黃酮類化合物的抗菌作用及機制研究進展
2023-02-15 01:44:18張珍珍梅秀珍徐業芬馮志新
江蘇農業科學 2023年1期
關鍵詞:黃酮
王 健, 張珍珍, 梅秀珍, 徐業芬, 馮志新,3
(1.西藏農牧學院動物科學學院,西藏林芝 860000; 2.江蘇省農業科學院獸醫研究所,江蘇南京 210014;3.南京農業大學動物醫學院,江蘇南京 210095)
在臨床上,人們通常使用抗生素來治療細菌性疾病,由于抗生素的廣泛使用,細菌對廣泛使用的抗菌藥物的耐藥性增強,導致耐藥細菌感染問題逐漸增多,嚴重影響了現有抗菌治療的療效和安全。現有抗生素的耐藥機制是由于細菌突變后,病原菌可以通過細菌外排泵加速抗生素外排,以減少藥物在細菌內擴散所需的時間[1],并改變細菌孔蛋白的結構,降低細菌對抗生素內流的滲透性[2],破壞水解酶的抗菌性,改變抗生素的結合位點[3],抗生素無法對細菌起作用。因此有必要尋找替代抗生素的新型藥物。
黃酮類化合物又稱多酚類化合物,因其具有抑制多種病原微生物(包括多藥耐藥細菌)生長的趨勢而受到關注[4]。具有多種生物活性功能,包括抑菌、抗病毒、抗腫瘤、酶抑制作用等,由于其具有顯著的抑菌活性,有替代部分抗生素的可能,已被應用于抑菌治療[5]。這類化合物可以改變膜的通透性、抑制核酸合成、破壞細胞壁、抑制細胞質膜功能和抑制細菌活性[6]。黃酮類化合物還能通過與宿主關鍵細胞信號通路等眾多細胞靶標的相互作用,發揮調控炎癥等重要作用,包括預防和治療各種細菌感染引起的疾病,如傷口感染、呼吸道感染、胃腸道疾病和尿路感染等[7]。黃酮類化合物通過核因子-κB(NF-κB)通路和絲裂原活化蛋白激酶(MAPKs)通路等途徑抑制白細胞介素-1β(IL-1β)、白細胞介素-6(IL-6)、腫瘤壞死因子-α(TNF-α)等細胞因子的釋放,最終抑制細菌導致的炎性癥狀[8]。
1 黃酮類化合物
黃酮類化合物是一類低分子量的多酚類物質,普遍存在于水果、蔬菜中。黃酮類化合物的核心結構是一個二苯丙烷(C6—C3—C6)骨架,黃酮類化合物的化學性質取決于3碳鏈的不飽和度和氧化程度,現階段已經發現了超過9 000種黃酮類化合物,根據其化學結構可以分為黃酮、黃酮醇、黃烷酮、黃烷醇、異黃酮和花青素(表1)等[9-10]。
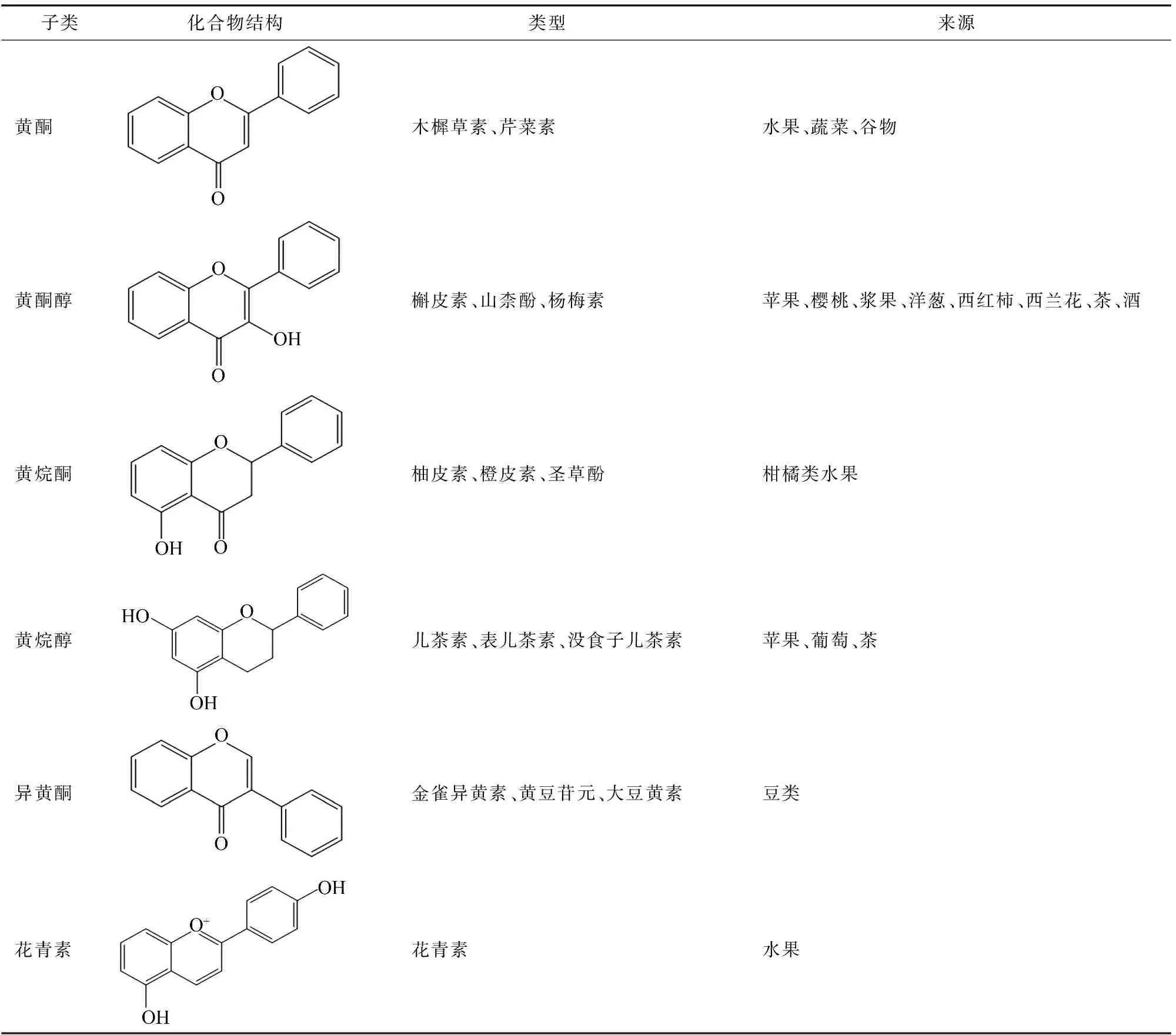
表1 黃酮類化合物分類和結構及來源
1.1 黃酮
