大黃魚養(yǎng)殖海域沉積物中抗生素抗性基因分布特征及其影響因素
2023-03-06 08:02:18黃薇劉洋饒秋華羅欽王為剛宋永康羅土炎
農業(yè)環(huán)境科學學報 2023年1期
關鍵詞:環(huán)境
黃薇,劉洋,饒秋華,羅欽,王為剛,宋永康*,羅土炎*
(1.福建省農業(yè)科學院農業(yè)質量標準與檢測技術研究所/福建省農產品質量安全重點實驗室,福州 350003;2.福建師范大學生命科學學院,福州 350108;3.連江縣水產技術推廣站,福州 350500)
抗生素在醫(yī)療、畜禽養(yǎng)殖和水產養(yǎng)殖中都發(fā)揮著十分重要的作用。在水產養(yǎng)殖過程中,抗生素被用于防治各種細菌性疾病,同時兼具促進水生動物生長等功效[1]。為了提高養(yǎng)殖產量、降低養(yǎng)殖成本,抗生素在水產養(yǎng)殖過程中被長期過度使用,這導致了水產養(yǎng)殖環(huán)境和養(yǎng)殖動物體內耐藥菌和抗生素抗性基因(Antibiotic resistance genes,ARGs)[2]的產生。ARGs于2006 年由Pruden 等[3]首次確認為新型的環(huán)境污染物,由于可以通過水平基因轉移在環(huán)境和生物體內長久持續(xù)傳播,ARGs 的危害性相對于抗生素和耐藥菌更強,因此其被世界衛(wèi)生組織列為21 世紀人類醫(yī)療健康面臨的三大威脅之一[4]。
水產養(yǎng)殖環(huán)境被證明是ARGs 的潛在儲藏庫,ARGs 在水產養(yǎng)殖環(huán)境(水體、沉積物)及水產養(yǎng)殖對象中普遍存在[5-7]。Hedberg 等[8]證實沿海水域的ARGs 主要來源于水產養(yǎng)殖。水產養(yǎng)殖環(huán)境中的ARGs可以通過食物鏈直接與人類腸道菌群進行水平基因轉移傳播[9]。Yang 等[10]的研究顯示,海水養(yǎng)殖沉積物中的ARGs 序列與人體致病菌中的ARGs 序列高度相似。因此,隨著全球水產養(yǎng)殖業(yè)的快速發(fā)展,水產養(yǎng)殖環(huán)境中的ARGs 引起了更多的關注,尤其是沉積物。與水不同,沉積物中的ARGs 具有更穩(wěn)定、更持久的特點,其ARGs 賦存量顯著大于水,被認為是ARGs 的“匯”[11]。Yuan 等[12]認為隨著時間的推移,水中的ARGs 會在沉積物中不斷積累,沉積物很可能增強了抗生素抗性的傳播。
不同養(yǎng)殖區(qū)域、不同養(yǎng)殖對象以及不同養(yǎng)殖模式造成的ARGs賦存情況顯著不同[11]。大黃魚是我國養(yǎng)殖規(guī)模最大的海水魚,也是海區(qū)網箱養(yǎng)殖最主要的品種[13],然而對大黃魚養(yǎng)殖海域沉積物ARGs 賦存特征的研究尚未見報道。目前,ARGs 的檢測方法主要以PCR 技術為主,雖然很多研究人員已經利用PCR 方法對水產養(yǎng)殖環(huán)境中的ARGs 進行了定量研究,且取得了一定成果,但對研究ARGs 的種類有限制,很難得到環(huán)境中微生物抗生素抗性基因組的全面且詳細的信息[14]。近年來,隨著高通量測序和功能基因組篩選技術的發(fā)展,宏基因組測序分析技術不再局限于PCR引物設計,可以為研究環(huán)境抗生素抗性基因組提供ARGs 的廣譜特征,從而全面客觀地反映環(huán)境中ARGs 的多樣性。因此,本文以大黃魚養(yǎng)殖海域沉積物為研究對象,利用宏基因組測序分析技術調查大黃魚養(yǎng)殖海域內ARGs 的污染狀況,并探討ARGs 與環(huán)境因子以及微生物群落結構之間的相關性,揭示ARGs 在水產養(yǎng)殖環(huán)境中的擴散和傳播規(guī)律,以期為評價大黃魚養(yǎng)殖區(qū)域ARGs 的生態(tài)風險和污染控制提供參考依據(jù)。
1 材料與方法
