北方白菜型冬油菜的春化率、抗寒性和遺傳多樣性分析
2023-03-29 06:14:18路曉明武軍艷王新發徐愛遐李學才劉麗君蒲媛媛牛早霞王會文朱明川范軍強孫萬倉
干旱地區農業研究 2023年1期
路曉明,武軍艷,王新發,徐愛遐,李學才,劉麗君,馬 驪,蒲媛媛,牛早霞,王會文,朱明川,范軍強,孫萬倉
(1.甘肅農業大學農學院/甘肅省油菜工程技術研究中心/甘肅省作物遺傳改良與種質創新重點實驗室/省部共建干旱生境作物學國家重點實驗室,甘肅 蘭州 730070; 2.中國農業科學院油料作物研究所,北京 100000; 3.西北農林科技大學農學院,陜西 楊凌 712100)
我國油菜生產依據播種季節,劃分為冬油菜與春油菜兩大產區[1],以甘肅省的天水附近為分界線[2]。由于強抗寒品種的育成及其在北方地區的推廣應用,我國冬油菜的種植區域擴大,初步形成了北方冬油菜產區。這個區域冬油菜分布范圍廣,西起新疆阿勒泰,東至河北、北京等北方多個省市區,生態條件復雜,品種生態型豐富。由于冬油菜含油率高、產量高,而且可利用冬閑田生產,在冬春季具有優異的地表覆蓋、保土、保水、保肥功能,對我國食用植物油生產及環境保護具有重要意義。
北方冬油菜生產的首要問題是如何選擇和應用抗寒品種,確保安全越冬。有關白菜型油菜種質的抗寒性評價已有研究,趙彩霞[3]研究結果表明,SOD活性、氣孔密度、單位面積氣孔總長度、葉片組織結構的柵/海比和葉片組織細胞結構疏松度等均可作為油菜抗寒性鑒定指標。陳奇[4]研究表明越冬率是評價冬油菜抗寒性最可靠直觀的依據。許耀照等[5-6]研究表明,白菜型冬油菜抗寒性與冬前低溫下葉片光合特性以及干物質積累有關,并提出采用準確的、無傷害的葉綠素熒光技術鑒定冬油菜的抗寒性。楊寧寧[7]研究指出根據春播條件下春化率高低可以判斷油菜品種的冬性強弱。不同品種經相同春化時間處理后,其現蕾率和開花率表現不同[8],說明不同品種之間春化率存在差異。春化率是油菜抗寒性強弱的直觀體現,且影響其種植范圍和經濟價值。目前,在北方冬油菜產區,生產上應用的品種繁雜,抗寒性和親緣關系不明,對育種研究及品種利用造成了一定影響。因此本研究基于春播條件下的春化率、半致死溫度結合分子標記技術評價白菜型冬油菜抗寒性及遺傳多樣性。
