海濱雀稗以hpt與bar基因為篩選標記的轉化體系比較研究
2023-02-07 02:22:42姜凱吳雪莉劉奕君馬越宋洋盧文杰王增裕
草業學報 2023年1期
關鍵詞:效率
姜凱,吳雪莉,劉奕君,馬越,宋洋,盧文杰,王增裕
(青島農業大學草業學院,山東 青島 266109)
海濱雀稗(Paspalum vaginatum)作為具有較強耐鹽性的暖季型草坪草,在園林綠化、運動場建植、植物修復改良鹽堿地中具有極大的應用潛力。海濱雀稗是典型的二倍體鹽土植物,可以耐受土壤中高達3.4%(電導率為54 dS·m-1)的可溶性鹽含量,在海岸的沙丘地帶、咸水池塘或河口能正常生長,并用部分海水直接灌溉[6]。海濱雀稗對于土壤環境和水質的適應性較強,在鹽堿地土壤、水質修復和沙丘固定等方面具有極大的應用潛力[6-9]。通過建立鹽生植物海濱雀稗的遺傳轉化體系,進而深入揭示耐鹽基因功能和調控機理,以便利用分子設計育種提高作物的耐鹽性,可以為農作物的耐鹽品種選育提供理論基礎,為提高鹽堿地的利用提供優質種質資源。
近年來,關于海濱雀稗耐鹽機理方面的研究主要集中在鹽脅迫下的形態結構和生理、生化響應等方面,而對關鍵耐鹽基因在耐鹽調控中的作用和通路研究較少。2008年利用海濱雀稗‘Sea Isle 1’的未成熟的花序莖段誘導產生的胚性愈傷組織,建立了再生體系[10]。通過化學誘變海濱雀稗離體再生組織,獲得了耐低溫的突變體[11]。2018年建立了海濱雀稗的遺傳轉化體系,但遺傳轉化效率較低且不穩定[12],且常因種子活力和純度低等問題影響后續研究。因此,進一步優化海濱雀稗的遺傳轉化體系,探索更加高效穩定的轉化技術,是后續進行分子生物學研究的基礎。本研究比較了兩種不同篩選劑對轉化效率的影響,同時進一步優化了轉化條件,并獲得了轉基因植株。
1 材料與方法
1.1 植物材料的消毒及處理
選取完全成熟且飽滿的海濱雀稗‘Sea Spray’品種的成熟種子為外植體,于2020年3月在青島農業大學草業育種實驗室開展試驗。種子用流水沖洗掉包衣、雜質和稃皮后,裝入50 mL離心管中,在超凈臺中進行種子的消毒處理。用75%酒精處理30 s后,使用10% NaClO溶液渦旋消毒30 min,然后用無菌水沖洗4~6次,吸除多余水分后,并用無菌濾紙吸干表面,每次接種5~10個培養皿。
1.2 菌株、質粒、引物
選用2種質粒載體(圖1A,B):內含GUS基因(β-葡萄糖醛酸糖苷酶基因)和hpt基因(潮霉素磷酸轉移酶,具有潮霉素抗性)的pCAMBIA1305.2和內含GUS和bar基因(膦絲菌素乙酰轉移酶基因,具有草丁膦抗性)的pCAMBIA3301;轉入根癌農桿菌(Agrobacterium tumefaciens)菌株AGL1中,作為農桿菌介導轉化的菌液。
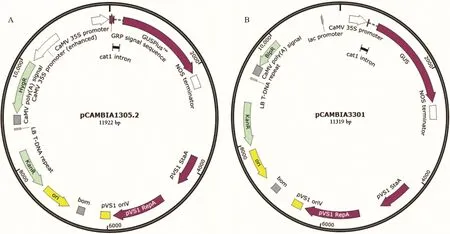
圖1 植物表達載體圖譜Fig.1 Map of plant expression vector
1.3 不同類型的培養基
在再生體系和遺傳轉化體系中,使用的培養基類型及配方見表1。