1.1 采樣點及樣品采集
2022年1月選取6個大黃魚養(yǎng)殖密集區(qū)采集養(yǎng)殖魚排下的沉積物樣品,具體采樣點分布如圖1所示,采樣點信息見表1。每個采樣點選取3個不同的網箱(分別用A、B、C 表示),選取的網箱之間至少間隔一個網箱,在選取的網箱框架踏板上使用抓斗采泥器采集沉積物樣品,樣品采集后立即放入無菌的聚乙烯塑料袋中,放到低溫(冰浴)冷藏箱中運至實驗室,置于-80 ℃冰箱保存待測。
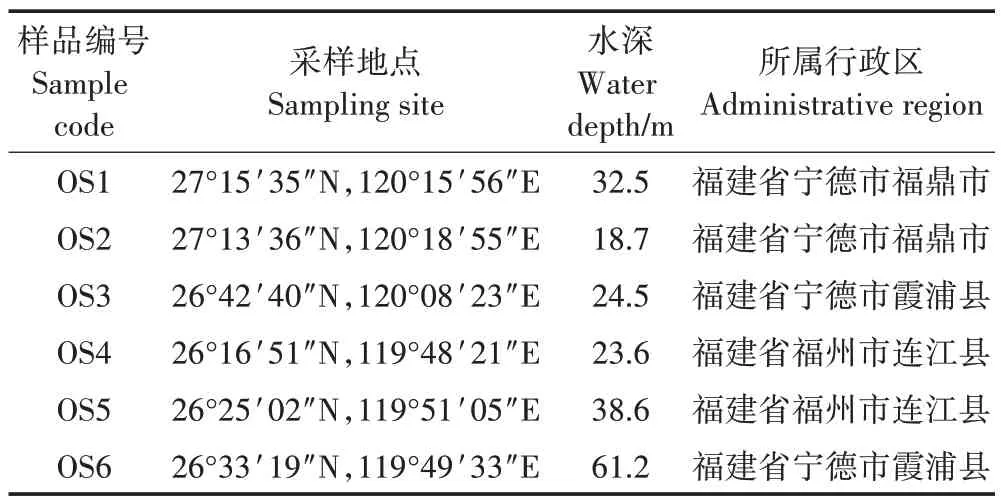
表1 樣本的基本信息Table 1 Basic information of the samples

圖1 采樣點分布示意圖Figure 1 Location of sampling sites
1.2 沉積物理化因子測定
樣品pH和電導率(含鹽量)通過出海采樣實時檢測得到,pH 使用便攜式pH 計(METTLER TOLEDO,瑞士)獲取,電導率通過便攜式電導率儀(METTLER TOLEDO,瑞士)測定。有機碳(TOC)采用重鉻酸鉀氧化-還原容量法測定,總氮(TN)采用凱氏滴定法測定,總磷(TP)采用分光光度法測定。
1.3 沉積物微生物DNA提取
使用PowerSoil DNA Isolation Kit 試劑盒(MOBIO,Carlsbad,CA,美國),參照試劑盒說明書進行沉積物總基因組DNA 的提取,使用Qubit 2.0 Fluorometer(Invitrogen,Carlsbad,CA,美國)檢測DNA 樣品的濃度,然后將DNA 樣品放置于-80 ℃下保存?zhèn)溆谩?6S rRNA 基因高通量測序樣品為每個采樣點的3 個平行沉積物樣品分別提取DNA,宏基因高通量測序樣品為每個采樣點的3 個平行沉積物樣品等質量混合后提取DNA,最后共獲得18 個用于16S rRNA 基因高通量測序的基因組DNA 樣品和6 個宏基因組高通量測序的基因組DNA樣品。
1.4 16S rRNA基因高通量測序及生物信息學分析
16S rRNA基因高通量測序DNA樣品送上海美吉生物醫(yī)藥科技有限公司,于Illumina MiSeq 平臺進行測序,16S rRNA 基因高通量測序擴增16S rRNA 基因V3~V4 可變區(qū),通用引物為338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′ -GGACTACHVGGGTWTCTAAT-3′)。Miseq 測序得到的原始序列首先根據(jù)overlap 關系進行拼接,之后裁切序列中的正反向引物,同時對序列質量進行質控和過濾,區(qū)分樣本后按97%序列相似度進行OTU 聚類分析劃分分類操作單元(OTUs),再運用Ribosomal Database Project(RDP)classifier進行物種注釋,得到每條序列從門到屬各個水平的分類信息,基于分類學信息,統(tǒng)計每個生物樣本的多樣性指數(shù)。
1.5 宏基因組高通量測序及生物信息學分析
宏基因組高通量測序DNA 樣品送上海美吉生物醫(yī)藥科技有限公司,于Illumina Hiseq 2500 測序平臺進行測序。Hiseq 測序得到的原始序列首先進行質控,剪切接頭序列、低質量序列及含N堿基序列,然后利用Megahit 與Newbler 拼接軟件對質控數(shù)據(jù)進行拼接組裝,再使用MetaGene 軟件對拼接結果中的contigs進行ORF預測。預測出的基因序列按相似度95%進行聚類,每類取最長的基因作為代表序列,構建非冗余基因集;將數(shù)據(jù)上傳CARD數(shù)據(jù)庫進行比對,獲得基因對應的ARGs功能注釋信息,包括基因名稱、抗生素抗性類型、耐藥機制以及基因如何與抗生素藥物有關的描述等;ARGs 的相對豐度使用百萬分率(10-6)表示,即在1 000 000 個質控序列中有一個被標注為ARG的序列。
