不同粒度貓尾草對羔羊體外發酵特性和微生物數量的影響
2023-03-21 09:19:10王靜孔令瑩徐建風康靜沈振峰劉婷
草業學報 2023年3期
王靜,孔令瑩,徐建風,康靜,沈振峰,劉婷
(甘肅農業大學動物科學技術學院,甘肅 蘭州 730070)
貓尾草(Uraria crinita)為多年生禾本科牧草,因其葉量豐富、草質細嫩、適口性好,常作為賽馬和兔子等觀賞動物飼料的纖維來源[1]。奶牛采食貓尾草能增加逆嘔次數,延長產奶期,提高生產性能和產奶量[2-3]。但以貓尾草作為幼齡反芻動物開食料纖維素來源的研究卻鮮有報道。中性洗滌纖維(neutral detergent fiber,NDF)是評價粗飼料營養價值的重要指標,其作用效果受到粒度、來源和水平等因素的影響[4-5],其中,粗飼料粒度在促進瘤胃發酵和維持瘤胃內環境穩態過程中發揮著重要的作用[6-7]。前期研究發現,粗飼料物理形態和粒度影響斷奶后幼齡反芻動物瘤胃發酵和微生物區系,但研究結果存在較大的差異[8],這是不同粗飼料的物理和化學性質差異所致。基于此,探求貓尾草適宜粒度是利用其作為幼齡反芻動物開食料纖維素來源的關鍵問題。本試驗在相同營養水平基礎上,通過研究含有不同粒度貓尾草的全混合日糧(total mixed ration,TMR)對3 月齡羔羊體外瘤胃發酵參數和瘤胃微生物菌群數量的影響,以期為貓尾草在幼齡反芻動物開食料中的合理利用提供科學依據。
1 材料與方法
1.1 試驗動物及飼糧
于2021年1 月,在甘肅農業大學動物科學實訓教學中心開展試驗。選擇3 只健康且體重相近[(13.42±0.73)kg]的3 月齡瘺管羊作為瘤胃液供體動物,每天飼喂3 次,自由采食和飲水,保證羊舍衛生干凈并定期消毒。貓尾草購自甘肅潤牧生物工程有限責任公司,帶回實驗室測定初水分后,將其斬(碾)碎過篩,制備6 個不同粒度,分別為1.00、2.36、3.35、4.75、8.00 和12.50 mm。參照NRC[9]13 kg 綿羊羔羊飼養標準配制TMR 作為發酵底物(表1),其中貓尾草是試驗飼料纖維素唯一來源。各處理組飼料除貓尾草粒度不同外,其他飼料原料粒度(0.425 mm)均保持一致。

表1 飼糧組成及營養成分Table 1 Dietary composition and nutrient composition(dry matter basis)
