甘藍型油菜葉綠素含量的主基因+多基因遺傳分析
2023-03-21 01:51:20李遠紅余坤江何雨琦王曉東
江蘇農業科學 2023年4期
李遠紅,余坤江,何雨琦,王曉東
(1.貴州大學農學院,貴州貴陽 550025; 2.江蘇省農業科學院經濟作物研究所,江蘇南京 210014)
油菜是我國食用植物油的第一大來源,同時兼具觀、菜、蜜、飼、肥等[1]多維度利用價值。2020年,我國油菜種植面積、油菜籽產量已達3 610.7萬hm2、7 179.6萬t,在我國食用油市場中具有舉足輕重的地位[2-3]。近年來,利用雜種優勢來大幅度提高油菜產量難度較大,高光效育種受到越來越多研究者的重視[4]。葉片是植物進行光合作用的主要器官,決定植物體內干物質的積累。葉綠素在綠色植物進行光合作用時起到至關重要的作用,植物吸收太陽輻射后體內會激發反應中心色素從而啟動光化學反應,為生物圈提供動力。葉綠素含量的高低直接影響植物光合潛力和初級生產[5],對油菜籽粒的產量也具有顯著影響[6-7]。因此,解析油菜葉綠素含量的遺傳機制對于推進我國的油菜高光效育種具有重要的理論及實踐意義。
關于油菜葉綠素含量的研究報道較多,國內外學者已對其性狀的遺傳做了相關研究。Zhao等分離出油菜黃綠葉(yvl)突變體,通過BSA-seq定位在BnaA03,全基因組測序發現CHLH在突變體中堿基替換,導致CHLH活性降低,從而影響葉綠體發育和葉綠素的合成[8]。Ye等構建了近等基因系 (NIL),對葉綠素基因座進行了精細定位,研究發現cqSPDA2是歐洲油菜葉綠素含量的一個顯性QTL,在A02上被精細定位到 21.89~22.04 Mb 區間[9]。郎麗娜在研究油菜在鹽脅迫中的QTL時,在LG5發現與葉綠素相關基因座qSPAD5[10]。代文東等利用紫紅葉油菜和綠葉油菜通過DarT-seq技術對BC1群體單株進行基因分型并構建連鎖遺傳圖譜,共檢測出8個葉綠素含量QTL,分別位于A0l、A03、A06、A07、C01、C03和C04連鎖群上[11]。此外,有學者利用構建的分子遺傳連鎖圖譜,定位了油菜種胚[12]、角果皮[13]部位葉綠素含量性狀相關的QTL。這些研究均根據田間各遺傳群體測量葉綠素含量來進行遺傳分析,而通過數量性狀遺傳分離分析方法進行深入系統研究的報道較少。植物主基因+多基因混合模型是蓋鈞鎰等提出來解決數量性狀遺傳的分析方法[14],并在多種作物中已普遍運用。雖已有研究對油菜葉綠素進行QTL定位,但葉綠素合成和調控機制仍較復雜。彭門路等以有、無突變體(yl1)雜交的6個世代(P1、P2、F1、F2、B1、B2)為材料,利用主基因+多基因混合遺傳模型對突變性狀進行遺傳分析,結果表明甘藍型油菜葉綠素缺失性狀受1對加性-顯性主基因控制,yl1葉綠素缺失為不完全顯性遺傳[15]。筆者旨在應用該分析方法研究甘藍型油菜葉片葉綠素含量,解析其遺傳變異規律,為相關性狀的分子標記開發和高光效育種改良奠定基礎。
1 材料與方法
1.1 試驗材料