1.6 統(tǒng)計分析
采用Excel 軟件計算數(shù)據(jù)的平均值和標準偏差。采用SPSS軟件進行方差分析和相關性分析,方差分析采用LSD檢驗,相關性分析選擇spearman算法。應用R 語言vegan、gplots、venn diagram 和pheatmap 等軟件包繪制PCoA圖、柱狀圖、韋恩圖、熱圖和相關性熱圖。
2 結果與分析
2.1 大黃魚養(yǎng)殖海域沉積物中抗生素抗性基因分布特征
本研究從6 個沉積物底泥樣本中共獲得了80 604 948~94 023 130條(平均89 055 467條)宏基因組高通量測序質量過濾序列,其中275 976~557 910條(0.32%~0.36%)序列被注釋為CARD 抗性基因序列(BLASTp,e-value≤10-5)。圖2 為豐度前50 的ARGs,OS1~OS6 位點豐度最高的ARG 均為macB(254.31 × 10-6~436.85 × 10-6),其次均為tetA(58)(131.64×10-6~241.69×10-6)、evgS(87.00×10-6~193.42×10-6)和bcrA(76.75×10-6~128.89×10-6),上述優(yōu)勢ARGs分別占ARGs序列總數(shù)的15.44%~16.48%,表明不同大黃魚養(yǎng)殖海域沉積物中的優(yōu)勢抗生素抗性基因種類基本一致。
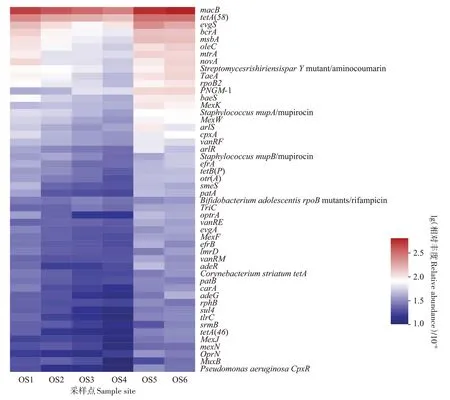
圖2 大黃魚養(yǎng)殖海域沉積物中ARGs賦存特征Figure 2 Occurrence characteristics of ARGs in the different sediment samples from large yellow croaker culture area
根據(jù)統(tǒng)計結果,在大黃魚養(yǎng)殖海域沉積物樣品中共識別出38 種抗生素抗性類型,781 個ARGs 亞型。圖3a為不同抗生素抗性類型ARGs的相對豐度,在大黃魚養(yǎng)殖海域沉積物中,ARGs 的主要抗生素抗性類型為四環(huán)素類(Tetracyclines,1 049.03×10-6~2 061.39×10-6)、大環(huán)內酯類(Macrolides,999.59×10-6~1 946.88×10-6)和氟喹諾酮類(Fluoroquinolones,660.02×10-6~1 292.33×10-6)。圖3b 為不同耐藥機制的ARGs 占ARGs 序列總數(shù)的相對百分比情況,從結果中可以看出,不同大黃魚養(yǎng)殖海域沉積物樣品中ARGs 引起細菌耐藥的機制基本一致,其中外排泵(Antibiotic efflux)是主要的耐藥機制,占ARGs 序列總數(shù)的62.63%~63.84%,其次為抗生素靶點改變(Antibiotic target alteration),占ARGs序列總數(shù)的18.76%~20.03%。
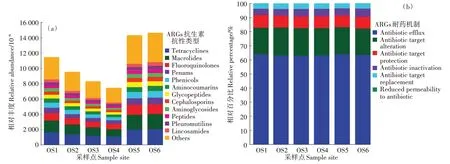
圖3 大黃魚養(yǎng)殖海域沉積物中ARGs的抗生素抗性類型和耐藥機制Figure 3 Antibiotic resistance types and resistance mechanisms of ARGs in different sediment samples from large yellow croaker culture area
2.2 大黃魚養(yǎng)殖海域沉積物ARGs與環(huán)境因子之間的關系
2.2.1 大黃魚養(yǎng)殖海域沉積物中環(huán)境因子分布特征