1.2 試驗設計
采用單因素和雙因素試驗設計,按照貓尾草的粒度將試驗分為6 個處理組,分別為1.00 mm 組、2.36 mm 組、3.35 mm 組、4.75 mm 組、8.00 mm 組和12.50 mm 組。應用Rusitec-s 人工瘤胃系統(Sanshin,東京,日本),將8 個發酵罐分為2 個處理,每個處理組4 個重復,每2 個處理組共同發酵48 h,在同等試驗條件下共進行3 批,進行為期16 d 的試驗,前10 d 為人工瘤胃系統穩定期,后6 d 為正試期。
1.3 人工瘤胃培養試驗
1.3.1 瘤胃液采集 晨飼前利用采集器收集試驗羊瘤胃液,經4 層細紗布過濾至已預熱[(39±0.5)℃]處理過的保溫桶內,在此過程中持續通入CO2以保證厭氧環境,然后立即密封。與此同時,收集固體食糜后進行保溫處理,帶回實驗室置于(39±0.5)℃恒溫水浴箱中進行分裝。
1.3.2 體外發酵培養 按照Mc Dougall[11]的方法配制人工瘤胃緩沖液。并在試驗開始前置于水槽內預熱至(39±0.5)℃,與瘤胃液按1∶1 分裝至發酵罐,并將裝有70 g 固體食糜的尼龍袋與10 g 樣品一并投入發酵罐內。利用Rusitec-s 人工瘤胃系統,參照Kajikawa 等[12]的體外發酵方法進行48 h 的體外發酵。其中,緩沖液流量為0.39 mL·min-1,流出液口篩網孔徑為1.2 mm,攪拌機攪拌頻率為4~5 次·min-1,發酵罐內保持(39±0.5)℃和厭氧環境。
1.3.3 樣品采集與測定 分別在體外發酵3、6、9、12、24 和48 h 時,使用酸度計(PHS-3C,雷磁儀器廠,上海,中國)測定每個罐中發酵液pH。48 h 發酵結束后,收集各罐中發酵液和發酵殘渣用于測定發酵參數和瘤胃微生物菌群數量。采集發酵液5 mL 至液氮罐中,帶回實驗室后保存在-70 ℃冰箱,用于測定瘤胃微生物菌群的數量。另外,采集2 mL 發酵液,4 ℃、12000 r·min-1離 心10 min 取 上 清 液1 mL 于2 mL 離心管內,-20 ℃保存,用于發酵液揮發性脂肪酸(volatile fatty acid,VFA)和 氨 態 氮(ammonia nitrogen,NH3-N)濃度的測定。參照馮宗慈等[13]的方法測定NH3-N 濃度。