如圖3所示:隨著接種量的增加,水解度呈現先增大后減小的趨勢,在接種量為4%處達到最大值。隨著接種量的繼續增加,水解度反而降低,可能是由于山羊乳中的蛋白難以滿足過多菌體生長需求,產酸過多抑制了蛋白的水解;或菌體釋放的蛋白酶不利于在強酸條件下作用,活性被抑制。

表1 培養基的類型和配制方法Table 1 Types and preparation methods of culture medium
1.4 胚性愈傷組織的誘導繼代和再生
將消毒且吸干表面水分的滅菌種子平鋪于誘導培養基中,在28℃、黑暗條件下培養5~10 d時及時拔除幼芽。根據愈傷組織的誘導情況,在30~40 d時將產生的愈傷組織在誘導培養基中繼代。待顆粒狀的胚性愈傷組織產生后,選擇顏色鮮黃且結構緊實的愈傷組織顆粒進行繼代培養擴繁。擴繁90~180 d生長健壯的胚性愈傷組織均可作為后續遺傳轉化的材料,進行轉基因試驗。
選擇3個不同基因型的愈傷組織,挑選生長狀態良好的愈傷組織每4周繼代1次,在16、24、36、48周繼代時,分別選取部分愈傷組織接種于再生培養基中,在光照培養箱(28℃,16 h光照,8 h黑暗)中進行再生。每次進行再生試驗的愈傷組織繼代3個培養皿即3次重復,培養4~5周后統計愈傷組織的生長時間對再生率的影響,摸索出愈傷組織具有最佳再生效率的生長時間,確定愈傷組織能夠作為遺傳轉化材料的最佳時期。
1.5 胚性愈傷組織的遺傳轉化
從-80℃冰箱中分別取出含有質粒pCAMBIA1305.2和pCAMBIA3301的農桿菌菌液,分別接種于20 mL含有50 mg·L-1卡那霉素和35 mg·L-1利福平的YEP液體培養基中,在28℃搖床中,200 r·min-1振蕩培養12~18 h,當菌液濃度OD600值為0.6~1.0時,吸取農桿菌菌液加入滅菌離心管中,在離心機中以2000 r·min-1離心5 min,倒掉YEP液體培養液收集菌體。
用MS液體繼代培養基重新懸浮至菌液濃度OD600值為0.6~1.0。將挑選出的顏色鮮黃、顆粒緊實的愈傷組織浸泡在菌液中;浸染20~30 min后,吸除愈傷組織上多余的菌液,并用滅菌濾紙吸干,將愈傷組織平鋪在有滅菌濾紙的MS繼代培養基中,在28℃培養箱中黑暗條件下共培養2 d。共培養后,用含有200 mg·L-1頭孢霉素的無菌水漂洗掉愈傷組織上多余的菌液,將愈傷組織轉移到選擇繼代培養基上,28℃暗培養2~3周,有農桿菌溢菌時,要及時轉移至新的選擇繼代培養基中。每個質粒各進行了5個批次的遺傳轉化試驗,每批轉化材料接種20~30個培養皿。
1.6 篩選壓的選擇
選取繼代16周且再生率高的基因型,進行愈傷組織繼代和再生階段的最佳篩選壓試驗。將一定數量的愈傷組織分別接種到含0、40、80、100、120 mg·L-1潮霉素的繼代培養基、含0、10、20、60、80 mg·L-1潮霉素的再生培養基;含0、0.2、0.4、0.8、1.2 mg·L-1草丁膦的繼代培養基和再生培養基。生長5周后,分別移入不含篩選劑的再生培養基中生長5周后統計再生率,3次重復,以確定抑制愈傷組織再生的最低篩選劑濃度為最佳篩選壓。
1.7 遺傳轉化條件的優化
