勻馬鈴苣苔(P<0.05)。

表1 葉片光合色素含量Table 1 Leaf photosynthetic pigment content
2) 葉綠素熒光參數(shù)。由圖1可見,輻花苣苔的初始熒光(Fo)為320.5 μmol/(m2·s),顯著高于紫花粗筒苣苔和都勻馬鈴苣(P<0.05),最大光化學效率(Fv/Fm)、PSⅡ潛在光學活性(Fv/Fo)、實際光化學量子產(chǎn)量Y(Ⅱ)、表觀電子傳遞速率(ETR)在3種植物中為最小,且Fv/Fm和Y(Ⅱ)顯著低于同屬另外2種植物(P<0.05),輻花苣苔的Fv/Fo顯著低于紫花粗筒苣苔(P<0.05),但與都勻馬鈴苣苔差異不顯著;最大熒光(Fm)和表觀電子傳遞速率(ETR)與同屬另2種植物差異不顯著。
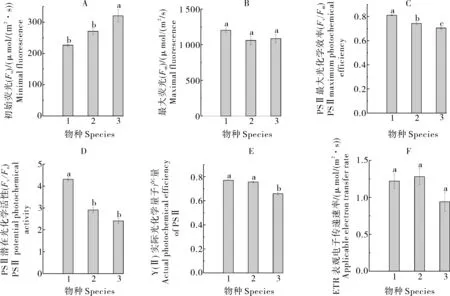
圖1 輻花苣苔與紫花粗筒苣苔和都勻馬鈴苣苔的葉片葉綠素熒光參數(shù)比較Fig. 1 Comparison of chlorophyll fluorescence parameters in leaves of O. esquirolii between O. elegantissima and O. duyunensis
2.2 光合日進程及參數(shù)比較
3種植物的凈光合速率(Pn)日變化呈現(xiàn)出較一致的雙峰曲線特點(圖2A),Pn值均在11:00時呈現(xiàn)第1個明顯的峰值,隨后Pn急劇下降,在15:00時有所回升,呈現(xiàn)第2個微峰值,在日落18:00降為最低。3種植物的光合作用主要在上午進行,在15:00會有1個小的峰值,但是總體明顯低于12:00前的光合作用。3種植物相比,輻花苣苔的日均Pn值[0.31 μmol/(m2·s)]為最低,相對波動幅度最小,紫花粗筒苣苔的Pn值[1.47 μmol/(m2·s)]為最高,波動幅度最大。
3種植物的胞間CO2濃度(Ci)趨勢大致呈“W”型,輻花苣苔和都勻馬鈴苣苔Ci的第1個波谷是在上午11:00,而紫花粗筒苣苔Ci的第1個波谷是12:00,3種植物Ci的第2個波谷均出現(xiàn)在下午15:00,3種苦苣苔在上午11:00-12:00以及下午15:00時光合作用較強且呈現(xiàn)出輻花苣苔>都勻馬鈴苣苔>紫花粗筒苣苔的趨勢(P<0.05,圖2B)。
氣孔導度(Gs)隨時間推移呈下降趨勢,14:00出現(xiàn)短暫升高后下降,呈現(xiàn)都勻馬鈴苣苔>紫花粗筒苣苔>輻花苣苔的趨勢,且輻花苣苔的Gs值顯著低于同屬另2種植物(P<0.05,圖2C)。
蒸騰速率(E)在12:00降到較低點,期間上升到14:00至最大值,說明3種植物在12:00出現(xiàn)短暫的休眠,E大小依次為都勻馬玲苣苔>紫花粗筒苣苔>輻花苣苔(P<0.05,圖2D)。

圖2 輻花苣苔與紫花粗筒苣苔和都勻馬鈴苣苔的葉片光合日變化參數(shù)比較Fig. 2 Comparison of diurnal variation parameters of leaf photosynthesis between O. esquirolii between O. elegantissima and O. duyunensis
3種植物的飽和水汽壓虧缺(VPD)趨勢基本一致,呈單峰型,VPD最小值均出現(xiàn)在08:00,最大值均出現(xiàn)在15:00(圖2E)。