參照鄭琛等[14]的方法使用氣相色譜儀(6890N,Agilent,美國)測定VFA,色譜柱為HP 19091N-213 毛細管柱。色譜分析條件為:進樣口溫度220 ℃,N2流量2.0 mL·min-1,分流比40∶1,程序升溫模式為120 ℃保持3 min,然后10 ℃·min-1升溫至180 ℃,保持1 min,火焰氫離子檢測器(flame ionization detector,FID)250 ℃,FID 空氣、H2和N2流量分別為450、40 和45 mL·min-1[15]。
收集發酵殘渣烘干后,分別按照Cunniff[16]和Van Soest 等[17]的方法測定干物質(dry matter,DM)、粗蛋白質(crude protein,CP)、中性洗滌纖維(NDF)和酸性洗滌纖維(acid detergent fiber,ADF)含量,并計算發酵底物中各營養成分的體外降解率,計算公式如下:
1.4 微生物菌群數量的測定
參照Yuan 等[18]的方法提取發酵液微生物總DNA。使用超微量紫外分光光度計(NanoDropTMOne,美國)測定發酵液微生物總DNA 吸光度(OD),OD260/OD280均為1.8~2.0,可進行下一步分析。按照SYBRGreenⅡ預混試劑盒建議的20 μL 反應體系,使用實時定量PCR 對發酵內容物總菌(total bacteria,TB)、溶纖維丁酸弧菌(Butyrivibrio fibrisolvens,B.f)、產琥珀酸絲狀桿菌(Fibrobacter succinogenes,F.s)、白色瘤胃球菌(Ruminococcus albus,R.a)、總產甲烷菌(totalMethanogens,TMe)和甲烷桿菌(Methanobacteriaceae,Mba)進行測定。目的微生物的數量分別以16S rRNA 或18S rRNA 基因拷貝數(拷貝數·mL-1)表示,引物序列見表2。

表2 瘤胃微生物引物序列Table 2 Rumen microbial primer sequence
1.5 數據統計與分析
使用SPSS 21.0 軟件對試驗數據進行分析,48 h 養分降解率、發酵液參數和微生物絕對數量用單因素方差分析,發酵液動態pH 值用雙因素方差分析,差異顯著時采用Tukey 法進行多重比較,P<0.05 表示差異顯著。