大黃魚養(yǎng)殖海域沉積物中5 種環(huán)境因子的檢測結果見表2。6 個位點的大黃魚養(yǎng)殖海域沉積物樣品的pH 和電導率相似,但TOC、TN 和TP 存在顯著差異(P<0.05),TOC 范圍為0.48%(OS1)~0.59%(OS4),TN含量范圍為398(OS1)~563 mg·kg-1(OS6),TP 含量范圍為546(OS5)~777 mg·kg-1(OS1)。根據(jù)我國《海洋沉積物質量》(GB 18668—2002),6 個位點的TOC 均超過第三類海洋沉積物質量標準,根據(jù)加拿大安大略省環(huán)境和能源部1992 年制定的沉積物標準,OS6中TN 屬輕度污染,OS1、OS2、OS3 和OS6 中TP 為輕度污染。
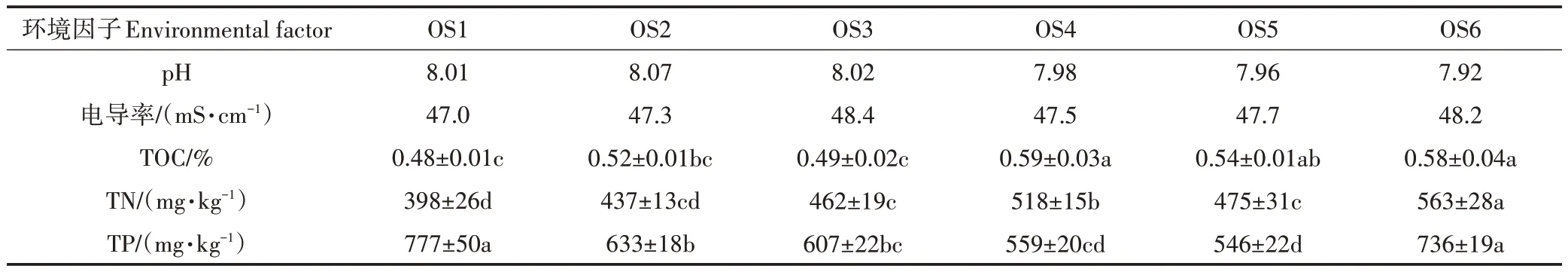
表2 大黃魚養(yǎng)殖海域沉積物中環(huán)境因子分布特征(平均值±標準誤,n=3)Table 2 Distribution characteristics of environment factors in different sediment samples from large yellow croaker culture area(Mean±SEM,n=3)
2.2.2 沉積物中環(huán)境因子與ARGs之間的相關性
為探究大黃魚養(yǎng)殖海域沉積物中環(huán)境因子對ARGs 分布特征的影響,使用相關性分析解析6 個位點大黃魚養(yǎng)殖海域沉積物中的5 種環(huán)境因子與豐度前20 的ARGs 之間的關系,結果見圖4。如圖4 所示,大黃魚養(yǎng)殖海域沉積物中,pH 與豐度前20 的ARGs均存在負相關關系,TN 與豐度前20 的ARGs 均存在正相關關系,但除PNGM-1 與電導率和TN 呈顯著正相關(P<0.05)外,其余ARGs 與所檢環(huán)境因子之間的相關性均不顯著(P>0.05)。
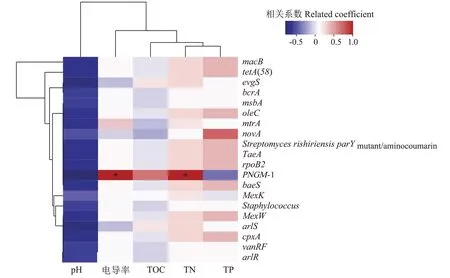
圖4 大黃魚養(yǎng)殖海域沉積物中ARGs和環(huán)境因子的關系Figure 4 Correlation between ARGs and environmental factors in the sediment of large yellow croaker culture area
2.3 大黃魚養(yǎng)殖海域沉積物ARGs與微生物群落的關系
2.3.1 大黃魚養(yǎng)殖海域沉積物中細菌群落結構
18 個大黃魚養(yǎng)殖海域沉積物樣品Illumina MiSeq測序數(shù)據(jù)統(tǒng)計分析結果見表3。從表3 可知,18 個沉積物樣品獲得的16S rRNA 序列數(shù)為56 007~66 002條,OTUs 為2 066~3 451,每個樣品的OUT 覆蓋率均大于96%,表明Illumina MiSeq 測序獲取的數(shù)據(jù)量能夠很好地反映特定樣品的細菌多樣性情況。Alpha多樣性分析結果表明,盡管OS2、OS3和OS4沉積物樣品的覆蓋率要顯著低于OS1、OS5 和OS6(P<0.05),但是OS2、OS3 和OS4 的OTUs、ACE 指數(shù)和Chao 指數(shù)要顯著高于OS1、OS5 和OS6(P<0.05),表明OS2、OS3 和OS4 樣品中的物種數(shù)要高于OS1、OS5 和OS6。此外,除OS1 的Shannon 指數(shù)外,各樣本的Shannon 指數(shù)和Simpson 指數(shù)的差異不顯著(P>0.05),說明各樣品間的物種豐度和多樣性差異不顯著。Beta 多樣性分析(PCoA 分析,圖5)結果表明,除OS3 和OS4 微生物組成結構相似外,不同采樣點的微生物組成結構存在明顯差異。
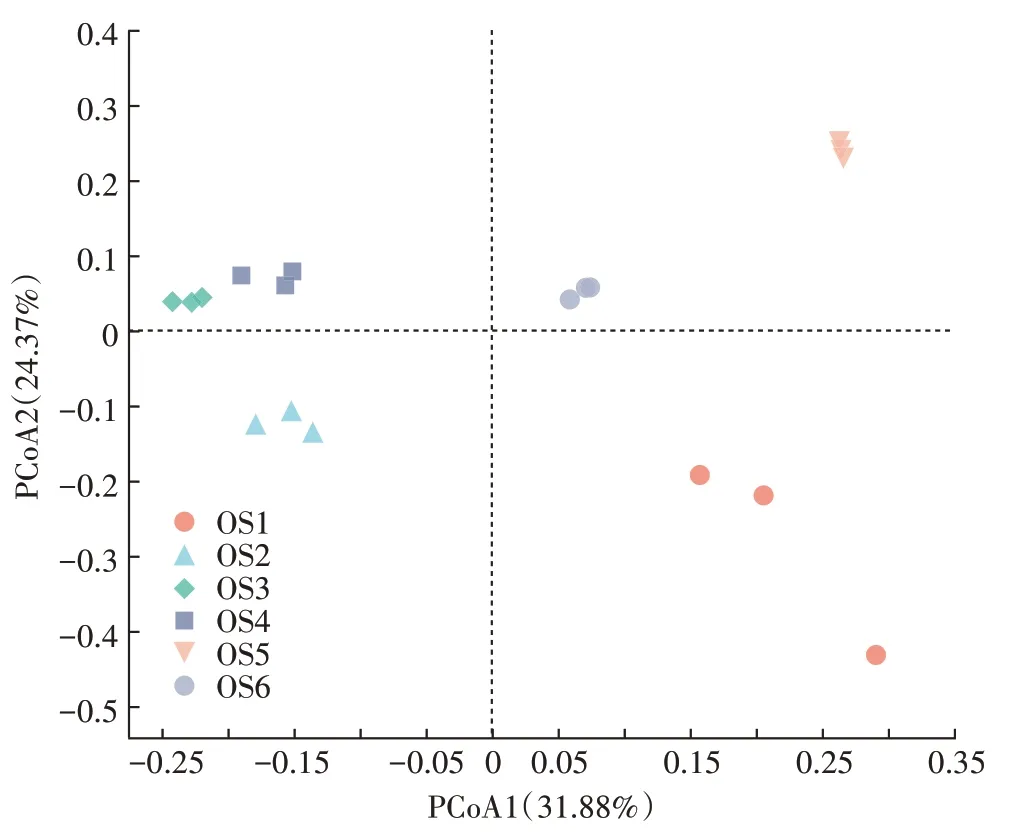
