Plasticity of callosal neurons in the contralesional cortex following traumatic brain injury
2023-01-21 04:42:28AlexandraChovsepianLauraEmplFlorenceBareyre
中國神經再生研究(英文版) 2023年6期
Alexandra Chovsepian,Laura Empl,Florence M.Bareyre
Traumatic brain injury (TBI) represents a significant cause of disability worldwide.It creates a vast array of damaging macro-and microscopic changes in the affected brain area(s),ranging from neuronal cell death,changes in structural spine integrity and dynamics to axonal injury and overall neuronal circuit disruption,ultimately leading to functional and cognitive deficits in both humans and animal models (Nudo,2013).
Several studies in humans and animals have shown that this loss of function can often be recovered (in part) and compensated for by circuit and cellular plasticity in the brain (Nudo,2013),suggesting that the brain possesses an important potential for innate plasticity and can thus mediate some form of recovery of function.In this regard,studies have often implicated the intact contralesional cortex as an important player in recovery processes after unilateral brain injury,indicating that disrupted but intact neurons are plastic and could be able to compensate for the loss of function of the injured areas and the neighboring penumbra.
What mechanisms,and more precisely which neuronal populations,facilitate such a degree of plastic changes in the brain is still not fully understood.In this perspective,we will briefly recapitulate the current understanding on the role of the contralesional cortex following brain injury and reveal which neuronal cell types adapt in response to injury.
The role of the contralesional cortex following brain injury:Aside from the plasticity that occurs in the adjacent,intact motor cortex after focal injury,the reaction of the contralesional cortex has been extensively studied over the years.Clinical studies have used functional magnetic resonance imaging to show activity changes in the contralesional cortex following brain injury with or without rehabilitation indicating an important contribution of the contralateral hemisphere to recovery (Dodd et al.,2017).
Many experimental reports also point to rapid contralesional changes,such as hyperexcitability,that can occur as early as 1 hour after injury.Although continuous contralesional hyperexcitability can be detrimental for recovery,a subset of well-recovered patients shows contralesional motor activity and decreased functional capability upon contralesional hemisphere inhibition (Dodd et al.,2017).Both sensory and motor representations also rapidly expand in the contralesional cortex indicating a strong level of structural plastic adaptation following brain injury.Functional plasticity has also been documented.For example,upregulation of N-methyl-D-aspartic acid receptors,downregulation of γ-aminobutyric acid receptors,and changes in gene expression,in particular of genes important for axonal guidance and growth or cytoskeletal organization were observed,among others (Nudo,2013).Structural plasticity under the form of axonal or dendritic sprouting has also been reported following cortical lesions both clinically and experimentally (Napieralski et al.,1996;Ueno et al.,2012;Dancause et al.,2015;Jones,2017).We recently were able to show a significant general decrease of spine density in the contralesional cortex early after TBI.Analysis of spine dynamics of newly formedvs.eliminated spines at later time-points further showed that spine dynamics was skewed in favor of a significant increase of newly formed spines over eliminated spines,suggesting some recovery of spine density through changes in spine plasticity after traumatic cortical injury (Empl et al.,2022;Figure 1).
To date,it is not clear,which specific neuronal population adapts in the contralesional cortex and becomes plastic following cortical injury.Only a few studies have reported on the plasticity of specific neuronal populations following cortical injury.One early study reported on the sprouting of corticostriatal input from the spared contralesional cortex following cortical lesion(Napieralski et al.,1996).Ueno et al.,(2012) also reported that the contralesional corticospinal neurons can react to a cortical lesion by sprouting fibers into the denervated cervical spinal cord and by making new contact with interneurons.They also pointed out that this newly formed contact mediates recovery and is dependent on brain-derived neurotrophic factor.While such rearrangements are often key to functional recovery,it was also shown that the contralesional adaptations of cortico-reticulospinal connections might preserve low motor control at the cost of fine motor function (McPherson et al.,2018).Our latest study shows a selective sensitivity of callosal neurons to TBI,but also their potential for adaptive circuit plasticity and recovery of function after unilateral TBI,as opposed to other non-callosal neuronal populations in the intact contralesional hemisphere.We describe specific responsiveness in callosal neurons following TBI using an extensive toolbox of techniques spanning from longitudinalin vivotwo-photon imaging of spine dynamics,retrograde tracer and viral labeling,mono-synaptic circuit tracing,tissue clearing andin vivocalcium imaging to reveal the specific and time-dependent adjustment of contralesional callosal neurons to injury (Empl et al.,2022;Figure 1).