簡單重復序列(Simple sequence repeat, SSR)是以PCR擴增技術為基礎的共顯性分子標記,具有多態性豐富和擴增結果重現性高的優點[9-10],在植物中用于鑒別其資源的遺傳多樣性。SRAP(Sequence-related amplified polymorphism)相關序列擴增多態性標記,是針對蕓薹屬開發的一種標記,主要以擴增開放閱讀框(ORF)為主。基于全基因組重測序開發的插入/缺失[Insertion/Deletion(InDel)]標記在類群劃分、品種純度鑒定、親緣關系鑒定方面廣泛應用[11]。
本研究基于春播條件下的春化率結合半致死溫度的測定、SRAP、SSR和InDel 3種標記及冬前植物學性狀特性鑒定,對目前生產上應用的北方白菜型冬油菜種質資源進行抗寒性評價及遺傳多樣性分析,以探究不同抗寒性材料之間的差異,為北方白菜型冬油菜抗寒性鑒定及強抗寒品種選育和品種選擇提供理論依據。
1 材料與方法
1.1 參試材料
本研究選取北方各地種植面積較大的26個白菜型冬油菜品種(系)為試驗材料,分別來自甘肅天水、甘肅慶陽、甘肅蘭州和陜西洛川四地,詳情見表1。
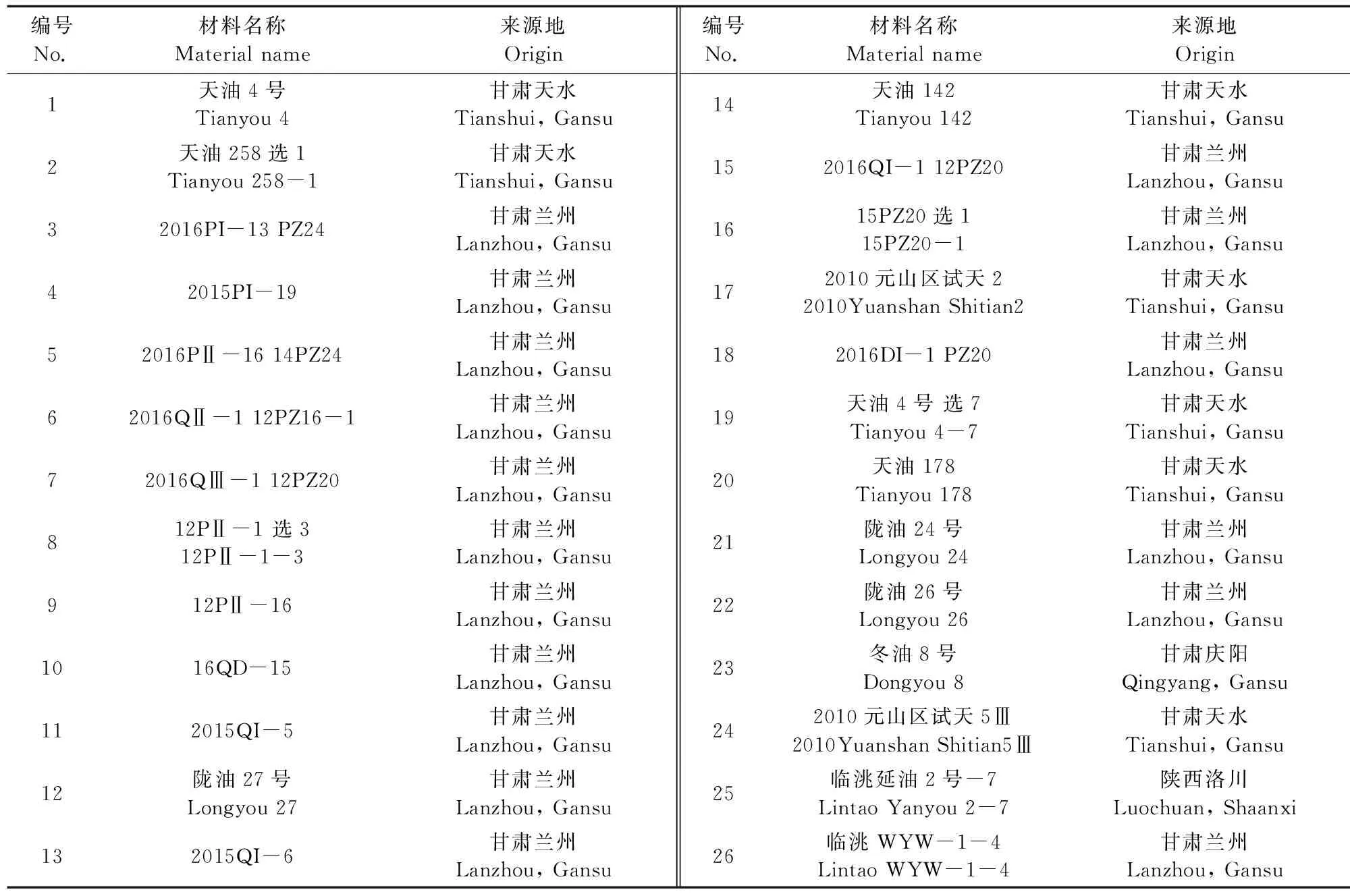
表1 參試材料Table 1 Test materials
1.2 試驗方法
1.2.1 春化率測定 春播試驗于2018年4月20日在蘭州市永登縣上川鎮播種,小區面積10m2,3次重復,隨機區組設計。于9月10日統計每個品種各生長階段(未現蕾、現蕾、開花、結角、成熟)的植株數,現蕾及現蕾后開花、結角、成熟的植株即為通過春化植株數,計算春化率,春化率=通過春化植株數/總植株數×100%。
1.2.2 冬前植物學性狀測定 秋播試驗于2019年8月22日在永登縣上川鎮進行播種,10月21日進行冬前苗期植物學性狀的記載統計。小區面積為1 m2,3次重復。依據《北方旱寒區白菜型冬油菜品種試驗記載規范》(NY/T2912-2016)進行[12],主要觀測記載油菜生長習性、心葉色澤、刺毛、葉色、葉脈色澤和葉緣共6個非數值型性狀(表2),以及葉柄長度、葉片寬度、葉片長度和裂葉對數共4個數值型性狀。

表2 白菜型冬油菜非數值型性狀及其賦值Table 2 Code designed for qualitative traits in winter rapeseed (Brassica rapa L.)