乙酰丁香酮(acetosyringone,AS)添加濃度的優化:在浸染菌液濃度OD600為0.6中浸染30 min,使用超聲波處理5 min,比較浸染菌液中添加50、100、200 μmol·L-1濃度AS對GUS瞬時表達效率的影響。
Silwet L-77(表面活性劑)添加濃度的優化:在浸染菌液濃度OD600為0.6中浸染30 min,使用超聲波處理5 min,添加100 μmol·L-1AS的轉化條件下,比較浸染菌液中分別添加0.01%、0.1%、0.3%濃度Silwet L-77對GUS瞬時表達效率的影響。
真空或超聲波處理時間的優化:在浸染菌液濃度OD600為0.6中浸染30 min,添加100 μmol·L-1AS的轉化條件下,分別比較超聲波處理或真空處理0、10、20 min,對GUS瞬時表達效率的影響。
以上每個因素的優化處理,均3次重復,共培養2 d后分別轉入篩選培養基中進行轉化體的篩選。
1.8 抗性愈傷組織篩選和再生
將共培養后的胚性愈傷組織在選擇繼代培養基上每3~4周繼代一次(暗培養4~6周);然后繼代到選擇再生培養基并轉入光照培養箱(28℃,8 h黑暗,16 h光照)培養,每3~4周繼代一次。抗性植株再生后轉移至選擇壯苗培養基繼續培養,3~5周后將抗性苗移至溫室中養護。
1.9 GUS基因瞬時、穩定表達染色
將共培養2 d后的愈傷組織和篩選12~16周的抗性愈傷組織和抗性植株葉片浸于GUS染液中,置于37℃恒溫箱中12 h,使用96%乙醇脫色后,統計GUS瞬時表達或穩定表達的結果。
1.1 0抗性植株的PCR檢測
通過CTAB法少量粗提野生型和抗性再生植株的DNA,通過snapgene軟件設計引物并交由生工生物工程(上海)股份有限公司合成。hpt基因的PCR擴增516 bp引物為:hpt-F(5′-ATGTTGGCGACCTCGTATT-3′);hpt-R(5′-CGTTATGTTTATCGGCACTTT-3′)。bar基 因 的PCR擴 增434 bp引 物 為:bar-F(5′-GCTGCCAGAAACCCACG-3′);bar-R(5′-CTGCACCATCGTCAACCAC-3′)。
1.1 1數據統計與分析
分別統計每個批次進行農桿菌介導轉化的愈傷組織數,在之后的篩選過程中,統計抗性愈傷組織數、抗性愈傷組織和抗性再生植株葉片GUS穩定表達的數量及最終經檢測的陽性轉基因植株數。

運用SPSS 25.0統計分析軟件對所有數據進行單因素方差分析,并用Duncan法進行多重比較,數據為平均值±標準差(SD);采用Excel 2019進行試驗數據的整理和制圖。
2 結果與分析
2.1 愈傷組織培養年齡對再生的影響
海濱雀稗的成熟種子在誘導培養基中誘導培養6~8周,即可誘導出黃色顆粒狀且結構緊實的胚性愈傷組織(圖2A,B)。每粒種子培養成的愈傷組織標記為一個獨立的基因型。選取最優的愈傷組織顆粒進行擴繁,用鑷子將其分割成2 mm左右的小顆粒進行培養擴繁。在愈傷組織擴繁的同時,對來自不同基因型的部分愈傷組織進行再生(圖2C,D)。通過統計繼代時間對愈傷組織再生率和褐化率的影響,可選擇具有最高再生率的基因型和愈傷組織年齡作為后續遺傳轉化的材料。