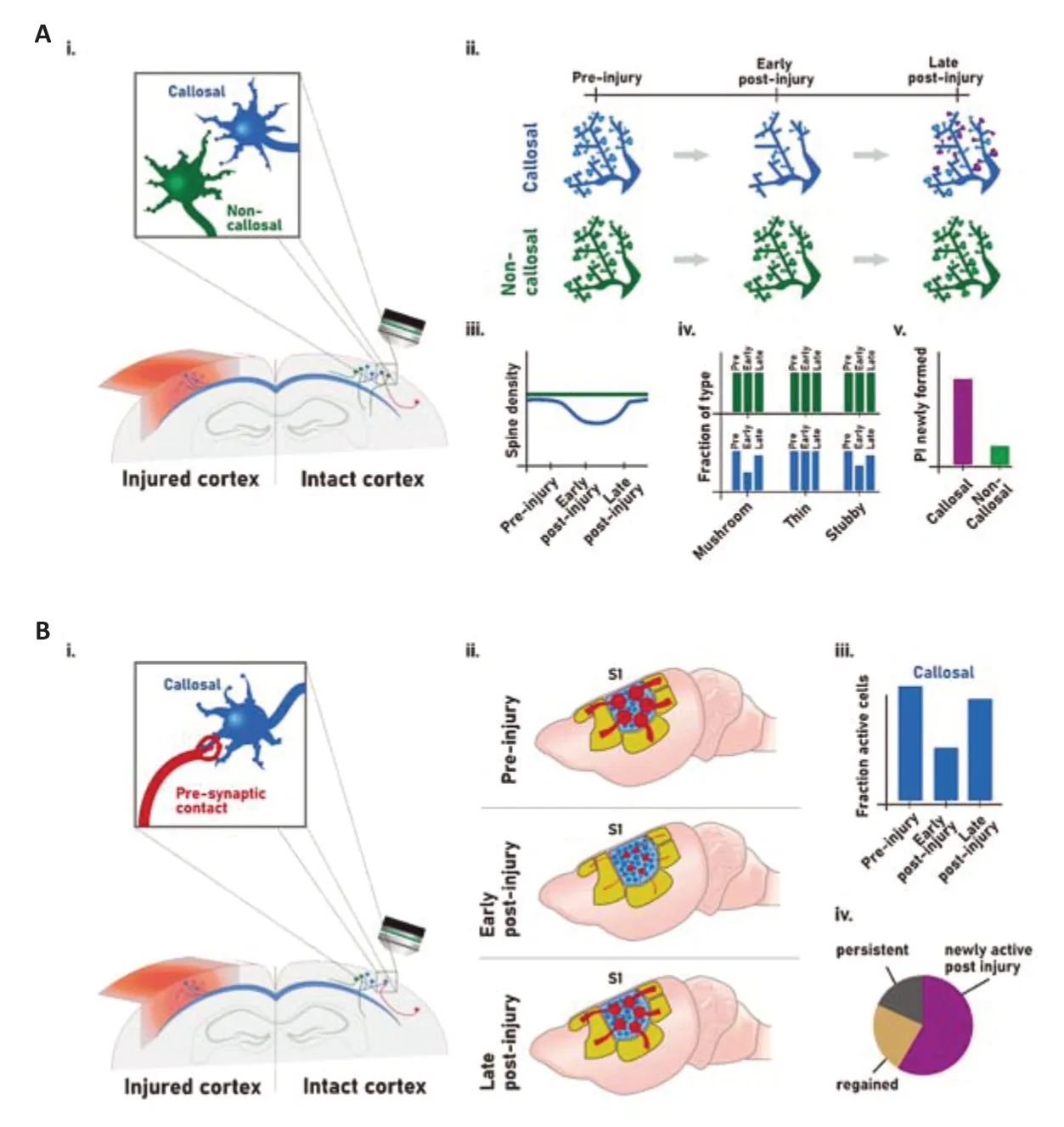
Figure 1|Contralesional changes in spines and in circuit structure and function following traumatic brain injury.