本研究以甘藍型油菜高葉綠素含量品系APL01、低葉綠素含量品系Holly及2個品系雜交后衍生的198個重組自交家系群體(AH群體目前已發展到F2:13代)為材料,種植于貴州省貴陽市貴州大學教學試驗場(26°N、106°E、海拔為1 100.55 m、土壤為黃壤)。采用隨機區組試驗設計,每個品系種植4行,行距40 cm,株距20 cm,每行15株。試驗地肥力均勻一致,田間管理參照當地大田栽培管理措施。所有試驗材料均由江蘇省農業科學院經濟作物研究所提供。
1.2 葉綠素含量的測定
在2019、2020、2021年3個年份環境下,使用葉綠素計(型號:SPAD-502;品牌:柯尼卡美能達;產地:日本)測定APL01和Holly以及AH群體各品系9張葉片期葉片的葉綠素相對含量(SPAD值),每個品系測量3株,每株測量3張葉(由上至下第3、4、5張葉),每張葉測量3個點,取其平均值,測量時避開主葉脈。2019、2020、2021年3個年份環境分別用19GY、20GY、21GY表示。
1.3 統計分析
對2019—2020年3個年份環境的甘藍型油菜葉片葉綠素含量,利用Excel、SPSS 21.0軟件[16]進行數據統計及群體變異分析。利用植物主基因+多基因混合遺傳模型,結合SEA-G3DH軟件對親本和AH群體3個年份環境的SPAD值進行遺傳分析[17]。遺傳模型的選擇首先根據AIC值(Akaike’s information criterion)最小原則確定3~4個候選模型,然后對候選模型進行適合性檢驗,以確定最適模型,最后計算最適模型下的遺傳參數。分析軟件由南京農業大學章元明教授提供。
2 結果與分析
2.1 葉綠素含量的變異分析
由表1可知,在3個不同年份環境下種植的AH群體葉綠素含量的變異系數大小順序依次為21GY>19GY>20GY,說明第3個年份環境測量葉綠素含量時其變異豐度最大,而3個年份環境間群體葉綠素含量存在變異,表明該性狀受環境影響。從平均值上看,19GY,21GY的平均葉綠素含量較接近,差值僅達2.16,20GY的均值較高于19GY和21GY,20GY與19GY的差值為7.32,20GY與21GY的差值為5.16。21GY的變異系數是3個環境中最高,可見在3個年份環境下所測得的葉綠素含量,第3個環境影響最大,其次是第1個環境,最后是第2個環境。
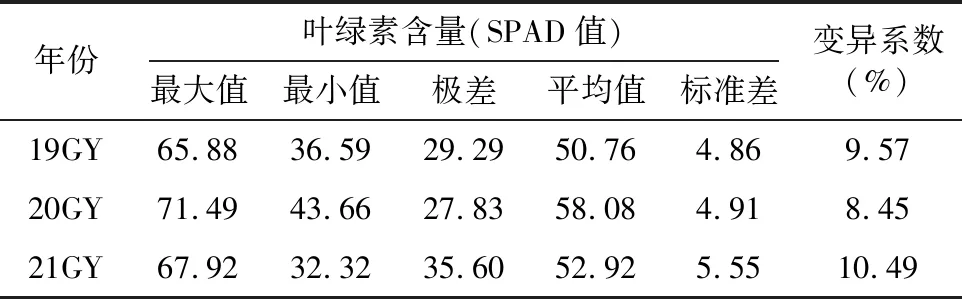
表1 在3個年份環境下AH群體的葉綠素含量變異情況
2.2 葉綠素含量分布特征分析
從圖1可以看出,在3個年份環境下的葉綠素含量在43~66的區間呈連續的分布,表現為典型的數量性狀特征。19GY、21GY的葉綠素含量主要都集中在40~60的區間范圍內,而在0~40和60以上的分布頻數較低。20GY的葉綠素含量主要集中在50~65的區間范圍內。其群體葉綠素含量分布呈現為正態分布,表明該群體中葉綠素含量的遺傳符合主基因+多基因遺傳特征。

2.3 葉綠素含量遺傳模型分析