1.2.3 半致死溫度測定與計算 盆栽試驗于2020年5月28日開始,播種后將盆栽置于培養箱中培養,取五葉期的盆栽苗葉片作為試驗材料,使用打孔器取各材料相同大小的小圓片3片,置于10 mL離心管中并加入200 μL去離子水,以空白去離子水為對照。于低溫循環儀中設置6個溫度梯度(0、-4、-8、-12、-16、-20℃),每個溫度處理設置4次重復,溫度降至所設溫度處理保持30 min后取出,用去離子水定容至10 mL,搖床上振蕩2 h后,使用電導率儀測定其初電導率(S1)及對照電導率(Lck),測定完成后煮沸30 min,搖床上振蕩冷卻后,測定其最終電導率值(S2),求得相對電導率;利用相對電導率和溫度得到Logistic線性方程[13]并進行半致死溫度(LT50)的計算。
相對電導率(REC)=(S1-Lck)/(S2-Lck)×100%
1.2.4 油菜基因組DNA提取及檢測 取盆栽苗五葉期的冬油菜葉片,采用CTAB法進行基因組DNA的提取,利用1%瓊脂糖凝膠進行電泳檢測,并用核酸測定儀測定濃度,將提取得到的DNA母液稀釋成50 ng·μL-1于-20℃環境下保存備用。
1.2.5 PCR擴增及引物初篩 參照臧珊等[14]結果及蕓薹屬數據庫(http://brassicadb.org/brad/),選用均勻分布在10條染色體上的30對InDel引物、20對SSR引物和48對SRAP引物進行初篩,每對引物3次生物學重復,依據引物多態性和可重復性,總共篩選到27對核心引物。SSR和InDel引物名稱及序列見表3,SRAP引物及序列見表4,上游引物和下游引物隨機組合,其中篩選到4對SRAP引物組合分別為Em4/Me8、Em5/Me8、Em12/Me8和EM12/Me18。