Contralesional callosal neurons have a unique vulnerability and adaptability to brain injury:Callosal neurons connect the two hemispheres of the cortex via their axons that form the corpus callosum.They primarily mediate interhemispheric communication and are responsible for higher order information processing,such as attention and language in a lateralized manner,and sensorimotor processing as a whole (Hinkley et al.,2016).As neuronal structures spanning both hemispheres,they are either directly affected by a brain injury in the case of the callosal neurons located in the ipsilesional hemisphere or directly anatomically connected to the site of a brain injury through their axons for the populations whose cell bodies are located contralesionally.In our recent paper (Empl et al.,2022),we have demonstrated the unique vulnerability and adaptability of contralesional callosal neurons to cortical injury.
Using GFP-M mice,retrograde tracing of callosal neurons and confocal microscopy,our study showed an initial spine loss early after injury on contralesional callosal neurons.The loss in spine density was followed by its recovery at later time points.Interestingly,these plastic changes in spine density were specific to callosal neurons,as noncallosal neurons remained stable over the whole study period.Furthermore,those changes were mirrored by alterations in spine morphology: The number of mushroom spines,that are thought to represent mature spines,significantly decreased early after injury before recovering at later time points while immature spines,stubby spines,transiently increased following cortical lesion.Again,non-callosal neurons were not affected indicating that those changes are specific to the callosal population.This likely indicates that less stable and immature spines are formed in response to the injury and could lead to the establishment of mature and more stable spines,reinstating “normal” spine density.To understand this process,we tracked single spines usingin vivotwo-photon microscopy in callosal,as well as noncallosal neurons.To performin vivotwo-photon imaging,spines were visualized with a retrograde adeno-associated virus expressing enhanced green fluorescent protein.Here,we could show that newly formed spines on callosal neurons were more stable than those that form on non-callosal neurons.This strengthens the idea,that while spine density may be significantly decreased in callosal neurons shortly after injury,an abundance of newly formed spines in callosal neurons,that transition into stable contacts,is able to reestablish initial spine number or even lost circuit connections.The implication of callosal neurons in the recovery process has also been demonstrated in different systems and injury paradigms.For example,following visual cortical injury the intact cortex and the corpus callosum can compensate the visuo-motor function (Celeghin et al.,2017).Plasticity of callosal inputs has also been implicated in cortical takeover following unilateral peripheral lesion under the form of whisker denervation (Petrus et al.,2020).Those adaptive processes might however be dependent on the extent of the damage inflicted and the number of cell lost after the brain injury.Indeed,previous studies have shown that there is a tight equilibrium between adaptive and mal-adaptive plasticity that is influenced by the extent of the initial brain injury.For example,Bradnam et al.(2012) demonstrated in patients with subcortical stroke that reducing contralesional M1 excitability improved upper limb control for patients with mild injuries while it worsened the outcome for moderate to severely impaired patients.Morecraftet al.,(2016) demonstrated that the size and location of the lesion differentially affected the remodeling of the ipsilateral corticospinal tract projection from the contralateral primary motor cortex.