圖5 大黃魚養(yǎng)殖海域沉積物樣品細菌群落結構PCoA分析Figure 5 Principal coordinates analysis(PCoA)of bacterial community compositions in the different sediment samples from large yellow croaker culture area
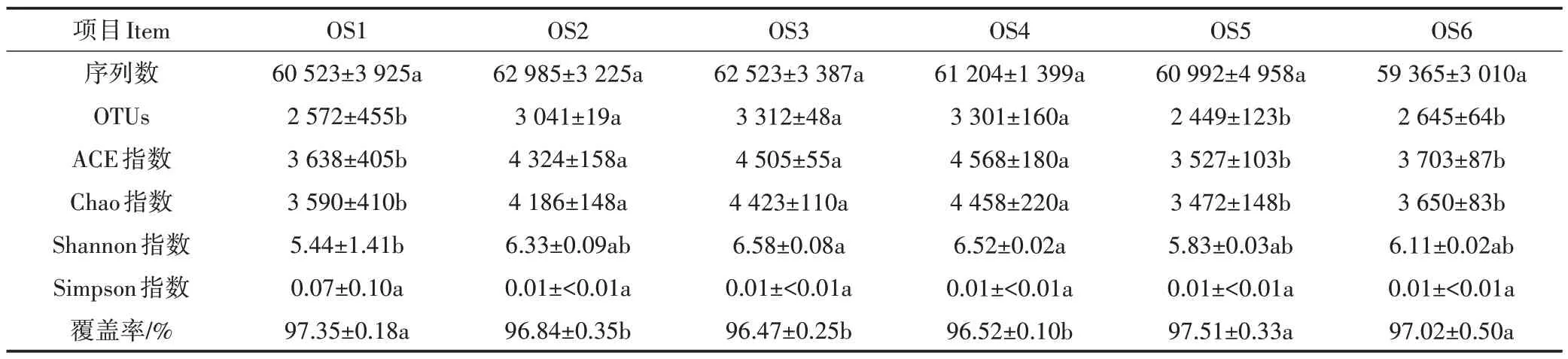
表3 Illumina MiSeq測序數(shù)據(jù)統(tǒng)計分析(平均值±標準誤,n=3)Table 3 Statistical indexes calculated based on the Illumina MiSeq sequencing data(Mean±SEM,n=3)
通過構建韋恩圖分析了不同大黃魚養(yǎng)殖海域沉積物樣品中所共有的和特有的OTU 數(shù)目(圖6)。如圖6 所示,18 個大黃魚養(yǎng)殖海域沉積物樣品共有1 059 個OTUs,統(tǒng)計分析表明,共有的OTUs 序列分別占樣品序列總數(shù)的13.68%~22.85%,特有OTUs 序列分別占樣品序列總數(shù)的0.26%~1.39%。從物種注釋結果來看,本研究共得到細菌59 個門(圖7a),981 個屬(圖7b)。對注釋結果進行統(tǒng)計分析,從門水平上看,除OS3 外,其余大黃魚養(yǎng)殖海域沉積物中豐度最高的細菌門均為Proteobacteria,其占樣品序列總數(shù)的21.36%±0.86%~44.19%±22.59%,OS3 豐度最高的細菌門為Desulfobacterota;其次,OS1、OS2 和OS6 為Bacteroidota(13.45% ± 6.97%、14.72% ± 1.57% 和15.82%±0.38%),OS5 為Acidobacteriota(11.22%±1.38%),OS4為Desulfobacterota(16.13%±0.87%),OS3為Proteobacteria(20.09%±0.67%);對樣品中前20 的門的豐度進行差異顯著性分析,結果顯示這些門類的細菌在不同大黃魚養(yǎng)殖海域沉積物中均存在顯著差異(P<0.05)。從屬水平上看,OS1 豐度最高的屬為Pseudoalteromonas,OS2、OS4 和OS5 豐度最高的屬為Woeseia,OS3豐度最高的屬g_no rank_o_Sva1033,OS6豐度最高的屬為g_no rank_f_unclassified。同時對樣品中前20 的屬的豐度進行差異顯著性分析,結果顯示 除g_no rank_o_Actinomarinales、Pseudoalteromonas不顯著(P>0.05)外,其他屬類細菌在不同大黃魚養(yǎng)殖海域沉積物中均存在顯著差異(P<0.05)。以上說明不同點位的大黃魚養(yǎng)殖海域沉積物的細菌豐度組成存在明顯差異。
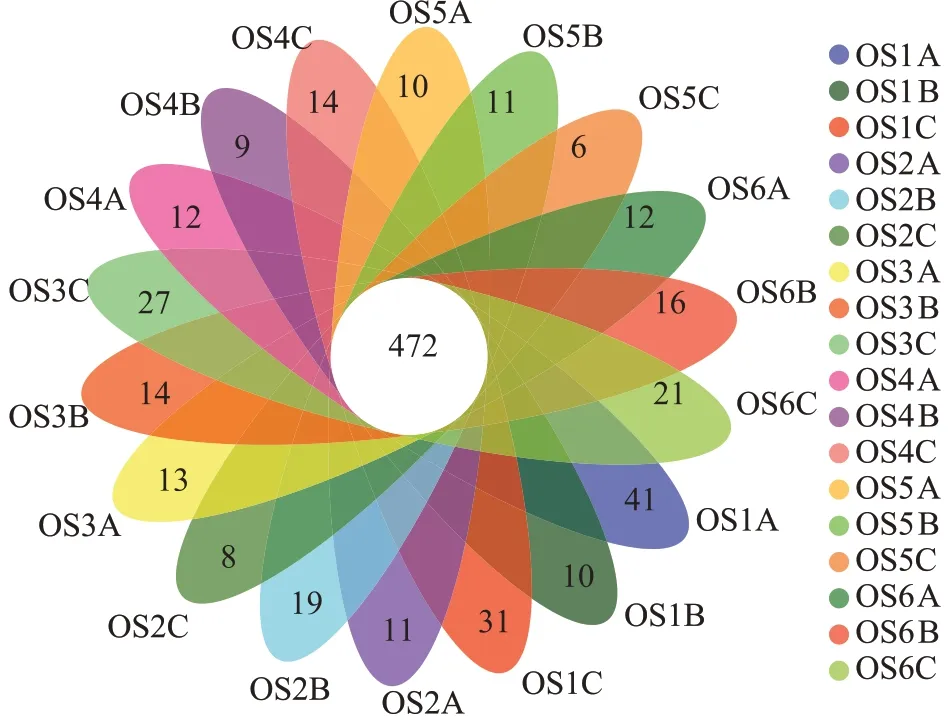
圖6 不同大黃魚養(yǎng)殖海域沉積物中共有和特有的OTUsFigure 6 Venn diagram displaying the number of shared and unique OTUs in the different sediment samples from large yellow croaker culture area
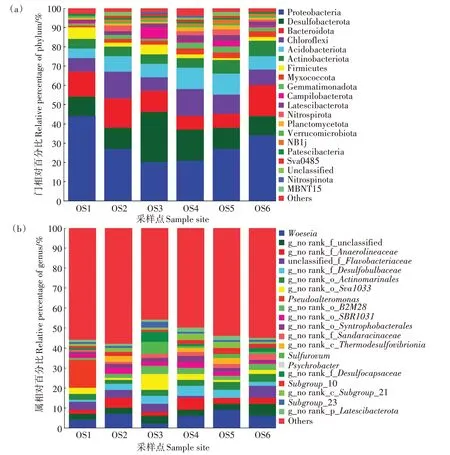
圖7 門水平下和屬水平下不同大黃魚養(yǎng)殖海域沉積物中細菌群落結構組成Figure 7 Bacterial community compositions in the different sediment samples from large yellow croaker culture area at the phylum level and at the genus level
2.3.2 沉積物中細菌群落對ARGs分布特征的影響