SSR和InDel標記總反應體系為10 μL,其中2×Taq PCR Master Mix為5 μL、10 μmol·L-1的引物各0.5 μL、50 ng·μL-1的DNA模板2 μL、滅菌雙蒸水2 μL。SRAP-PCR反應體系為10 μL,2×Taq PCR Master Mix為5 μL、上下游引物各0.6 μL,50 ng·μL-1的DNA模板2 μL,滅菌雙蒸水1.8 μL。擴增產物通過8%非變性聚丙烯酰胺凝膠電泳進行分離,電泳結束后采用硝酸銀進行銀染。
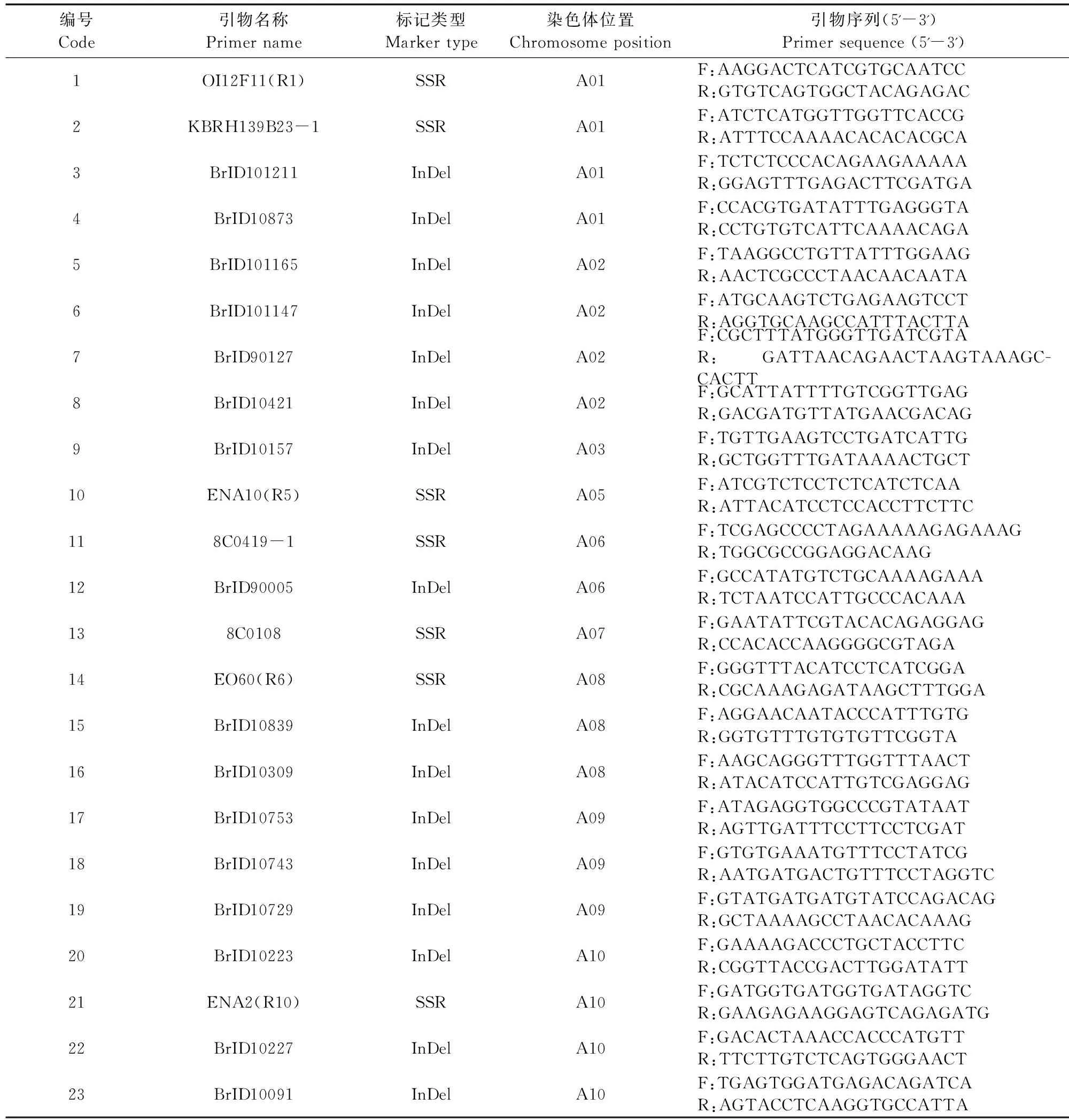
表3 標記名稱與序列Table 3 Marker names and sequences
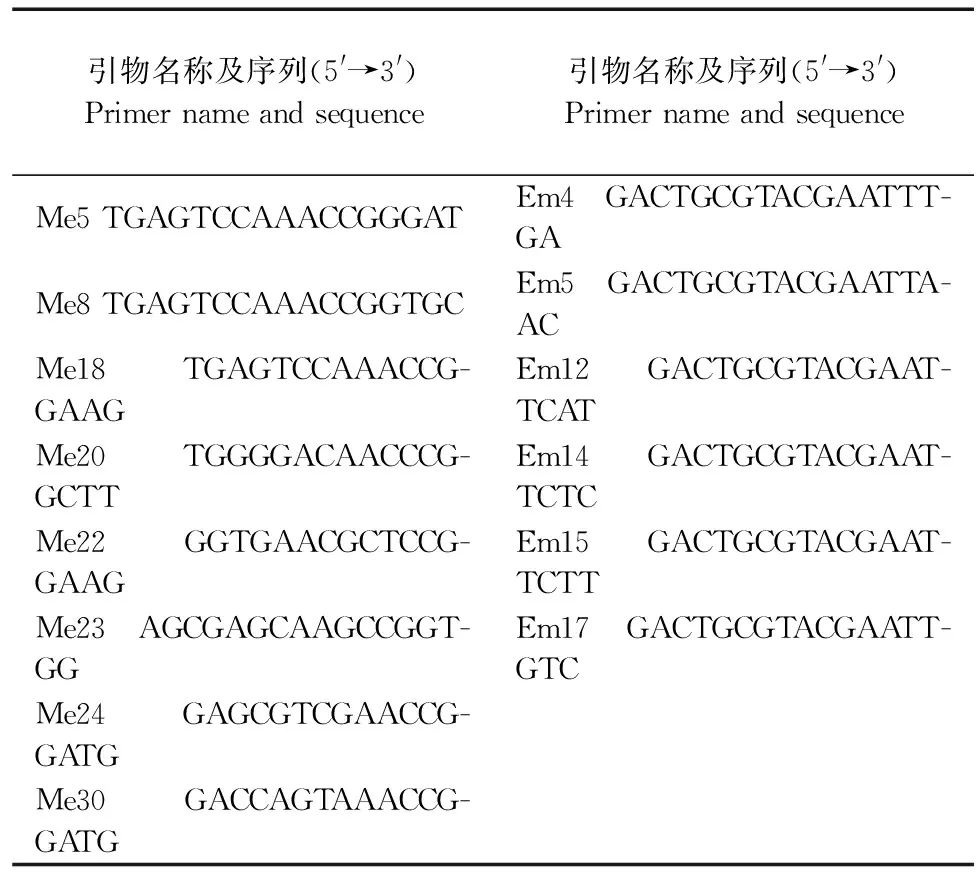
表4 SRAP引物序列Table 4 SRAP primer sequences
1.3 數據統計與分析
對26份北方白菜型冬油菜的6個非數值型性狀進行數值轉化處理(表2),4個數值型性狀進行分級處理[15],計算各植物學性狀的變異范圍、極差、平均數、變異系數、Simpson指數和Shannon-weaver指數,具體參考Pielou[16]的計算方法。
Simpson指數:
式中,Pi為某性狀第i級別內材料份數占總份數的百分比,ln為自然對數。