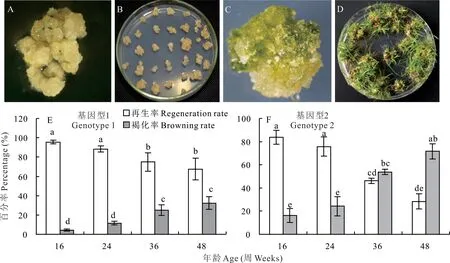
圖2 愈傷組織年齡對再生的影響Fig.2 Effect of callus age on regeneration
隨繼代培養時間的延長,所有基因型愈傷組織的再生率均降低。基因型1和2在繼代培養24周后的再生率均無顯著下降;繼代培養36周后的再生率均顯著下降,褐化率均顯著升高;繼代培養48周基因型2的再生率僅28.4%,愈傷組織的褐化率高且質量下降較快。由此可見,愈傷組織繼代培養的年齡對再生的影響因基因型的不同而不同。基因型1的愈傷組織狀態在多次繼代后,仍能保持較好的生長狀態,繼代48周的愈傷組織狀態大部分仍可以保持黃色的緊實顆粒狀,且再生率在67.6%以上,依然可以作為后續遺傳轉化的基因型。但基因型2在培養36周左右的愈傷組織質量下降明顯,已不適合做后續的遺傳轉化材料(圖2E,F)。
2.2 潮霉素篩選壓的確定
愈傷組織的繼代培養和再生過程對潮霉素敏感程度不同,因此不同篩選階段的最佳篩選壓也不同。繼代培養階段的愈傷組織對潮霉素抗性較高,在含有120 mg·L-1潮霉素的繼代培養基中培養4周后,轉移至再生培養基生長4周,仍有25.0%的愈傷組織具有再生能力(圖3A,C)。愈傷組織在含不同濃度潮霉素的再生培養基上表現得敏感度較高,當再生培養基中添加80 mg·L-1潮霉素時,再生率可降低至22.2%(圖3B,D)。由此可見,海濱雀稗胚性愈傷組織的再生階段對潮霉素較為敏感,是農桿菌介導轉化后抗性愈傷組織篩選的重要階段,可在繼代培養基中添加120 mg·L-1潮霉素篩選6~8周,然后轉入含有80或100 mg·L-1潮霉素的選擇再生培養基繼續進行篩選,并且可適當延長篩選時間,以盡量降低假陽性轉化體的存在。
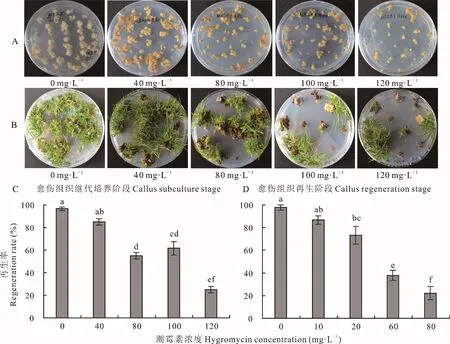
圖3 潮霉素對胚性愈傷組織繼代培養和再生的影響Fig.3 Effects of hygromycin on embryogenic callus subculture and regeneration
2.3 草丁膦篩選壓的確定
愈傷組織在繼代、再生階段對草丁膦的敏感程度均較高。接種于含有草丁膦繼代培養基中的愈傷組織的生長速度和質量均受到了明顯的抑制,愈傷組織隨著草丁膦濃度的升高,生長速度受抑制的程度越高;在含有1.2 mg·L-1草丁膦的繼代培養基中培養4周后,愈傷組織最小且顏色呈黃褐色;接種到再生培養基后僅有1.67%的愈傷組織能夠再生,且再生苗生長極慢(圖4A,C)。愈傷組織在含不同濃度草丁膦再生培養基中的敏感度相對較低,在再生培養基中添加0.4 mg·L-1草丁膦,對再生率影響較小;當添加1.2 mg·L-1草丁膦時,仍有15.0%的愈傷組織可以再生,再生苗部分生長受到較大抑制,但仍有再生苗生長較好(圖4B,C)。由此可見,海濱雀稗的胚性愈傷組織對草丁膦較為敏感的階段是在愈傷組織的繼代階段,也是農桿菌介導轉化后抗性愈傷組織篩選的關鍵階段,在繼代和再生階段均可以使用1.2 mg·L-1草丁膦作為適宜的篩選壓。