2 結果與分析
2.1 不同粒度貓尾草對體外營養物質降解率的影響
由表3 可知,體外干物質降解率(in vitrodry matter degradability,IVDMD)隨著貓尾草粒度的增加而降低,其中4.75 和12.50 mm 組 顯 著 低于1.00 mm 組(P<0.05);而 體 外 粗 蛋 白 質 降 解 率(in vitrocrude protein degradability,IVCPD)隨著貓尾草粒度的增加而升高,其中,12.50 mm 組顯著高于1.00、2.36 和3.35 mm 組(P<0.05);體外中性洗滌纖維降解率(in vitroneutral detergent fiber degradability,IVNDFD)隨著貓尾草粒度的增加而逐漸降低,其中,1.00 和2.36 mm 組顯著高于8.00 和12.50 mm 組(P<0.05)。而體外酸性洗滌纖維降解率(in vitroacid detergent fiber degradability,IVADFD)在各處理組間差異不顯著(P>0.05)。
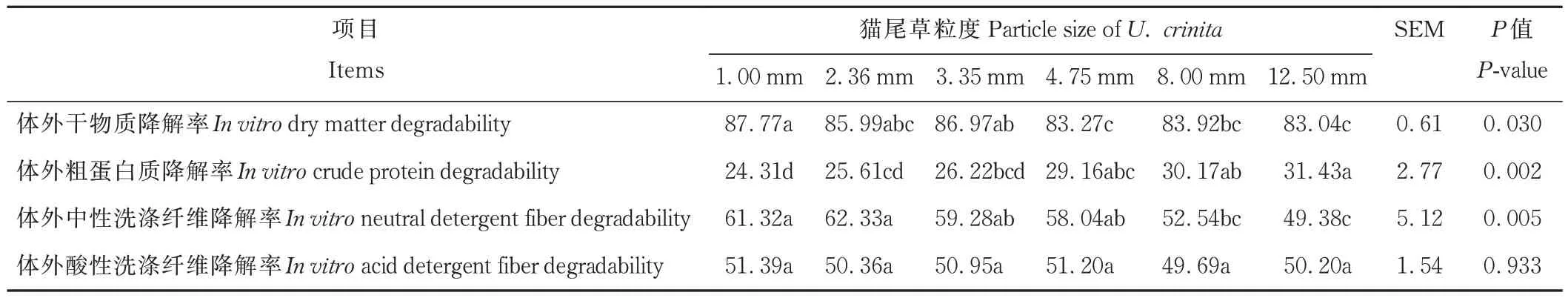
表3 不同粒度貓尾草48 h 體外發酵對養分降解率的影響Table 3 Effects of fermentation in vitro for 48 h on nutrient degradability of U.crinita with different particle sizes(%)
2.2 不同粒度貓尾草對體外發酵液參數的影響
由表4 可知,不同粒度貓尾草經48 h 體外發酵在6 個時間點的pH 值均無顯著差異(P>0.05),但都呈先降低后升高的趨勢,且時間和粒度間無顯著的交互作用(P>0.05)。
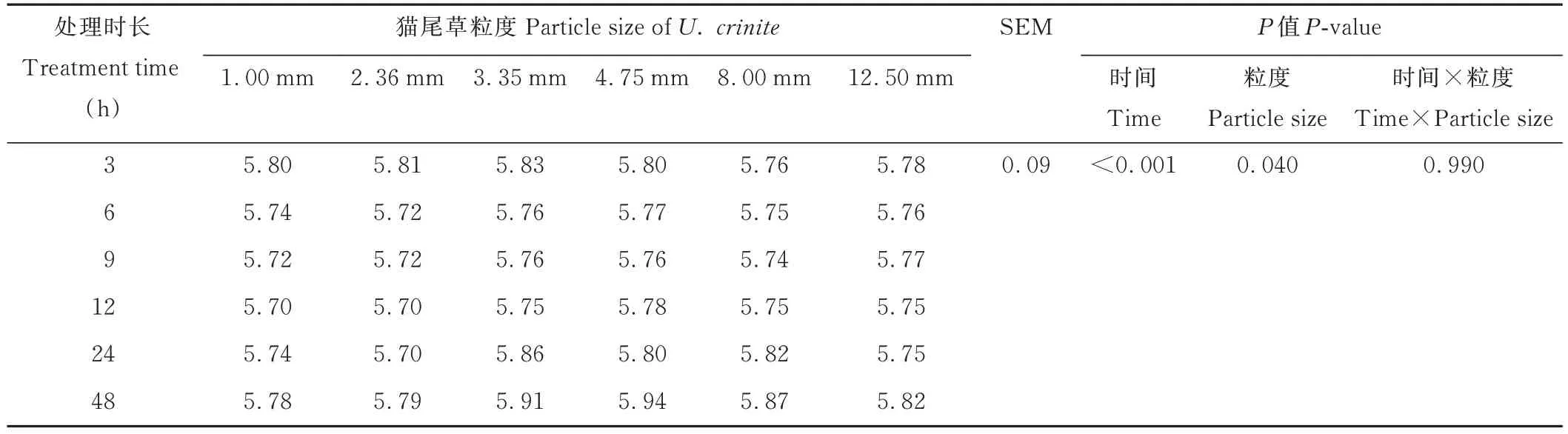
表4 不同粒度貓尾草體外發酵動態pH 值Table 4 Dynamic pH value in vitro fermentation of U.crinita with different particle sizes