2.3 響應曲線及特性參數(shù)比較
1) 光響應曲線。光響應曲線反映了植物光合速率隨光強改變而變化的規(guī)律。如圖3所示凈光合速率(Pn)隨著光合有效輻射的升高而增加,當光照強度達到飽和點后,Pn增長曲線變化趨于平緩。主要光合特性參數(shù)指標如表2所示,輻花苣苔葉片Pnmax為2.24 μmol/(m2·s),顯著低于都勻馬鈴苣苔的4.98 μmol/(m2·s)和紫花粗筒苣苔的7.01 μmol/(m2·s) (P<0.05);暗呼吸速率Rd和光補償點LCP均值分別為1.14 μmol/(m2·s)和51.85 μmol/(m2·s),顯著高于紫花粗筒苣苔和都勻馬鈴苣苔(P<0.05);光響應曲線光飽和點LSP均值為294.47 μmol/(m2·s) (P<0.05),顯著低于紫花粗筒苣苔的427.15 μmol/(m2·s),與都勻馬鈴苣苔的277.02 μmol/(m2·s)差異不顯著。

表2 3種植物葉片光響應曲線特征參數(shù)Table 2 Leaf light response curve characteristic parameters μmol/(m2·s)

圖3 輻花苣苔與紫花粗筒苣苔和都勻馬鈴苣苔的葉片光響應曲線Fig. 3 Light response curve of leaves of O. esquirolii between O. elegantissima and O. duyunensis
2)CO2響應曲線。通過擬合得到光合CO2響應曲線(圖4),由圖4可見,在CO2濃度為0~1 800 μmol/mol時,葉片凈光合速率Pn隨CO2濃度升高而增大;當CO2濃度超過1 800 μmol/mol時,3個物種的光合CO2響應曲線趨于平緩。由表3可知,輻花苣苔的最大羧化速率(vcmax)為7.63 μmol/(m2·s),顯著低于紫花粗筒苣苔的30.25 μmol/(m2·s)和都勻馬鈴苣苔的19.90 μmol/(m2·s),最大光合電子傳遞速率(Jmax)為26.04 μmol/(m2·s),也顯著低于紫花粗筒苣苔和都勻馬鈴苣苔(P<0.05)。

表3 3種植物葉片CO2響應曲線模擬的光合參數(shù)Table 3 Photosynthetic parameters simulated by leaf CO2 response curve μmol/(m2·s)
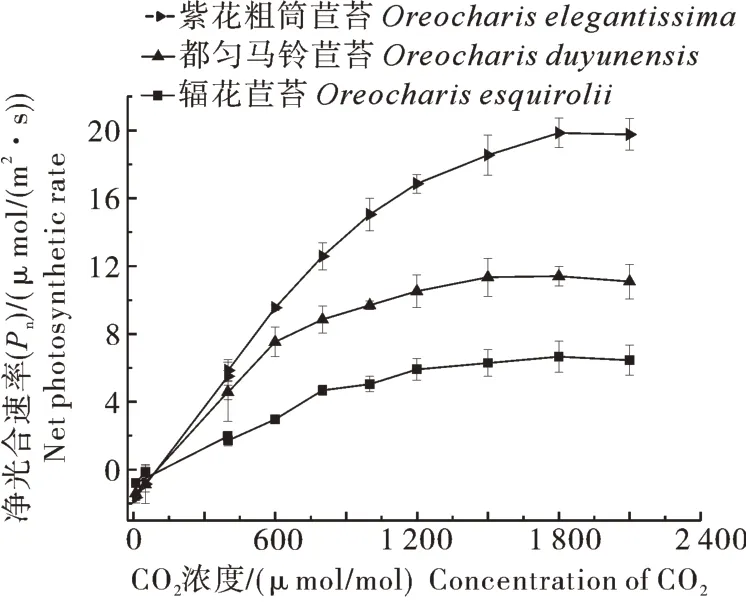
圖4 輻花苣苔與紫花粗筒苣苔和都勻馬鈴苣苔的葉片CO2響應曲線Fig. 4 CO2 response curves of leaves of O. esquirolii between O. elegantissima and O. duyunensis
2.4 葉片解剖結構的比較
經(jīng)光學顯微鏡觀察,輻花苣苔的橫切結構與紫花粗筒苣苔和都勻馬鈴苣苔存在較大差異,葉表面的絨毛密度較大,柵欄組織細胞較少(圖5)。解剖結構參數(shù)見表4,輻花苣苔平均葉片厚度(LT)為304.48 μm,顯著低于另2種植物(P<0.05);上表皮厚(UE)為37.80 μm,顯著大于紫花粗筒苣苔(P<0.05);下表皮(LE)為38.09 μm,顯著大于都勻馬鈴苣苔(P<0.05);柵欄組織厚度(PT)和海綿組織厚度(ST)分別為40.79和30.07 μm,均顯著低于另外2種植物(P<0.05);3種植物組織密實度(PT/LT)和組織疏松度(ST/LT) 差異不顯著。由此可知,輻花苣苔UE、LE較厚,PT和ST較薄,這可能與輻花苣苔長期生長在蔭蔽且土壤極薄的巖壁上的貧瘠生境有關。