在大黃魚養(yǎng)殖海域沉積物中,豐度前20 的細菌屬與豐度前20 的ARGs 之間的相關關系如圖8 所示。由圖8可知,除PNGM-1外,大部分ARGs之間呈顯著或極顯著正相關(P<0.05 或P<0.01)。從微生物群落結構與ARGs之間的關系來看,豐度前20的微生物菌屬中有8 種與ARGs 呈顯著或極顯著相關。其中,g_no rank_c_Subgroup_21 與豐度前20 種的15 種ARGs 呈顯著或極顯著負相關(P<0.05 或P<0.01),涉及8 種抗生素抗性類型和2 種耐藥機制;unclassified_f_Flavobacteriaceae與13種ARGs呈顯著或極顯著正相關(P<0.05 或P<0.01),涉及6 種抗生素抗性類型和2 種耐性機制;g_no rank_c_Thermodesulfovibrionia與8 種ARGs 呈顯著負相關(P<0.05),涉及5 種抗生素抗性類型和1 種耐藥機制;g_no rank_f_Desulfocapsaceae與6 種ARGs 呈顯著或極顯著正相關(P<0.05或P<0.01),涉及2種抗生素抗性類型和2種耐藥機制;g_no rank_f_unclassified 與4 種ARGs 呈顯著正相關(P<0.05),涉及3 種抗生素抗性類型和3 種耐藥機制;g_no rank_f_Anaerolineaceae與4種ARGs呈顯著負相關(P<0.05),涉及4 種抗生素抗性類型和1 種耐藥機制;g_no rank_o_Syntrophobacterales與2 種ARGs呈顯著或極顯著負相關(P<0.05 或P<0.01),涉及2 種抗生素抗性類型和1 種耐藥機制;Woeseia與2 種ARGs 呈顯著負相關(P<0.05),涉及1 種抗生素抗性類型和1種耐藥機制。從ARGs與微生物群落之間的關系來看,豐度前20 的ARGs 均與豐度前20 的菌屬有不同程度的顯著相關性,其中,macB、tetA(58)、evgS、bcrA、msbA、oleC、mtrA、novA、TaeA、rpoB2、PNGM-1、baeS、Staphylococcus mupA/mupirocin、MexW、vanRF和arlR分別與1種菌屬呈顯著或極顯著正相關(P<0.05 或P<0.01),分別與1、3、3、3、1、3、3、4、1、1、0、2、1、1、1 種和0 種菌屬呈顯著或極顯著負相關(P<0.05 或P<0.01);Streptimyoes rishiriensis parYmutant/aminocoimarin、MexK、arlS和cpxA與2 種菌屬呈顯著或極顯著正相關(P<0.05 或P<0.01),與0、0、1 種和2種菌屬呈顯著或極顯著負相關(P<0.05或P<0.01)。
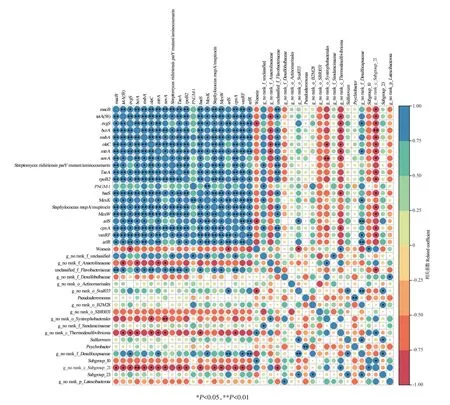
圖8 ARGs與微生物菌屬相關性分析Figure 8 Correlation analysis between ARGs and microbial genus
3 討論
本研究利用宏基因組高通量測序技術首次對大黃魚養(yǎng)殖海域沉積物中ARGs 賦存特征進行了分析。宏基因組高通量測序方法可以使檢出的ARGs 更加全面[9,15]。從測序分析結果可知,本研究在6 個大黃魚養(yǎng)殖海域沉積物中共檢測出781 個ARGs 亞型,其中有許多未曾在大黃魚養(yǎng)殖環(huán)境中報道,如fusH、QnrS7等。從ARGs 注釋結果來看,大黃魚養(yǎng)殖海域沉積物中ARGs 最主要的抗生素抗性類型為四環(huán)素類、大環(huán)內酯類和氟喹諾酮類,而在多數(shù)以PCR 檢測方法研究抗性基因的報道中,磺胺類抗性基因被認為是我國海水養(yǎng)殖場中常見的抗性基因,其豐度占環(huán)境總ARGs 的比例較高[5,16-17],存在這種差異的原因可能與PCR 檢測目標ARGs種類的局限性造成ARGs統(tǒng)計結果產生差異有關。