由表5 可知,NH3-N 濃度和總揮發性脂肪酸(total volatile fatty acid,TVFA)隨著貓尾草粒度的增加呈先降低后升高的趨勢,其中,12.50 mm 組顯著高于3.35 和4.75 mm 組(P<0.05);丙酸摩爾比也隨著貓尾草粒度的增加呈先降低后升高的趨勢,其中,1.00 mm 組顯著高于4.75 mm 組(P<0.05);而1.00 mm 組異丁酸和丁酸摩爾比顯著低于8.00 和2.36 mm 組(P<0.05),異戊酸和戊酸摩爾比顯著低于2.36 mm 組(P<0.05);乙酸摩爾比和乙酸/丙酸在各處理組間無顯著差異(P>0.05)。
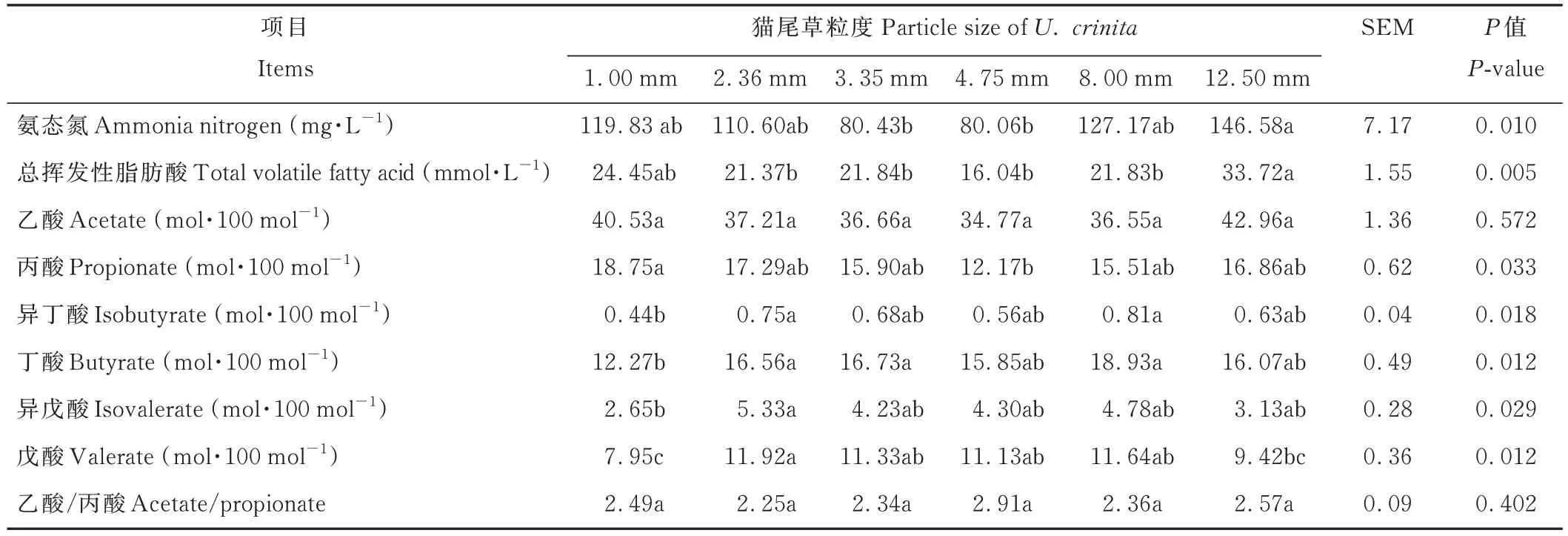
表5 不同粒度貓尾草對48 h 體外發酵液參數的影響Table 5 Effects of different particle sizes of U.crinita on parameters of fermentation broth in vitro for 48 h
2.3 不同粒度貓尾草對發酵液微生物絕對數量的影響
由表6 可知,隨著貓尾草粒度的增加,B.f 和Mba 的數量呈先升高后降低再升高的趨勢,其中,2.36 mm 組B.f顯著高于3.35 和4.75 mm 組(P<0.05),Mba 顯著高于3.35、4.75 和8.00 mm 組(P<0.05)。TMe 數量隨著貓尾草粒度的增加先降低后增加,其中,4.75、8.00 和12.50 mm 組顯著高于1.00 和2.36 mm 組(P<0.05),TB、F.s 和R.a 在各處理組間無顯著差異(P>0.05)。
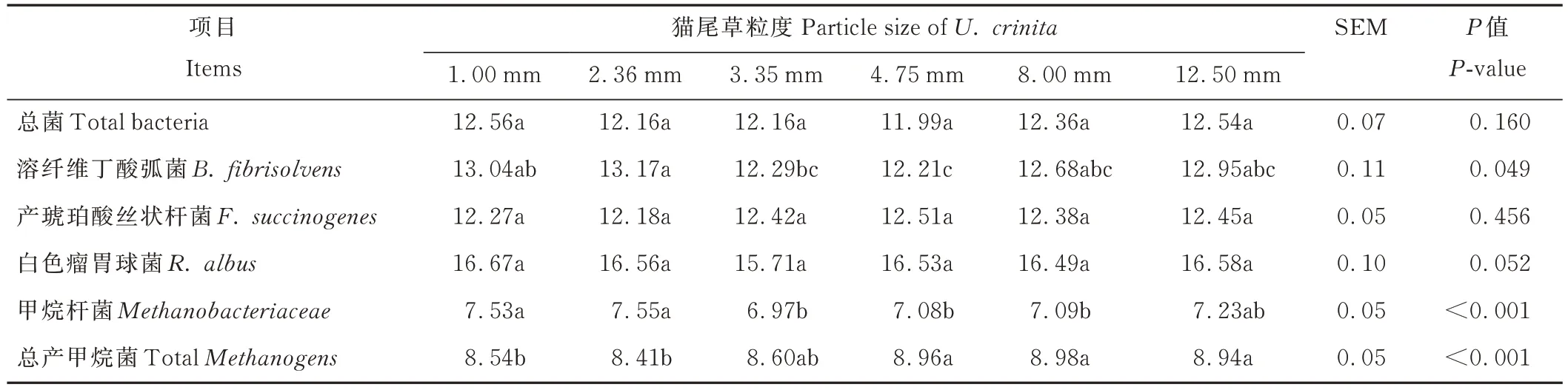
表6 不同粒度貓尾草對48 h 發酵液微生物絕對數量的影響Table 6 Effects of different particle sizes of U. crinita on the absolute number of microorganisms in fermentation broth for 48 h[log10(copies·mL-1)]
2.4 體外瘤胃發酵特征與微生物相關性分析