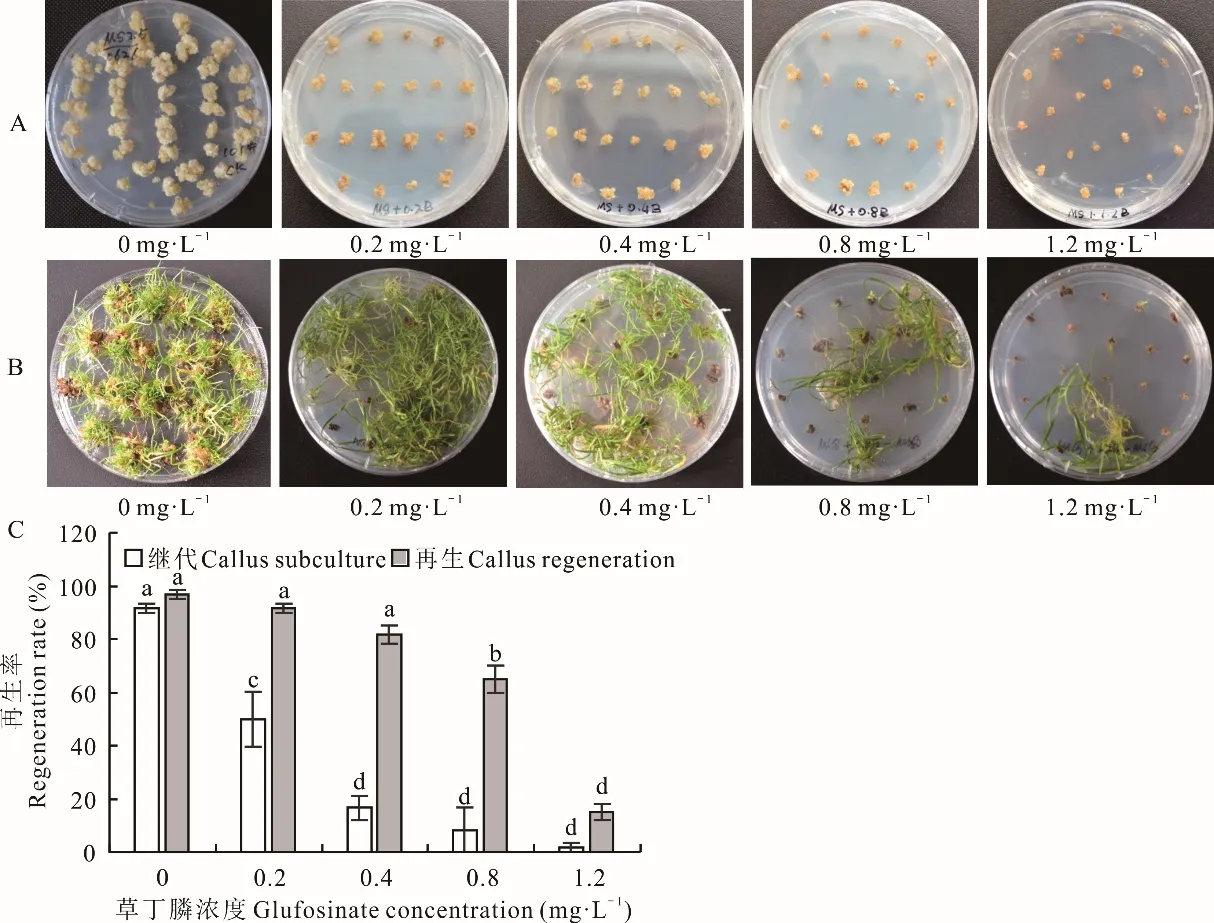
圖4 草丁膦對胚性愈傷組織繼代培養和再生的影響Fig.4 Effects of glufosinate on embryogenic callus subculture and regeneration
2.4 轉化條件的優化
為了進一步提高農桿菌介導的轉化效率,試驗通過GUS瞬時表達染色,對相關的影響因素進行了單因素條件的優化。在浸染轉化共培養2 d后,統計愈傷組織的GUS瞬時染色效率,從而摸索影響遺傳轉化的最佳處理條件。添加適當濃度AS到農桿菌的浸染液中,可以顯著提高瞬時轉化效率,但隨著添加AS濃度的增加對轉化效率的影響不顯著(圖5A,E)。在浸染的農桿菌菌液中添加低濃度的表面活性劑Silwet L-77可顯著提高瞬時表達效率,添加0.01%和0.1%的Silwet L-77后其GUS瞬時表達效率顯著提高,分別為78.07%和80.1%,雖然添加0.5%高濃度的Silwet L-77的GUS瞬時表達效率最高,但是卻對愈傷組織造成明顯的傷害,在共培養后,愈傷組織明顯褐化且表面呈現粘稠狀,極大降低了愈傷組織的后續繁殖和再生(圖5B,F)。在浸染階段,通過增加超聲波處理,可在愈傷組織表面產生更多有利農桿菌介導轉化的小傷口,可以顯著地提高瞬時表達效率。超聲波處理20 min的GUS瞬時表達效率最高且差異顯著,愈傷組織的GUS瞬時表達效率高達94.17%。因超聲波處理后愈傷組織狀態沒有明顯改變,且后續篩選過程中無法跟蹤評估更長時間的超聲波處理是否對愈傷組織的傷害程度更高,因此可以選擇20 min超聲波處理,或者適當降低處理時間(圖5C,G)。在浸染階段通過真空處理,可有效增加愈傷組織和菌液的接觸,從而顯著提高GUS瞬時表達效率,但處理10或20 min對表達效率無顯著差異,因此為盡量降低對愈傷組織的傷害,可以選擇10 min為最佳的真空處理時間(圖5D,G)。
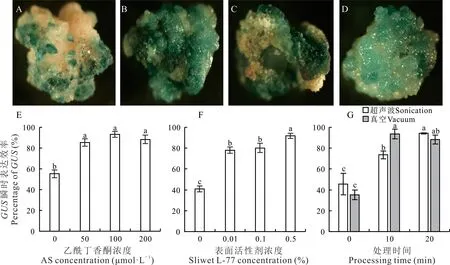
圖5 不同因素對GUS瞬時表達效率的影響Fig.5 Factors affecting on percentage of transient GUS expression
2.5 農桿菌介導的不同質粒GUS穩定表達效率和轉化效率的統計
挑選黃色顆粒狀且結構緊實的愈傷組織,在滅菌的空培養皿中分別用含有兩種不同質粒的AGL1農桿菌菌液進行浸染,在浸染菌液濃度OD600為0.6時,菌液中同時添加100 μmol·L-1AS和0.01%的Silwet L-77,浸染過程中先超聲波處理20 min,然后再真空處理10 min,浸染30 min后共培養2 d。之后分別接種于含有120 mg·L-1潮霉素、200 mg·L-1頭孢噻肟(圖6A)和含有1.2 mg·L-1草丁膦、200 mg·L-1頭孢噻肟(圖6D)的選擇繼代培養基進行抗性愈傷組織的篩選,繼代3~5次后,將抗性愈傷組織進行GUS染色呈深藍色(圖6B,E),表明GUS基因在抗性愈傷組織中獲得了穩定表達。