黃酮是黃酮類化合物中最大的一個亞類之一,其特征是黃酮骨架中的C-2和C-3之間存在雙鍵,并且具有不飽和的C-3鏈。主要分布于高等植物中,研究最多的是芹菜素和木樨草素。芹菜素是黃酮類中最有代表性的化合物,存在于蔬菜、植物及其飲料中,芹菜、 歐芹中含有大量的芹菜素,具有顯著的抗菌作用、抗炎作用和抗痙攣作用,芹菜素能抑制變形鏈球菌,最低抑菌濃度(MIC)為 1.3 μg/mL[11-12]。木樨草素與芹菜素類似,存在于蔬菜和水果中,如西蘭花、芹菜、歐芹、蘋果等,作為一種清除活性氧(ROS)的抗氧化劑和由于自動氧化而作為一種促氧化劑,木樨草素具有抗炎作用、抗癌作用、抗過敏作用,木樨草素能抑制化膿性鏈球菌,MIC為78 μg/mL[13]。
1.2 黃酮醇
黃酮醇是一類以3-羥基-2-苯基色-4-酮為骨架的黃酮類化合物。這類黃酮類化合物有顯著的抗氧化、抗菌活性。常見于水果和蔬菜中,主要存在于果皮和葉子中,如洋蔥、韭菜、蘋果、葡萄及其制品中。黃酮醇類包括槲皮素、楊梅素、山柰酚等[14]。
槲皮素存在于洋蔥、西蘭花、蘋果、茶和紅酒中,它是一種水溶性植物色素,具有高抗氧化和抗炎活性,對包括金黃色葡萄球菌在內的一些菌株顯示出顯著的抗菌活性,槲皮素對牙齦卟啉單胞菌有顯著的抑制作用,MIC為0.012 5 μg/mL[15-16]。楊梅素常見于漿果、蔬菜以及植物衍生的茶和葡萄酒中。它以游離和糖苷結合的形式天然存在,難溶于水,楊梅素具有廣泛的生物活性,包括抗氧化、抗癌、抗糖尿病和抗炎作用,楊梅素作為抑制金黃色葡萄球菌感染的α溶血素抑制劑,MIC為16 μg/mL[17]山柰酚是一種黃酮醇抗氧化劑,存在于菠菜、羽衣甘藍和西蘭花中。山柰酚能抑制大腸桿菌,MIC為 25 μg/mL[18]。
1.3 黃烷酮
黃烷酮也稱二氫黃酮,黃烷酮類的結構骨架是2,3-二氫-2-苯基色烯-4-酮,具有飽和的C環,作為黃酮類化合物中最大的亞群之一,在植物中廣泛存在,尤其是柑橘類植物[19]。常見的黃烷酮類包括柚皮素、橙皮素、淫羊藿醇、異苦參素及各自的衍生物。黃烷酮類化合物中柚皮素的研究最為廣泛,具有抗菌活性、抗氧化活性、抗炎和抗微生物活性,柚皮素通過其抗氧化特性和清除自由基的能力來進行有效預防和治療肝病,抑制血小板凝結、抗粥樣動脈硬化等疾病,此外,柚皮素還可以抑制脂多糖誘導的巨噬細胞產生促炎細胞因子,減少硝酸鹽和亞硝酸鹽的產生,柚皮素對革蘭氏陽性菌如谷草桿菌、變形鏈球菌等有很好的抗菌活性,MIC為 200 μg/mL[20-21]。
1.4 黃烷醇
黃烷醇類是一類非常復雜的黃酮類化合物,其骨架結構為黃烷-3-醇。黃烷醇主要存在于茶、葡萄酒、可可、蘋果及其制品中,兒茶素被認為是單寧的主要組成成分,是黃烷醇的最重要代表。綠茶和紅茶中含有豐富的兒茶素,不同類型的兒茶素包括兒茶素、表兒茶素、沒食子兒茶素、表沒食子兒茶素、兒茶素沒食子酸酯、表兒茶素沒食子酸酯、沒食子兒茶素沒食子酸酯和表沒食子兒茶素沒食子酸酯[22-23],原花青素是兒茶素的二聚體、低聚體和聚合物,被稱為縮合單寧的一種。兒茶素可以抑制變形鏈球菌,MIC為1 mg/mL,聯合羅伊氏菌素使用可以降至 125 μg/mL[24]。