由表7 可知,TB 與NH3-N 和乙酸呈正相關(R2=0.84,P=0.038;R2=0.88,P=0.021);B.f 與丙酸呈正相關(R2=0.82,P=0.045),與pH 呈 負 相 關(R2=-0.98,P=0.001);TMe 與IVCPD 呈 正 相 關(R2=0.91,P=0.012),與IVDMD 和IVNDFD 呈負相關(R2=-0.86,P=0.027;R2=-0.83,P=0.043);Mba 與pH 呈負相關(R2=-0.90,P=0.015)。
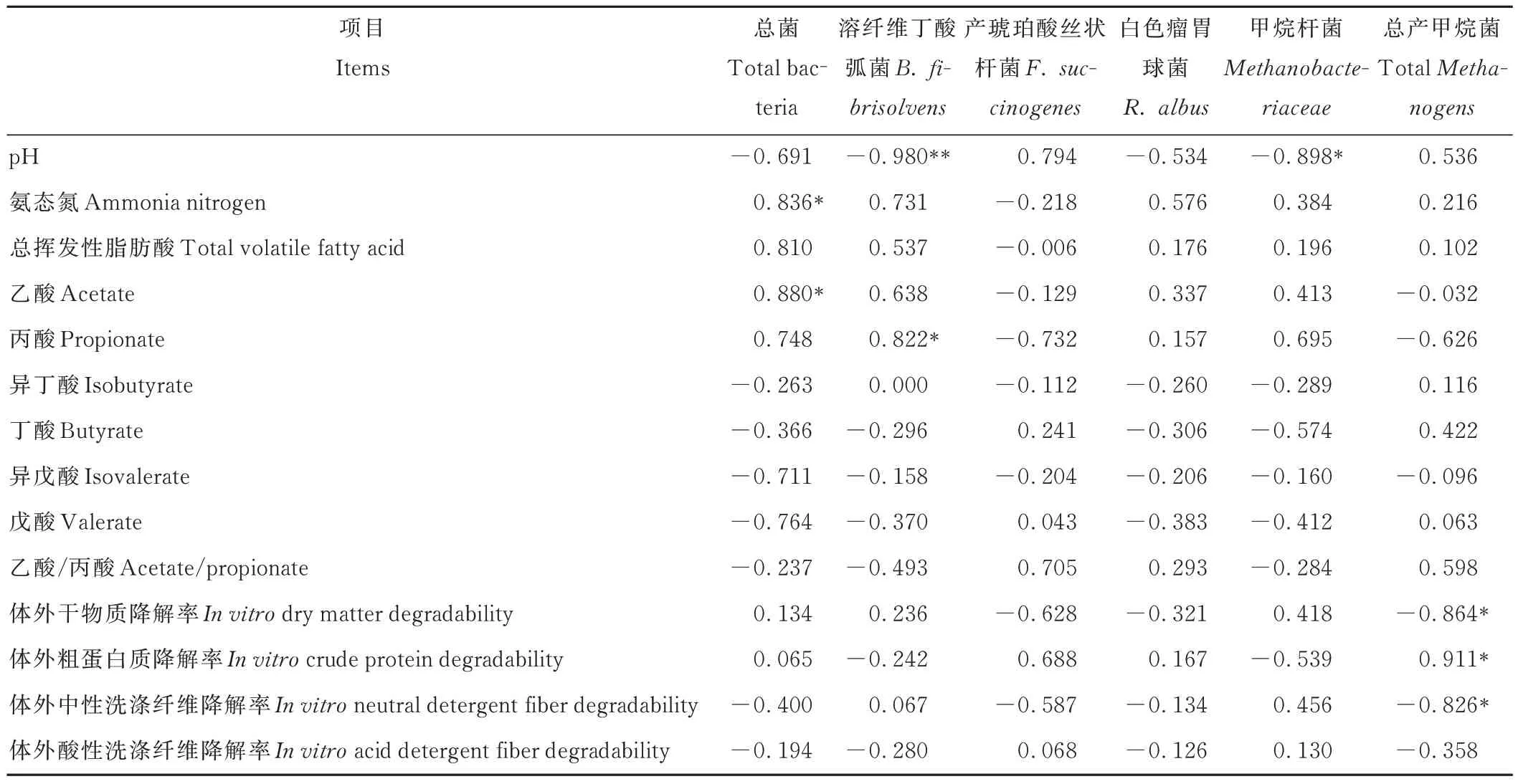
表7 瘤胃發酵特性與微生物相關性分析Table 7 Correlation analysis of rumen fermentation characteristics and microorganisms
3 討論
3.1 不同粒度貓尾草對體外營養物質降解率的影響
瘤胃內干物質降解率是影響反芻動物對營養物質降解與發酵的重要因素[24]。本試驗中,IVDMD 和IVNDFD 隨著貓尾草粒度的增加而降低,與多項研究結果一致,如Zhao 等[25]研究發現,在裝有瘤胃瘺管的肉牛日糧中含加工的大麥(Hordeum vulgare)籽粒粉碎比例越高,原位降解率越高;Ueda 等[26]發現,隨著飼料顆粒尺寸的減小,會增加顆粒與瘤胃微生物的接觸面積,結構型碳水化合物更容易被降解,從而提高瘤胃IVDMD 和IVNDFD。本試驗IVCPD 隨著貓尾草粒度的增加而升高,其中,12.50 mm 組顯著高于1.00、2.36 和3.35 mm組,然而韓海珠等[27]研究發現,在人工瘤胃試驗中大顆粒組(5 mm)日糧IVCPD 大于小顆粒組(0.45 mm),本試驗結果與此相反,可能是長粒度貓尾草在瘤胃內的流通速率慢,能夠充分促進養分消化,因此IVCPD 隨著貓尾草粒度的增加而升高。此外,因試驗所用尼龍袋孔徑為0.045 mm,發酵液流出時過濾網的孔徑為1.2 mm,1.00 mm 粒度組在微生物降解過程中許多小顆粒會隨著發酵液流出,可能導致發酵參數有誤。
3.2 不同粒度貓尾草對體外發酵液參數的影響
pH 是評價瘤胃內環境穩定性的重要指標,它決定著瘤胃微生物對飼糧的發酵和利用情況[28]。本試驗中,不同粒度貓尾草經48 h 發酵后的pH 無顯著差異,但呈現出先降低后升高的趨勢。各粒度貓尾草發酵后的pH 值為5.70~5.94,均處于正常范圍值內(5.5~7.5)[29],且整體偏低,這有利于微生物發酵。