將抗性愈傷組織再分別轉到含有80 mg·L-1潮霉素和200 mg·L-1頭孢噻肟或者含有1.2 mg·L-1草丁膦和200 mg·L-1頭孢噻肟的選擇再生培養基上,再生的同時進一步篩選,篩選過程中非抗性愈傷組織逐漸變褐死亡,而部分抗性愈傷組織能夠在選擇再生培養基中再生出健康的植株(圖6C,F)。將抗性再生的幼苗進行壯苗后,轉至溫室中單株種植(圖6G)。分別剪取部分抗性再生植株的葉片,進行GUS染色并用酒精脫色后葉片兩端仍呈深藍色(圖6H),說明GUS基因在抗性植株中穩定表達。

圖6 農桿菌介導的胚性愈傷組織的遺傳轉化及GUS染色Fig.6 Agrobacterium-mediated transformation of embryogenic calli and result of GUS staining
在以上農桿菌介導轉化的最優條件下,2種質粒各進行了5個批次的胚性愈傷組織轉化。共轉化hpt做篩選標記的愈傷組織329粒,篩選出74個克隆的抗性愈傷組織,其中55個克隆成功再生出抗性苗,其中抗性苗的葉片GUS染色呈現藍色的共48個克隆,占獲得抗性再生植株克隆數的87.3%,根據PCR鑒定的陽性抗性苗的平均轉化效率為14.46%。共轉化bar做篩選標記的愈傷組織355粒,篩選獲得抗性愈傷組織和抗性苗的數量均較低,平均轉化效率為5.65%。由此可見,以潮霉素為篩選標記的轉化效率較高,適合在海濱雀稗轉基因中使用(表2)。
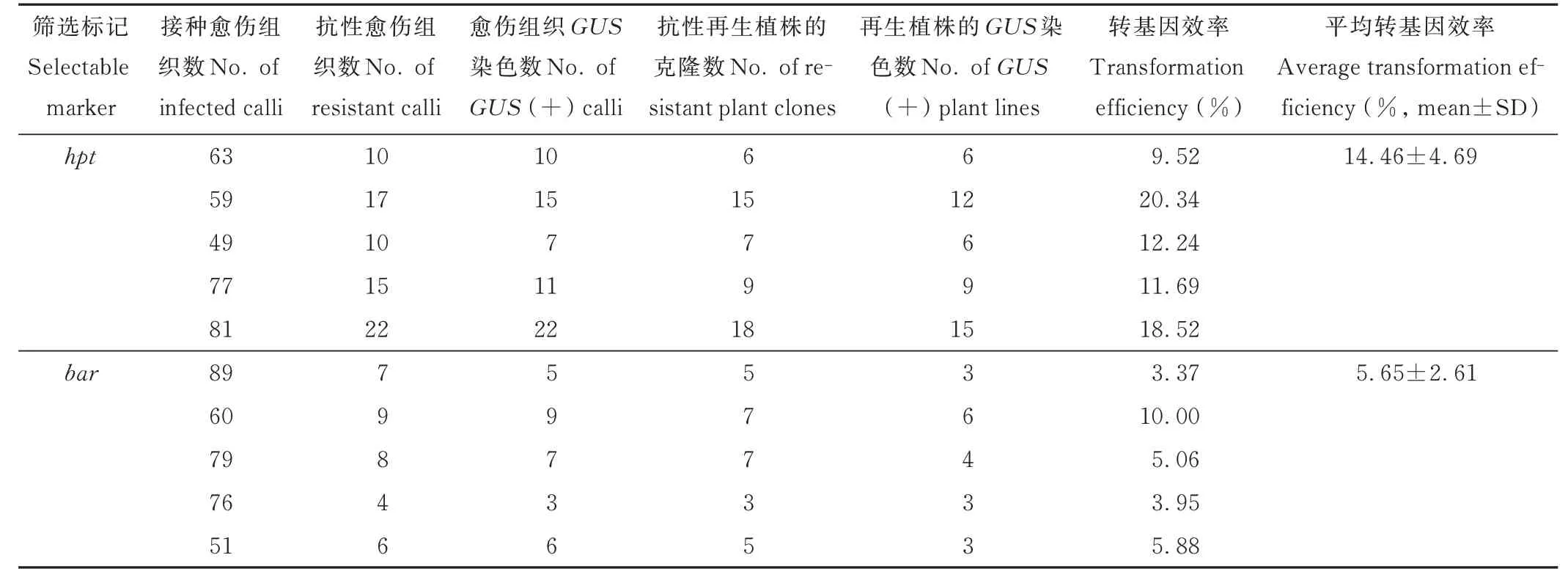
表2 兩種不同質粒的農桿菌介導的遺傳轉化效率Table 2 Summary of transformation efficiencies of Agrobacterium transformation using embryogenic callus
2.6 抗性再生植株的PCR檢測
通過分別擴增外源hpt或bar片段,對野生型和農桿菌介導轉化的抗性再生植株進行陽性檢測。結果表明,野生型植株在516或434 bp處均無擴增條帶,對獲得的葉片GUS染色為深藍色的64株具有潮霉素抗性的再生植株進行PCR檢測,除No.6沒有擴增出hpt條帶外,其他63株均在葉片中擴增出516 bp的陽性條帶,說明外源hpt基因在受體植株中獲得了穩定表達(圖7A)。在18株葉片GUS染色為深藍色的草丁膦抗性植株中,均擴增出了434 bp的陽性條帶,表明bar基因已在轉基因海濱雀稗植株的基因組穩定表達(圖7B)。
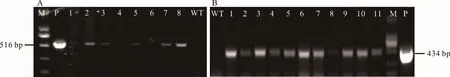
圖7 抗性再生植株的PCR檢測Fig.7 PCR test results of resistant regenerated plants
3 討論
隨著分子設計育種的發展,轉基因育種以其周期短、目標性狀精準可控等優點成為草坪草育種中的重要方法。海濱雀稗應用廣泛且具有非常強的耐鹽性,是目前最耐鹽的暖季型草坪草[13],然而目前國內育種進程緩慢,所有商業品種均來自國外[14-17]。目前,農桿菌介導的遺傳轉化具有多個突出優點,仍是禾本科轉基因的主要方法,然而禾本科因不是農桿菌的天然宿主,特別是暖季型草坪草的再生和轉化難度更大[18-19]。本研究建立了hpt和bar兩種篩選標記的遺傳轉化體系,并分別獲得了陽性轉基因植株,潮霉素作為篩選劑的平均轉基因效率可以達18.93%。海濱雀稗高效的遺傳轉化體系的建立為進一步加快育種進程奠定了堅實的基礎。