1.5 異黃酮
異黃酮類的結構骨架是3-苯基色滿骨架。異黃酮被列為最具雌激素的化合物之一,具有抑菌、抗菌、抗病毒和抗氧化活性。異黃酮的主要來源是豆類,包括大豆和大豆制品,紅三葉草和葛根中也含有大量的異黃酮,包括染料木黃酮、黃豆苷元、大豆黃素、芒柄花黃素等[25-26],現階段研究較多的是染料木黃酮和黃豆苷元這2種異黃酮。染料木黃酮被稱為植物雌激素,可以調節類固醇激素受體和多種代謝途徑,使其成為預防和治療常見疾病的重要成分,染料木黃酮能抑制金黃色葡萄球菌,MIC為 32 μg/mL[27]。黃豆苷元是另一種存在于大豆中的異黃酮,是染料木素的非活性類似物,黃豆苷元也能抑制金黃色葡萄球菌,MIC為32 μg/mL[27]。異黃酮結構上與雌激素相似,因此異黃酮可以與雌激素受體結合[28]。
1.6 花青素
花青素是多羥基和多甲氧基衍生物的糖苷。花青素不穩定,通常以糖基化形式存在。花青素通常作為植物色素賦予花朵和果實紅色、紫色和藍色3種顏色。黃酮結構中作為取代基存在的羥基和甲氧基的數量和位置不同形成不同的花青素。迄今為止,已在植物中鑒定出超過650種花青素化合物。天然存在的花青素主要包括飛燕草素、花青素、矮牽牛素、牡丹素、錦葵素、天竺葵素。花青素在水果和蔬菜中含量豐富,例如藍莓、紅甘藍、番茄、紫甘薯、茄子等[29]。酰化花青素能抑制金黃色葡萄球菌和大腸桿菌的活性,降低膜完整性、ATP濃度、pH值以及破壞細胞形態,MIC分別為0.312、4.000 mg/mL[30]。
2 抗菌作用機制
黃酮類化合物的抗菌機制包括2個方面,一方面直接抑制細菌生長,包括改變細胞膜通透性引起細胞質損傷、抑制核酸合成、抑制能量代謝、細胞膜損傷、細胞壁損傷;另一方面是間接抑制細菌生長,藥物作用能抑制相關信號通路(NF-κB、MAPKs等)的表達,抑制促炎細胞因子的釋放,調節T細胞分化以及促炎免疫細胞的激活和浸潤。
2.1 黃酮類化合物的直接抑菌活性
2.1.1 改變細胞膜通透性 細胞膜適當的細胞膜流動性對于細菌維持正常的細胞功能非常重要,例如細胞分裂、擴散和運輸離子,抗菌藥物一般通過靶向細菌的細胞膜和關鍵生物大分子來發揮作用,破壞細胞膜通常是抗菌物質的主要作用方式[31]。黃酮類化合物進入機體后,作用于菌體細胞,通過增加細胞膜的通透性或降低膜的流動性引起細胞質損傷[32]。例如柚皮素可以增加金黃色葡萄球菌細胞膜的流動性,改變細胞膜的通透性和完整性,進而導致細胞內物質、離子和遺傳物質的泄露,抑制細菌活性。當柚皮素的濃度為2.2 mmol/L時,14 h 未見金黃色葡萄球菌生長[33]。此外,3 μg/mL頭孢他啶和3 μg/mL芹菜素聯合使用會損害耐頭孢他啶的陰溝腸桿菌的細胞質膜,導致細胞內成分泄露,發揮抗耐藥菌作用[34]。