pH 值偏低可能與低纖維水平(10% FNDF)飼糧中非纖維性碳水化合物含量較高有關,加之貓尾草木質化程度較低,可降解的纖維含量較高,進入瘤胃后能被微生物快速降解,導致TVFA 迅速增加引起pH 值降低。曾銀等[30]研究表明,飼料粒度與瘤胃pH 值的關系存在一個臨界粉碎粒度,當粗飼料粒度范圍在某臨界值以下,會對瘤胃pH 值水平造成影響。周萬才等[31]研究報道,在自由采食條件下,直徑為3 mm 的顆粒料可以提高肉牛瘤胃發酵速度,降低瘤胃pH 值,當顆粒直徑提高至8 mm 時,可以降低瘤胃發酵速度。而Kononoff 等[32]和Yang 等[33]的研究發現,瘤胃pH 值并未受到玉米(Zea mays)青貯顆粒長短的影響。以上試驗結論的差異可能是因為飼糧顆粒大小存在臨界值造成的。
NH3-N 是瘤胃中粗蛋白質降解的終產物,也是瘤胃微生物合成菌體蛋白的來源,可以反映瘤胃中含氮物質的供應和利用情況[34]。本試驗中4.75 mm 組NH3-N 濃度顯著低于12.50 mm 組,Hildebrand 等[35]也報道了大顆粒組(4 mm)NH3-N 濃度顯著高于小顆粒組(1 mm),本試驗結果與此一致。李杰等[36]研究表明,飼喂較大顆粒(2.5~5.0 mm)的豆粕會降低瘤胃外流速度。因而12.50 mm 的貓尾草在瘤胃中停留時間長,致使微生物的分解時間增加,由此提高了NH3-N 濃度。
VFA 是碳水化合物發酵的最終產物,其含量及組成比例是反映瘤胃消化代謝活動的重要指標[37]。本試驗中,當貓尾草粒度增加到12.50 mm 時,TVFA 含量顯著升高,乙酸摩爾比和乙酸/丙酸值也最大。Rodriguez-Prado 等[38]和Hildebrand 等[35]研究表明,大顆粒組(分別為3 和4 mm)TVFA、乙酸和乙酸/丙酸均高于小顆粒組(1 mm),本試驗結果與此相一致。Benchaar 等[39]和Abderzak 等[40]認為,NDF 水平低的日糧可以提高淀粉分解菌的活性從而促進丙酸和丁酸的合成。本試驗結果顯示,粒度為1.00 mm 時顯著提高了丙酸摩爾比,粒度為2.36 mm 時顯著提高了異丁酸、丁酸、異戊酸、戊酸摩爾比和IVNDFD。這說明在低NDF 水平下隨著貓尾草粒度的減小,提高了淀粉在瘤胃中的消化率和IVNDFD[33,41],進而增加了其降解產物丙酸和丁酸的產生。王加啟等[42]報道,飼糧組成、碳水化合物的來源及加工處理技術和瘤胃環境是影響瘤胃發酵與VFA 產生量和比例的因素。本試驗中各組飼糧組成和碳水化合物來源均相同,VFA 含量及其比例是受到飼料加工處理和瘤胃環境的影響。