以成熟種子作為愈傷組織誘導的外植體,具有不受季節性影響并且取材方便的優點[20],目前已在多種草坪草如黑麥草(Lolium perenne)、紫羊茅(Festuca rubra)、日本結縷草(Zoysia japonica)、早熟禾(Poa annua)成功建立了離體再生體系[21]。此外,也可以采用未成熟胚、莖尖分生組織、根尖、花序、匍匐莖等部位作為禾本科的外植體來源,建立組織培養再生體系[22]。Liu等[11]以海濱雀稗‘Sea Spray’品種的成熟種子為外植體建立了再生體系,比較了不同外源物質對組織培養再生的影響。因海濱雀稗的自交不親和性,每粒種子代表著不同的遺傳信息,本研究選擇來自同一基因型的愈傷組織,比較了繼代時間對再生率的影響,最佳的可用于遺傳轉化的愈傷組織繼代時間為36周左右,既能保證有足夠數量的愈傷組織,又可以保證具有較高的再生效率。
潮霉素磷酸轉移酶基因(hpt)和膦絲菌素乙酰轉移酶基因(bar)是禾本科轉化中最常用的篩選標記基因[23-24],已在多種轉基因禾本科草中獲得成功[25-28]。海濱雀稗在不同的發育階段對潮霉素和草丁膦的敏感性有所不同。海濱雀稗的胚性愈傷組織在繼代階段對草丁膦較為敏感,是篩選的關鍵階段,1.2 mg·L-1草丁膦可作為整個篩選過程的適宜篩選壓。在繼代階段愈傷組織對潮霉素的抗性較高,在含有120 mg·L-1潮霉素的繼代培養基中仍有部分可再生,而再生階段對潮霉素的耐性較低,在80 mg·L-1潮霉素濃度的再生培養基中愈傷組織再生率較低。由此可見,海濱雀稗具有較高的潮霉素耐性,而在高羊茅(Festuca elata)、柳枝稷(Panicum virgatum)、假儉草(Eremochloa ophiuroides)和結縷草中使用的潮霉素篩選濃度在50 mg·L-1左右,并獲得了轉基因植株[29-32]。在農桿菌浸染轉化過程中,各種處理對愈傷組織都具有一定的傷害,而潮霉素對愈傷組織也具有不良影響,對陽性愈傷組織的再生具有較大壓力。因此,在選擇繼代的篩選階段使用相對低濃度的潮霉素,抑制部分假陽性愈傷組織生長的同時,可對陽性的愈傷組織生長提供緩沖的時間;而在愈傷組織選擇再生的篩選階段使用高濃度的篩選壓,由此獲得了更高的轉化效率。
遺傳轉化過程的不同處理可以明顯影響轉基因效率。本研究根據前期建立的海濱雀稗遺傳體系,在轉化條件菌液濃度OD600=0.6,輔以超聲波處理,共培養2 d的基礎上[12],通過添加100 μmol·L-1乙酰丁香酮和0.01%的Silwet L-77,并提高超聲波的處理時間至20 min,同時輔以真空處理10 min,明顯提高了海濱雀稗的平均遺傳轉化效率。以潮霉素作為篩選劑的平均轉基因效率提高至14.46%,并且還獲得了以草丁膦為篩選劑的轉基因植株。