表4 葉片解剖參數(shù)Table 4 Leaf anatomical parameters
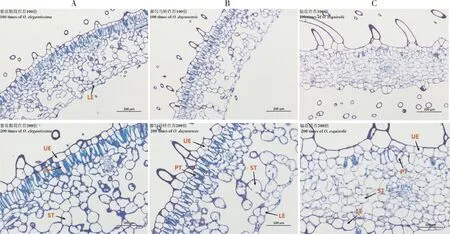
圖5 紫花粗筒苣苔(A)、都勻馬鈴苣苔(B)與輻花苣苔(C)的葉片解剖結構比較Fig. 5 Leaves anatomical comparison of O. elegantissima (A), O. duyunensis(B) and O. esquirolii(C)
3 討論
葉綠素a與葉綠素b具有捕獲和吸收光能的作用,是植物光合作用的物質基礎,其含量和比值反映植物適應和利用環(huán)境因子的重要指標。類胡蘿卜素等在弱光條件下輔助提高對總光量子的吸收[18-19]。本研究中輻花苣苔的葉綠素a及(葉綠素a+葉綠素b)含量顯著低于紫花粗筒苣苔(P<0.05),與都勻馬鈴苣苔差異不顯著;輻花苣苔的(葉綠素a+葉綠素b)含量顯著低于紫花粗筒苣苔和都勻馬鈴苣苔(P<0.05)。初步表明輻花苣苔對光能的吸收能力以及強光下的光保護能力較弱,而紫花粗筒苣苔對環(huán)境下的強光具有更強的適應能力。
葉綠素熒光參數(shù)作為評價植物光合能力的重要指標,對研究植物光系統(tǒng)及其電子傳遞過程起重要作用[20]。PSⅡ最大光化學效率(Fv/Fm)能反映葉綠素吸收光量子以及被過強光量子破壞的程度,F(xiàn)v/Fo值是反映植物光合作用過程中潛在光化學活性的重要參考指標。本研究中,模擬生境下的紫花粗筒苣苔的PSⅡ最大光化學效率Fv/Fm值最大,未受到光抑制的影響[21-22],紫花粗筒苣苔的PSⅡ潛在光化學活性Fv/Fo值顯著大于輻花苣苔和都勻馬鈴苣苔,輻花苣苔的葉綠體光系統(tǒng)Ⅱ實際光化學量子產(chǎn)量(Y(Ⅱ))及表觀電子傳遞速率(ETR)顯著低于其他2種苦苣苔,表明了輻花苣苔植株體內電子信號的傳導受抑制程度較高,進一步說明與紫花粗筒苣苔和都勻馬鈴苣苔植物相比,輻花苣苔對生境的適應能力較低。
光合作用日變化是衡量植物內在節(jié)律及對環(huán)境適應能力的重要指標,通常表現(xiàn)為“單峰”和“雙峰”型[23]。氣孔與非氣孔因子是導致光合作用下降的重要因素,由胞間CO2濃度及其變化作為主要依據(jù)[24]。飽和水汽虧缺壓(VPD)在全球溫度持續(xù)升高的趨勢下而增加[25]。VPD增加會顯著影響植株生長和發(fā)育過程,如增加蒸騰速率、降低凈光合速率、減小植株體內一些酶的活性等,這個過程是不可逆轉的[26-27]。本研究結果顯示,3種植物的凈光合速率(Pn)日變化呈現(xiàn)出較一致的雙峰曲線特點,均在11:00時呈現(xiàn)第1個明顯的峰值,在下午15:00時呈現(xiàn)第2個微峰值;胞間CO2濃度大致呈“W”型,第1個波谷出現(xiàn)在12:00,第2個波谷均出現(xiàn)在下午15:00;蒸騰速率在12:00降到較低點,均呈現(xiàn)出輻花苣苔最低的特點,3種植物均出現(xiàn)明顯的光合“午睡”現(xiàn)象,說明3種植物對高溫強光的適應能力較低,且可能是由植物的氣孔限制因素引起,導致影響植株體內碳積累過程并導致光合速率降低。在3種植物中輻花苣苔的各項光合指標均為最低,說明輻花苣苔的光合能力最弱。