在本研究中,大黃魚養(yǎng)殖海域沉積物中優(yōu)勢ARGs 的種類與已報道的珠江三角洲[18]、海南東寨港[19]、東海近海(杭州灣、象山灣和臺州灣)[20]等沉積物中的ARGs賦存特征存在明顯的差異。ARGs 的產生和環(huán)境殘留機制較為復雜,如抗生素是ARGs 發(fā)生和傳播的主要應激源,但抗生素與其相應的ARGs 也并非總是同時出現(xiàn),使用單一類型的抗生素也可能出現(xiàn)復雜的ARGs 污染[21-22]。近年來的研究表明,水產養(yǎng)殖環(huán)境ARGs 與養(yǎng)殖動物腸道菌群特征、環(huán)境因子、養(yǎng)殖藥物和飼料以及養(yǎng)殖模式等諸多因素均有密切聯(lián)系[23-25]。目前,有關大黃魚養(yǎng)殖環(huán)境中ARGs 污染情況的研究尚處在初期階段,ARGs 污染特征的時空分布特征以及影響因子還有待進一步深入研究。同時在本研究中,大黃魚養(yǎng)殖海域沉積物中優(yōu)勢ARGs 的豐度為76.75×10-6~436.85×10-6,這比相同算法的對蝦養(yǎng)殖沉積物的豐度大得多[26]。明確ARGs耐藥機制對防止或延緩耐藥性的產生具有重要意義[27]。本研究結果顯示大黃魚養(yǎng)殖海域沉積物中ARGs主要的耐藥機制為外排泵,這與杭州灣、我國沿海水域沉積物樣品中ARGs 的主要耐藥機制一致[28-29]。研究表明,抑制外排系統(tǒng)中外排泵蛋白的表達量可以降低細菌對藥物的外排作用,減緩耐藥性的產生[30]。
環(huán)境因素可能會驅動和增強ARGs 在環(huán)境中的傳播[31]。本研究測定了所取大黃魚養(yǎng)殖海域沉積物樣品中的5 種環(huán)境因子,探究了其分布規(guī)律,并解析了這5 種環(huán)境因子與ARGs 之間的關系。結果發(fā)現(xiàn),大黃魚養(yǎng)殖海域沉積物中的TOC、TN 和TP 的含量較高,存在污染情況。TOC、TN 和TP 代表養(yǎng)殖環(huán)境中的污染狀況[11],側面反映出漁業(yè)養(yǎng)殖是近海環(huán)境質量惡化的主要原因之一。對所檢環(huán)境因子與ARGs 進行相關性分析,發(fā)現(xiàn)除PNGM-1 與電導率和TN 呈顯著正相關(P<0.05)外,其余ARGs 均與環(huán)境因子之間相關性不顯著(P>0.05),這一結果與陳嘉瑜[20]的研究結果相似,但與Guo 等[31]的研究結果存在差異。水產養(yǎng)殖環(huán)境的理化因子復雜多變,除所檢的5 種環(huán)境因子外,還包括溫度、化學耗氧量、溶解氧、營養(yǎng)物質和重金屬等多種環(huán)境因子,且這些因子與氣候條件、飼料組分、投餌量和吸收率等都有較密切的關系[11]。考慮到環(huán)境因子的復雜性,在自然條件下確定哪種因素決定了ARGs 的賦存特征很難,未來需要擴大環(huán)境因子的檢測種類,增加樣品數(shù)量,構建綜合模型來更好地研究ARGs在環(huán)境介質中的行為。
ARGs 與細菌群落之間存在著連鎖效應、協(xié)同選擇和進化的關系[32-33]。本文利用16S rRNA 基因高通量測序技術對不同位置的大黃魚養(yǎng)殖海域沉積物的細菌群落結構進行分析,結果顯示不同采樣點的微生物組成結構存在明顯差異,其中OS2、OS3和OS4樣品中的微生物種類要顯著高于OS1、OS5 和OS6(P<0.05)。進一步將6 個位點的大黃魚養(yǎng)殖海域的水深(表1)與沉積物細菌群落Alpha 多樣性指數(shù)(表3)進行相關性分析,結果顯示物種總數(shù)指數(shù)(OTUs、ACE指數(shù)和Chao 指數(shù))均與水深呈極顯著或顯著負相關(P<0.01 或P<0.05),說明海水深度越深,微生物種類越少[34]。盡管不同采樣位置大黃魚養(yǎng)殖海域沉積物中的菌群結構和優(yōu)勢菌群存在顯著差異(P<0.05),但是不同海域沉積物中的ARGs基因類型基本一致。這個結果表明即使在不同地理位置上,同一養(yǎng)殖品種和同一養(yǎng)殖模式的海水養(yǎng)殖沉積物中ARGs的賦存狀況也較相似,這也從側面反映出海水養(yǎng)殖沉積物ARGs 的主要來源為水產養(yǎng)殖。這個結果與Hedberg 等[8]的研究結果一致。