乙酰丁香酮(AS)通過誘發農桿菌Ri或Ti質粒上Vir區基因的活化,可有效地提高轉化效率[33-34]。不同AS濃度(50~400 μmol·L-1)對不同植物受體轉化效率的影響程度不同[35]。添加50~300 μmol·L-1的AS,丁香羅勒(Ocimum gratissimum)的GUS表達效率呈升高趨勢,但添加400 μmol·L-1時表達效率下降[36]。在花生(Arachis hypogaea)轉化 的 共培養過 程的培養 基 中添加200 μmol·L-1,瞬時轉 化效率最 高 為80%[37]。在 金發草(Pogonatherum paniceum)的轉化中添加40 mg·L-1AS的轉化效率最高為67.3%[38]。海濱雀稗的轉化中通過添加50~200 μmol·L-1的AS均可獲得較高的GUS瞬時表達效率。
表面活性劑能夠在農桿菌介導轉化過程中,通過增加細胞膜通透性,提高農桿菌T-DNA插入受體細胞的效率[39-40]。植物遺傳轉化中,Silwet L-77是應用較為廣泛的一種表面活性劑,可有效提高轉化效率[41]。在菌液中添加0.01%~0.02%濃度的Silwet L-77,可提高玉米(Zea mays)幼胚愈傷組織的遺傳轉化效率[42]。本研究中,添加0.01%和0.1%濃度的Silwet L-77的瞬時轉化效率較高;濃度提高至0.5%的瞬時轉化效率最高,但愈傷組織外觀已部分或全部褐變。在玉米胚珠的轉化中添加0.1% Silwet L-77的轉化效率最高[41],本試驗結果與其相似。由此可見,添加適當濃度的Silwet L-77可有效提高瞬時轉化效率,但因受體材料的質地不同,對表面活性劑耐受力差異較大,需要通過試驗確定最佳添加濃度。
超聲波處理可通過在愈傷組織表面形成微傷口,使農桿菌更易穿透細胞壁[43];真空處理可在農桿菌浸染中增加和愈傷組織的深度接觸,從而均可有效地提高轉化效率[44]。但需根據受體材料的質地和特性,選擇合適的處理時間和強度。本研究中,在農桿菌浸染過程中分別輔以超聲波處理20 min和真空處理10 min,均可顯著提高轉化效率。
4 結論
本研究以海濱雀稗‘Sea Spray’成熟種子為外植體,進一步優化了海濱雀稗遺傳轉化體系。愈傷組織繼代36周后的再生率會顯著下降,以繼代24周的愈傷組織為轉化材料,在菌液(OD600=0.6)中添加100 μmol·L-1乙酰丁香酮和0.01%的Silwet L-77,浸染過程中輔以超聲波處理20 min或真空處理10 min,浸染30 min后共培養2 d。在繼代和再生階段使用最佳的篩選壓進行篩選,分別獲得了表達hpt和bar基因的轉基因植株。潮霉素更適合作為海濱雀稗遺傳轉化中的篩選劑,能夠獲得較高的轉化效率。
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經濟(2017年6期)2017-07-12 09:27:16
中國衛生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛生(2014年11期)2014-11-12 13:11:32