2.1.2 抑制能量生成 研究報道,黃酮類化合物可以抑制大腸桿菌ATP合成酶活性,ATP合成酶是一種高度保守的酶,具有F1和F02個區段,在大腸桿菌中,F1由α3β3γδεab2c10組成,而F0由ab2c10組成[35]。ATP的合成和水解發生在F1部分的3個催化位點,而質子的運動則通過包埋在膜上的F0區段進行[36]。大量的多酚化合物已被證明可以結合在不同的多酚結合位點,并抑制ATP合成酶合成,多酚結合位點位于F1區段的α、β、γ亞基的交界處,因此黃酮類化合物的抑制作用是通過結合在三磷酸腺苷合成酶的多酚結合位點上,阻斷γ亞基的順時針或逆時針旋轉[37]。例如,對大腸桿菌ATP合成酶最有效的抑制劑包括黃芩素、表兒茶素,表兒茶素通過降低ATP合成酶的活性來抑制變形鏈球菌的產酸和耐酸特性[35]。
2.1.3 抑制核酸合成 黃烷醇和異黃酮還可以通過抑制DNA拓撲異構酶活性來抑制核酸合成,芹菜素和槲皮素等黃酮類化合物對DNA旋轉酶活性有明顯的抑制作用,槲皮素可抑制細菌旋轉酶的超螺旋活性,并誘導DNA裂解[38]。研究發現,在 0.1 μmol/L 劑量下,金雀異黃素改變了哈維氏弧菌的細胞分裂,隨后抑制了其生長。同樣,參與細菌環狀染色體復制的DNA旋轉酶在抗大腸桿菌時也被鞣花酸抑制[39]。此外沒食子兒茶素沒食子酸酯通過抑制二氫葉酸還原酶的活性來抑制結核分枝桿菌、金黃色葡萄球菌、大腸桿菌的生長,MIC值分別為6.25、0.10、6.00 μg/mL[40]。
2.1.4 抑制細菌毒性 由革蘭氏陽性菌和革蘭氏陰性菌產生透明質酸酶作為重要的毒力因子可直接與宿主組織相互作用。在細菌發病機制中,透明質酸酶介導的透明質酸降解增加了結締組織的滲透性并降低了體液的濃度[41]。研究發現黃酮醇類化合物,如槲皮素可以作為透明質酸裂解酶抑制劑,來抑制無乳鏈球菌[42]。黃酮類化合物的抑制作用隨著黃酮類化合物結構中羥基數量的增加而增加[43]。
此外,黃酮類化合物,尤其是兒茶素由于抗氧化特性可以用于中和金黃色葡萄球菌細菌毒性因子,MIC為78~156 μg/mL[44]。同樣,黃芩苷和染料木黃酮也可以用于中和金黃色葡萄球菌的外毒素,MIC分別為300、128 μg/mL[45-47]。沒食子兒茶素沒食子酸酯可通過與細胞膜的結合來抑制放線桿菌聚合酶釋放白細胞毒素,MIC為 10 μg/mL[48]。