ARGs 在不同種類的細菌群落之間遷移、轉化和傳播,使其在環(huán)境中持續(xù)殘留,這比抗生素自身的殘留對生態(tài)環(huán)境的危害更大[35]。因此,對大黃魚養(yǎng)殖海域沉積物中ARGs 和微生物群落之間進行相關性分析至關重要。本研究的結果顯示,豐度前20 的微生物菌屬中有3 種菌屬與ARGs 呈顯著正相關(P<0.05),5 種菌屬與ARGs 呈顯著負相關(P<0.05)。徐秋鴻等[36]認為當微生物與目的ARGs 呈顯著正相關時,這些微生物可能是ARGs 的潛在宿主;當微生物與目的ARGs 呈顯著負相關時,這些微生物可能會影響ARGs 的水平轉移。在本研究中,盡管有些菌屬在沉積物樣品中的相對豐度較低,但其卻與多種ARGs存在顯著相關性,說明在研究微生物與ARGs 的關系時,應盡可能擴大研究范圍。此外,部分菌屬與多種抗生素抗性類型ARGs 顯著相關涉及多種耐藥機制,而有些菌屬僅與1 種抗生素抗性類型ARGs 顯著相關,表明部分微生物可能是多種ARGs 的共同潛在宿主,這種存在也可能對ARGs 的分布特征和水平轉移產生影響,導致細菌產生多重耐藥性[37]。而少數(shù)ARGs 與2 種菌屬之間呈顯著或極顯著正相關(P<0.05 或P<0.01),顯示這些ARGs 可能有多個潛在的微生物宿主[38]。不同微生物攜帶ARGs 的偏好不同[36],后續(xù)研究也應積極探索ARGs 對養(yǎng)殖環(huán)境微生物之間轉移的機理,為通過控制相關菌屬豐度來消減ARGs提供理論基礎。
4 結論
(1)利用宏基因組高通量測序方法能夠獲得更加全面的ARGs 信息。在大黃魚養(yǎng)殖海域沉積物中,共檢測出38種抗生素抗性類型,781個ARGs亞型,豐度最高的ARG 為macB(254.31×10-6~436.85×10-6),其次為tetA(58)(131.64×10-6~241.69×10-6)、evgS(87.00×10-6~193.42×10-6)和bcrA(76.75×10-6~128.89×10-6),不同站點沉積物中的優(yōu)勢ARGs 種類基本一致,主要的ARGs 抗生素抗性類型為四環(huán)素類、大環(huán)內酯類和氟喹諾酮類,主要的ARGs耐藥機制為外排泵。
(2)對大黃魚養(yǎng)殖海域沉積物中的pH、電導率、TOC、TN和TP進行檢測,結果顯示大黃魚養(yǎng)殖海域沉積物已受到污染;相關性分析結果顯示,除PNGM-1與電導率和TN 呈顯著正相關(P<0.05)外,其余ARGs均與環(huán)境因子之間相關性不顯著(P>0.05)。
(3)Proteobacteria 是各大黃魚養(yǎng)殖海域沉積物樣品中豐度最高的細菌門,但各站點沉積物中的微生物組成結構存在明顯差異,且微生物群落結構對環(huán)境ARGs 的產生和分布具有顯著影響,豐度前20 的微生物菌屬中有8 種與ARGs 呈顯著或極顯著相關(P<0.05 或P<0.01),豐度前20 的ARGs 均與豐度前20 的菌屬呈顯著或極顯著相關(P<0.05或P<0.01)。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環(huán)境監(jiān)察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38