3.3 體外瘤胃發酵特征與微生物相關性分析
瘤胃細菌附著于飼料顆粒表面,通過侵蝕方式逐漸降解植物細胞壁[43],且能分泌纖維素酶和蛋白酶等消化酶[44],由多種細菌和酶相互協同作用才能使結構復雜的纖維降解[45],最終產生可用于生長和維持的終產品使宿主受益,因此瘤胃微生物對宿主的生存至關重要。本試驗結果表明,B.f 與丙酸摩爾比呈正相關,與pH 值呈負相關。當粒度為2.36 mm 時,B.f 數量和IVNDFD 都顯著升高,丙酸摩爾比也相對較高,而48 h 時pH 值相對較低。這表明羔羊瘤胃內B.f 降解纖維素的能力在粒度為2.36 mm 時最強,發酵程度較好導致pH 下降[31]。Mba 與pH呈負相關,TB 與乙酸摩爾比呈正相關;當粒度為4.75 mm 時,48 h 的pH 值最高,TVFA 和Mba 數量顯著降低,乙酸摩爾比和TB 數量也最低。此結果表明,粒度為4.75 mm 時瘤胃內發酵程度較低,TVFA 含量低,pH 值上升,幼齡動物對乙酸的利用率隨著產甲烷菌數量的減少而降低。許多瘤胃細菌代謝產生乙酸,生長也需要乙酸[46],成艷芬等[47]研究表明,產甲烷菌可以利用其他微生物的代謝產物乙酸生成甲烷,從而降低動物體對乙酸的利用率。本試驗中,TB 與NH3-N 濃度呈正相關,TMe 與IVCPD 呈正相關,這說明隨著瘤胃內發酵程度的提高,蛋白質降解度增加,NH3-N 濃度也相應升高,為瘤胃微生物提供了豐富的氮源,從而增加了TB 數量,提高了產甲烷菌對氮的利用率。研究表明,飼料粒度影響通過瘤胃的食糜大小和在瘤胃內的停滯時間,從而影響瘤胃微生物的活性[48]。然而,目前不同粒度對低纖維水平飼料發酵特性和瘤胃微生物的影響還存在很大的爭議,且體外試驗和體內試驗結果也存在差異,確定不同粒度貓尾草對瘤胃微生物-宿主的互作關系需要進一步進行動物體內試驗的驗證。
4 結論
低纖維水平飼料中不同粒度的貓尾草能夠調控發酵液中微生物數量,改變其發酵特征從而影響飼料的體外降解率;瘤胃微生物數量與VFA 和養分降解率存在一定的相關性,這種相互作用對幼齡動物的生長發育有一定的作用。