2.1.5 對細菌細胞質膜的損傷 細胞質膜負責滲透壓調節、呼吸和轉運過程、肽聚糖的生物合成以及脂質的合成。維持膜完整性是先決條件,其被破壞可直接或間接導致代謝功能障礙并最終導致細菌死亡[49]。細菌細胞壁的主要成分是肽聚糖,黃酮類化合物可以使細菌細胞壁中肽聚糖以及蛋白質的合成受阻,最終導致細菌細胞膜和細胞壁的損傷。沒食子兒茶素沒食子酸酯通過產生ROS,導致弧菌細胞膜通透性改變和膜損傷,MIC為256 μg/mL[50]。另外,FAS-Ⅱ通路在革蘭氏陰性菌細胞壁和細胞膜的合成中是必不可少的,分枝桿菌的細胞壁中含有一種特有的脂肪酸為分枝桿菌酸,是由哺乳動物FAS-Ⅰ型和細菌FAS-Ⅱ型合成,使該脂肪酸具有高抗性,因此可以通過抑制這2種脂肪酸合成酶活性來尋找新的藥物。據報道,槲皮素、山柰酚、桑黃素、楊梅素、黃芩素、木樨草素和兒茶素可以抑制哺乳動物型FAS-Ⅰ,異甘草素可用于抑制牛分枝桿菌細菌型FAS-Ⅱ的活性[51]。
2.2 黃酮類化合物的間接抑菌活性——通過調節宿主的免疫反應發揮抗菌作用
2.2.1 黃酮類化合物治療幽門螺旋桿菌感染引起的胃黏膜病變 幽門螺旋桿菌是引起胃腸病變的主要原因之一,幽門螺旋桿菌感染可引起慢性胃炎、胃潰瘍和十二指腸潰瘍,更嚴重的可引起胃癌。幽門螺旋桿菌感染宿主后,通過Toll樣受體(TLR4)激活下游信號通路,包括NF-κB通路,機體IL-6、IL-12、TNF-α和γ干擾素(IFN-γ)等促炎細胞因子的水平顯著增加,導致胃黏膜細胞和胃組織病變增加,胃黏膜屏障破壞,引起中性粒細胞浸潤和細胞骨架破壞[52]。
研究發現,荷葉黃酮對幽門螺旋桿菌感染引起的胃黏膜損傷有抑制作用,荷葉黃酮含有山柰酚、槲皮素等化合物,但黃酮類化合物無法直接對幽門螺旋桿菌起抑制作用[53]。化合物作用于胃組織后,能夠抑制NF-κB通路,下調胃組織中IL-6、IL-12、TNF-α和IFN-γ細胞因子的水平,降低炎癥反應,修復胃黏膜并改善黏膜屏障功能[54]。
除此以外,黃酮類化合物如芹菜素能有效抵抗幽門螺旋桿菌感染,用芹菜素處理(9.6~74.0 μmol/L)顯著增加了IκBα的表達,進而抑制NF-κB的激活和炎性因子COX-2、ROS、IL-6和IL-8的水平,能有效治療幽門螺旋桿菌感染引起的胃上皮細胞炎癥[55]。