光響應曲線中最大凈光合速率(Pnmax)是衡量葉片光合能力的重要指標。判斷植物耐蔭性的重要指標主要是葉片的光飽和點和光補償點,也反映了植物在生境中對光照條件的要求,3種馬鈴苣苔屬植物葉片的LSP為427.15~294.47 μmol/(m2·s),LCP為18.46~51.85 μmol/(m2·s),符合陰性植物光飽和點與光補償點的特征[28],有較低的暗呼吸速率(Rd)和光補償點(LCP)是植物能夠適應弱光環(huán)境并獲取最大物質收益適應性策略的標志[29-30]。CO2通過抑制呼吸作用及增加與羧化酶活性位點的結合來提高植物光合能力[31]。最大羧化速率(vcmax)與羧化酶活性有顯著相關性[32],其大小主要受到葉片內部羧化酶活性和數(shù)量的影響;最大電子傳遞速率(Jmax)的降低會使葉片的vcmax受到限制,是因為降低后的電子傳遞速率會限制光合磷酸化和還原輔酶Ⅱ(NADPH)的再生[33]。本研究發(fā)現(xiàn),輻花苣苔的最大凈光合速率(Pnmax)顯著低于同屬其他2種苦苣苔,光補償點(LCP)顯著高于紫花粗筒苣苔和都勻馬鈴苣苔,說明輻花苣苔對弱光的利用率較低,可能是此物種極度瀕危的因素之一,紫花粗筒苣苔葉片的光飽和點(LSP)值較高,對強光的適應能力相對較強(表3),紫花粗筒苣苔的vcmax與Jmax顯著高于都勻馬鈴苣苔與輻花苣苔,表明紫花粗筒苣苔在羧化階段RuBP再生能力更強,能更適應環(huán)境并利用光能進行碳同化,提升光合作用能力。
已有的研究表明,葉片適應弱光環(huán)境的生理特性主要為表皮厚度減小、柵欄組織及海綿組織發(fā)達[34]。影響葉片厚度的內部因素是植物葉片內的細胞體積,外部因素是對水分及營養(yǎng)的吸收,研究表明植株生長需要有較厚的表皮細胞及發(fā)達的柵欄組織保持其內部水分[35-36]。柵欄組織厚度與海綿組織厚度可以作為判斷植物生產(chǎn)力強弱的指標,柵欄組織與海綿組織越厚,越有利于提高光合利用率,光合能力越強[37-38]。上下表皮的厚度決定了植株能否有效避免強光對葉肉細胞的灼傷及有效地進行氣體交換,植物葉片的組織結構密實度、組織結構疏松度與水分利用率是反映植物適宜強光照射程度的重要指標[39]。在葉片解剖結構中,輻花苣苔葉毛密度大于紫花粗筒苣苔和都勻馬鈴苣苔,有利于增強光合作用能力(圖5),輻花苣苔葉片厚度、柵欄組織厚度與海綿組織厚度顯著低于紫花粗筒苣苔和都勻馬鈴苣苔,與凈光合速率的趨勢一致,表明輻花苣苔對弱光環(huán)境的適應能差,光能利用率低。
綜上,3種植物的光合生理特性與葉片結構之間的差異,反映其適應外界生態(tài)環(huán)境調節(jié)機制的差異。作為國家一級保護的珍稀瀕危植物輻花苣苔各項光合指標整體低于同屬的非瀕危植物紫花粗筒苣苔與都勻馬鈴苣苔,對環(huán)境的適應能力最弱,這可能是其瀕危的主要內在因素之一。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