2.3.1 遺傳模型的選擇 采用是植物數量性狀主基因+多基因混合遺傳模型對親本及AH群體的葉綠素含量性狀進行遺傳分析,估算獲得了0對主基因(0MG模型)、1對主基因(1MG模型)、2對主基因(2MG模型)、多基因(PG模型)、1對主基因+多基因(MX1模型)、2對主基因+多基因(MX2模型)、3對主基因(3MG模型)、3對主基因+多基因(MX3模型)、4對主基因(4MG模型)等9類共39種遺傳模型的極大似然函數值和AIC值(表2)。根據Akaike準則,選取AIC值最小及與最小AIC值比較接近的遺傳模型作為候選模型,每個環境確定4個候選模型。用2019年葉綠素含量數據獲得19GY的候選模型為3MG-A、3MG-CEA、MX3-AI-AI、4MG-AI,用2020年葉綠素含量數據獲得20GY的候選模型為3MG-A、3MG-CEA、MX3-AI-AI、MX3-AI-A;21GY獲得候選模型為MX3-AI-AI、MX3-AI-A、MX3-A-A、4MG-A。

表2 葉綠素含量各遺傳模型的AIC值
從上述結果來看,其中2個年份環境(20GY、21GY)的最適模型皆為MX3-AI-AI,而19GY年份環境下所檢驗出的最適模型為4MG-AI,而3個年份環境下2個候選模型的AIC值(MX3-AI-AI為1 248.57,4MG-AI為1239.92)較接近,造成這個分析結果的原因可能是因為19GY的環境差異較大,葉片的葉綠素含量所受環境的影響也相對加強,結合3個環境結果,推測出AH群體葉綠素的最適模型為MX3-AI-AI。
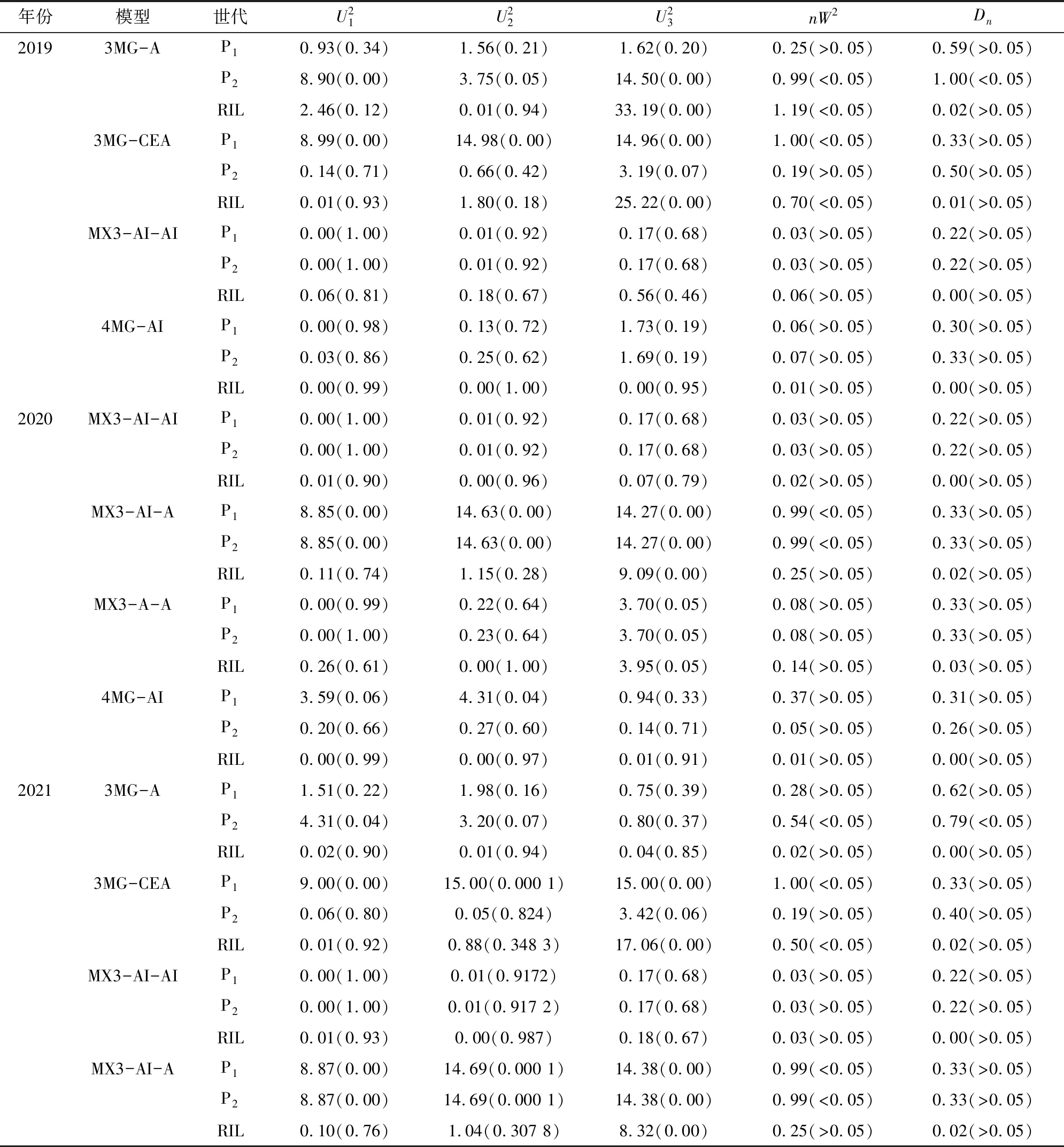
表3 AH群體的模型適合性檢驗
2.3.3 最適模型下遺傳參數的估計 分別計算油萊AH群體在3個年份環境下葉片葉綠素含量在最適遺傳模型下的各遺傳參數,結果(表4)表明,在19GY環境下,群體葉綠素含量的3對主基因的加性效應均表現為負向遺傳效應,效應大小順序為da>dc>db;主基因上位性效應分別為-6.35、-6.35、-6.34、-6.34;主基因遺傳率為63.22%;多基因遺傳率為35.7%。 20GY環境下的群體葉綠素含量3對主基因的加性效應均表現為負向遺傳效應,加性效應大小順序為da>dc>db;主基因上位性效應分別為-6.60、-6.61、-6.60、-6.60;主基因遺傳率為73.09%;多基因遺傳率為26.32%。21GY環境下的群體葉綠素的3對主基因的加性效應均表現為負向遺傳效應,加性效應大小順序為da>dc>db;主基因上位性效應分別為-7.24、-7.25、-7.24、-7.24;在19GY、20GY中,主基因加性效應和主基因上位性效應值都是負值,說明2個效應都能使葉綠素含量降低。在21GY中,主基因加性效應值都為負值,主基因上位性效應為正值,說明主基因加性效應使葉綠素含量降低,則主基因上位性效應提高葉綠素含量。綜上所述,AH群體葉綠素含量受主基因加性和上位性效應調控,同時,存在明顯的多基因加性效應影響。
連續2年3個環境的數據中,主基因遺傳率為63.22%~73.33%,多基因遺傳率在25.94%~35.7%,主基因遺傳率平均值69.88%,顯著大于多基因遺傳率,可見葉片葉綠素含量的遺傳體系中,主基因的貢獻率占主要部分。此外,AH群體葉綠素含量在3個環境方差都為0.66左右,說明葉綠素含量易受環境影響。
3 討論與結論
3.1 油菜葉片葉綠素含量變異豐富