2.2.2 黃酮類化合物治療大腸桿菌引起的結腸炎 在結腸炎感染大鼠模型中,大鼠感染致病性大腸桿菌后,受腸道微生物群的影響,NF-κB和MAPKs信號通路被激活,還原型輔酶Ⅱ表達增加,IκB磷酸化水平升高和NF-κB p65活化,從而導致腸屏障功能破壞。此外T細胞被分化為促炎Th1和Th17效應細胞,并增加了感染期間免疫細胞被激活和浸潤的程度[56]。大鼠經34.6 mg/kg木樨草素懸液灌胃后,可以抑制腸道內各種病原物的生長并增加各種有益菌群例如乳酸桿菌等來保護腸屏障的完整性,調節T細胞抑制Th1和Th17效應細胞分化,產生抗炎IL-10,并抑制NK-κB和MAPKs信號通路,下調還原型輔酶Ⅱ的表達,減少氧化應激,抑制IκB磷酸化和NF-κB p65與DNA的結合活性[57]。
2.2.3 黃酮類化合物治療支原體引起的肺炎 在肺炎支原體感染小鼠模型中,支原體感染后,激活了肺組織中Wnt/β-連環蛋白(β-catenin)和 NF-κB 通路,β-catenin和NF-κB p65水平升高,進而促進T細胞、成纖維細胞和巨噬細胞中促炎因子IL-6、IL-1β和TNF-α的釋放,并抑制抗炎因子IL-10的水平。而使用黃酮類化合物桑辛素后,Wnt/β-catenin 和 NF-κB 信號通路被明顯抑制,β-catenin 和NF-κB p65水平降低,促炎因子IL-6、IL-1β和TNF-α降至正常水平,同時抑炎因子IL-10的水平顯著降低[59],能有效抑制肺炎支原體感染引起的肺炎。
雞毒支原體(MG)通過激活Toll樣受體(TLR)信號轉導NF-κB通路和促炎細胞因子的表達,引起嚴重的肺部炎癥和細胞損傷。黃芩苷是從黃芩中提取的黃酮類化合物,具有抗菌和抗炎活性,能回調因MG感染引起的雞成纖維細胞活性下降,抑制TLR6、髓樣分化因子88(MyD88)和NF-κB的mRNA表達水平,抑制NF-κB p65核轉位,抑制MG感染后雞肺部IL-1β、IL-6和TNF-α的表達。結果表明,黃芩苷可以通過Toll樣受體6(TLR6)介導的NF-κB通路抑制雞毒支原體誘導的肺部炎癥[60-61]。
2.2.4 黃酮類化合物治療由肺炎鏈球菌引起的流感病毒并發癥 甲型流感病毒(IAV)感染通常與由細菌病原體引起的繼發癥有關,最常見的是肺炎鏈球菌,IAV和肺炎鏈球菌存在一種由神經氨酸酶(NA)介導的致命協同機制,NA存在于2種病原體上,病毒NA促進病毒繁殖和傳播,肺炎鏈球菌生成NA并接近肺上皮細胞上的受體,同時病毒NA為肺炎鏈球菌定殖和生長提供附著受體和營養物質。因此,需要尋找能夠對病毒和細菌NA起雙重抑制作用的藥物。研究發現中草藥桑白皮作為抗感染藥物,其異戊二烯黃酮衍生物:桑根酮G和桑根酮醇A,能抑制流感病毒和肺炎鏈球菌的NA,進而抑制肺炎鏈球菌的生長和生物膜的形成[62]。此外,小鼠體內感染肺炎鏈球菌模型中,7-羥基黃烷酮(1 mg/小鼠)能顯著減少肺中存活的肺炎鏈球菌數量[63]。