Our study also analyzed the connectivity and functionality of contralesional callosal neurons(Empl et al.,2022).Using a combination of 3D tissue clearing and retrograde mono-synaptic circuit tracing with a modified rabies virus,we have determined that at chronic time points following cortical injury,there is little pre-synaptic connectivity to ectopic brain areas and that most brain areas connected to the callosal neurons before the injury were also found to provide input post-injury.This is an important finding as a reestablishment of the original circuitry is likely to be a general innate adaptive mechanism within callosal neurons and is potentially associated with functional recovery.The fact that spine density and circuit connectivity are restored at chronictime points suggests that those adaptive processes might be maintained over an even longer period following the brain injury.Finally,we investigated neuronal activity of contralesional callosal neurons following cortical lesions with functionalin vivotwo-photon calcium imaging using the genetically encoded calcium indicator GCaMP6m.Interestingly both callosal and non-callosal cells showed a decreased fraction of contralesional active cells early following lesion,which in turn recovered later on.Furthermore,we observed that a large part of the contralesional neurons active at the late time-point in fact only became active after the injury,indicating that existing neurons may have overtaken functions to compensate for lost activity in initially present cells.The loss of early activity in callosal neurons was further substantiated by a decrease in excitatory contacts onto dendrites of callosal neurons.Electrophysiological studies have previously demonstrated in the context of brain injury a change in excitability in the contralesional cortex with a generally increased excitability in the contralesional cortex as early as one hour following injury that might transiently return to normal (Verley et al.,2018;Dodd et al.,2017).Those distinct findings of increased or decreased excitability can be affected by the time point examined or the lesion position or extent.However,using electrophysiological measurements,functional magnetic resonance imaging or calcium imaging,TBI always leads to an interhemispheric excitability imbalance that likely impacts the ability of patients to recover from the lesion.
Our own work and that of others therefore point to a specific vulnerability and plasticity of callosal projection neurons following brain injury.As they are directly anatomically linked to the lesion site in unilateral cortical lesions,contralesional callosal neurons may compensate for their initial loss in spine number,changes in morphology and activity by slowly increasing their number of newly formed spines after injury,which in turn are able to persist longer and thereby re-establish functional connection to their formerly lost presynaptic partners.This way this particular neuronal population might help in triggering recovery.Taken together this puts callosal neurons,and their resilient potential for circuit adaption following brain injury,on the map as a new possible therapeutic target.One possibility to specifically target these neurons could be to use adenoassociated viruses-based gene therapy delivered to layer 2/3 cortical neurons using stereotactic surgery combined to the use of specific promoters targeting neurons such as a hsyn promoter.As callosal neurons seem to play a crucial role in previously reported processes of functional recovery after brain injury,they may also very well be an appropriate point of research in other injury paradigms such as stroke or other neurological disorders.
Work in FMB’s lab is supported by grants from the Deutsche Forschungsgemeinschaft (DFG,TRR274/1,ID 408885537,Project C03),the Wings for Life and the Humboldt foundation.FMB is also supported by the Munich Center for Systems Neurology (DFG,SyNergy;EXC 2145/ID 390857198).
Alexandra Chovsepian,Laura Empl,Florence M.Bareyre*
Institute of Clinical Neuroimmunology,Biomedical Building and Clinic of the Ludwig-Maximilians Universit?t München,Munich,Germany(Chovsepian A,Empl L,Bareyre FM)Munich Cluster of Systems Neurology (SyNergy),Munich,Germany (Bareyre FM)
*Correspondence to:Florence M.Bareyre,PhD,florence.bareyre@med.uni-muenchen.de.https://orcid.org/0000-0002-0917-1725(Florence M.Bareyre)
Date of submission:July 18,2022
Date of decision:September 23,2022
Date of acceptance:October 5,2022
Date of web publication:November 18,2022
https://doi.org/10.4103/1673-5374.360167
How to cite this article:Chovsepian A,Empl L,Bareyre FM (2023) Plasticity of callosal neurons in the contralesional cortex following traumatic brain injury.Neural Regen Res 18(6):1257-1258.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative CommonsAttributionNonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.
- 中國神經再生研究(英文版)的其它文章
- The relationship among amyloid-β deposition,sphingomyelin level,and the expression and function of P-glycoprotein in Alzheimer’s disease pathological process
- Translational bioengineering strategies for peripheral nerve regeneration: opportunities,challenges,and novel concepts
- The role of crm-1 in ionizing radiation-induced nervous system dysfunction in Caenorhabditis elegans
- Chitosan conduits enriched with fibrin-collagen hydrogel with or without adipose-derived mesenchymal stem cells for the repair of 15-mm-long sciatic nerve defect
- Argon preconditioning protects neuronal cells with a Toll-like receptor-mediated effect
- Glial cell line-derived neurotrophic factor and brainderived neurotrophic factor regulate the interaction between astrocytes and Schwann cells at the trigeminal root entry zone