自20世紀70年代以來,許多國家將遺傳改良的重點放到了提高作物光合效率上,并且明確指出提高光效是作物育種的重要內容[18]。本研究利用APL01、Holly品系及2個品系雜交后衍生的198個重組自交家系群體,葉綠素含量在2019、2020、2021年的變異系數分別為9.57%、8.45%、10.49%,葉綠素含量(SPAD值)為50.76、58.08、52.92。從結果分析來看,薦紅舉等對連續2年588份甘藍型油菜品種的葉綠素分析后發現,在不同年份中葉綠素含量變化較小,變異系數分別為11.27%、12.73%[19]。有研究對紫紅葉和綠葉油菜品系構建遺傳分離群體的葉綠素含量分析,葉綠素含量的變異系數為8.00%[11]。研究結果表明,油菜葉綠素含量變異系數較穩定,具有豐富的變異,本研究與前人研究結果相符。而造成葉綠素含量變異的原因,主要是受環境的影響[20-21]。

表4 AH群體葉綠素含量在MX3-AI-AI模型下遺傳參數估計值
3.2 葉綠素含量受主基因+多基因共同控制
葉色調控在光合作用和干物質生產中具有重要作用,明確其遺傳機制有利于后續育種實踐。關于植物葉綠素或黃化性狀的遺傳,國內外已有較多研究,但針對甘藍型油菜葉片葉綠素含量相關QTL或候選基因的研究相對較少。本研究采用植物主基因+多基因混合遺傳模型,結合SEA-G3DH軟件得出AH群體葉綠素含量最優遺傳模型為MX3-AI-AI,即3對加性-上位性主基因+多基因加性-上位性混合遺傳模型,與前人研究結果不盡相同。田穎等利用油菜黃化突變體L638-y與正常葉色材料2598雜交后代用于黃化基因標記,發現葉色由2對隱性核基因gr1、gr2調控[22-23]。相關研究認為,調控葉綠素含量受1對加性-顯性主基因控制,不存在多基因效應的影響[14]。唐亮等研究了水稻群體葉綠素含量基因的遺傳規律,發現葉綠素含量受2對主基因+多基因共同控制[24]。可見,葉綠素含量的遺傳機制相對復雜。代文東等研究檢測出8個葉綠素含量QTL,證實葉綠素含量同時也受加性效應的調控,加性效應在4.99~9.20[10]。本研究加性效應在6.35~7.25,與前人研究結果不同,為進一步挖掘相關QTL提供參考。造成這些異同點的原因,首先是試驗材料的不同,揭示甘藍型油菜葉綠素遺傳機制的研究較少。其次,研究方法大都采用蓋鈞鎰等編制的植物數量性狀分離分析軟件。本研究采用南京農業大學劉兵等研制的植物數量性狀分離分析Windows軟件包SEA,該軟件具有友好的操作界面,能夠計算出一階遺傳參數和部分二階遺傳參數,結果更易分析。葉綠素屬數量性狀,環境因素干擾大,大多研究群體較小,本研究擴大育種群體規模,更利于考察該性狀。甘藍型油菜葉綠素含量性狀是由主基因控制和多基因共同控制,且主基因遺傳率顯著高于多基因遺傳率。本研究明確了甘藍型油菜主基因間作用方式為累加作用,并且得到了主基因加性效應、主基因上位性效應各效應值,這為深入研究甘藍型油菜葉綠素含量性狀的遺傳機制提供了基礎。研究結果表明,葉綠素含量的遺傳力高,育種過程中在早期世代對葉綠素含量高的材料進行選擇是有效的,在本試驗中,后代存在葉綠素含量超過親本的材料,說明可以采用雜交和回交方式轉移主基因,獲得超親材料。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
光學精密工程(2016年6期)2016-11-07 09:07:19