2.2.5 黃酮類化合物治療肺炎克雷伯菌感染 肺炎克雷伯菌是一種革蘭氏陰性菌,兼性厭氧菌,感染機體后可引起急性損傷,導致包括肺炎和敗血癥在內的嚴重感染。肺炎克雷伯菌感染后肺組織出現水腫、炎性細胞大量聚集和肺泡出血,NF-κB和MAPK通路激活,p38、ERK、JNK以及IκB和NF-κB p65的磷酸化水平升高,炎性細胞因子TNF-α、IL-6、IL-1β和PGE2水平顯著升高,氧化應激指標髓過氧化物酶(MPO)和丙二醛(MDA)水平顯著增加,超氧物歧化酶(SOD)和谷胱甘肽(GSH)水平下降[64]。研究發現從百蕊草中分離的紫云英苷(kaempferol-3-O-glucoside,KA)可以通過抑制一氧化氮合成酶(iNOS)合成來發揮抗炎活性,在感染肺炎克雷伯菌前1 h按50、100、200 μg/kg的KA劑量腹腔注射小鼠,與對照組相比,KA處理小鼠中性粒細胞和巨噬細胞數量減少,NF-κB 和MAPK通路被抑制,隨著KA劑量的升高,p38、ERK、IκB和NF-κB p65的磷酸化逐漸被抑制,炎性細胞因子的水平逐漸降低。并且氧化應激被抑制,MPO和MDA水平降低,SOD、GSH水平升高[65]。
3 黃酮類化合物應用前景
細菌抗生素耐藥性對全世界人類和動物的健康構成了嚴重的威脅,植物中含有許多天然化合物,可作為抗菌藥物和替代藥物,如黃酮類、生物堿等,許多研究將天然化合物作為抗生素的替代物,這些化合物易分離且對機體幾乎無害[66]。黃酮類化合物通過不同的機制發揮抑菌作用。可以直接抑制細菌活性,以及聯合抗生素來抑制細菌活性,能夠增加耐藥菌株的敏感性,并且可以減少抗生素的劑量,還可以通過調控宿主細胞的免疫反應間接抑制細菌誘導的炎癥。這將為今后探索替代抗生素藥物或者減少抗生素劑量提供新的思路。
然而,動物機體對黃酮類化合物的生物利用率較低,大部分被機體吸收后迅速被代謝排出體外,需要在維持活性不變的基礎上提高黃酮類化合物的水溶性,并進行結構修飾,使其具有針對性和高效性[67]。因此,有必要提高并優化黃酮類化合物的提取技術,形成一套完整高效的提取工藝流程,為今后新藥物的研發奠定基礎。
猜你喜歡
四川蠶業(2021年2期)2021-03-09 03:15:32
四川蠶業(2021年3期)2021-02-12 02:38:46
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
哈爾濱醫藥(2016年1期)2017-01-15 13:43:16
天然產物研究與開發(2016年11期)2016-06-15 20:29:17
湖南師范大學自然科學學報(2015年1期)2015-02-27 14:50:04
安徽醫藥(2014年12期)2014-03-20 13:15:15