基于Excel和SPSS 22.0軟件對春化率、半致死溫度和植物學性狀進行統計及系統聚類。通過人工計數的方式識別26份白菜型冬油菜品種的凝膠電泳條帶,形成0/1矩陣,通過Dataformer軟件轉化成各軟件所需格式[17]。使用Popgen 32軟件計算各遺傳參數,PowerMarker V3.25軟件計算PIC和品種間的遺傳距離,得到非加權組平均法(UPGMA,unweighted pair-group method with arithmetic means)聚類結果。用iTOL對聚類結果進行修飾得到系統發生樹[18]。NTSYS-pc2.10e軟件基于遺傳距離矩陣,進行主坐標分析[19]。
2 結果與分析
2.1 不同北方白菜型冬油菜春播后的春化率差異
北方冬油菜感溫性強,但品種間有較大差異,不同品種感溫性差異可表現為其對春播條件下氣溫的適應性不同,主要表現為不同感溫性材料的田間春化率的不同,因此根據田間春化率的高低可以鑒別冬油菜抗寒性的強弱[7]。本研究的參試材料春播后,群體中各單株表現出不同的生長發育狀態,主要有未現蕾、現蕾、開花、結角、全結角和成熟共6種發育狀態(圖1,見9頁;表5)。根據田間生長發育狀態表型的統計結果,各材料春化率為6.59%~69.88%,臨洮WYW-1-4和16QD-15的春化率最低,分別為6.59%和6.94%,處于營養生長階段植株占群體總株數的比例分別為93.41%和93.06%;天油4號的春化率最高,為69.88%,其中現蕾期植株占群體總株數 26.51%。此外,天油258選1、2016PI-13 PZ24、天油142、2010元山區試天2、天油178、冬油8號、2010元山區試天5Ⅲ、臨洮延油2號-7共8份材料均表現出較高的春化率。

圖1 參試材料春播后的田間表現Fig.1 Field performance of materials after spring sowing
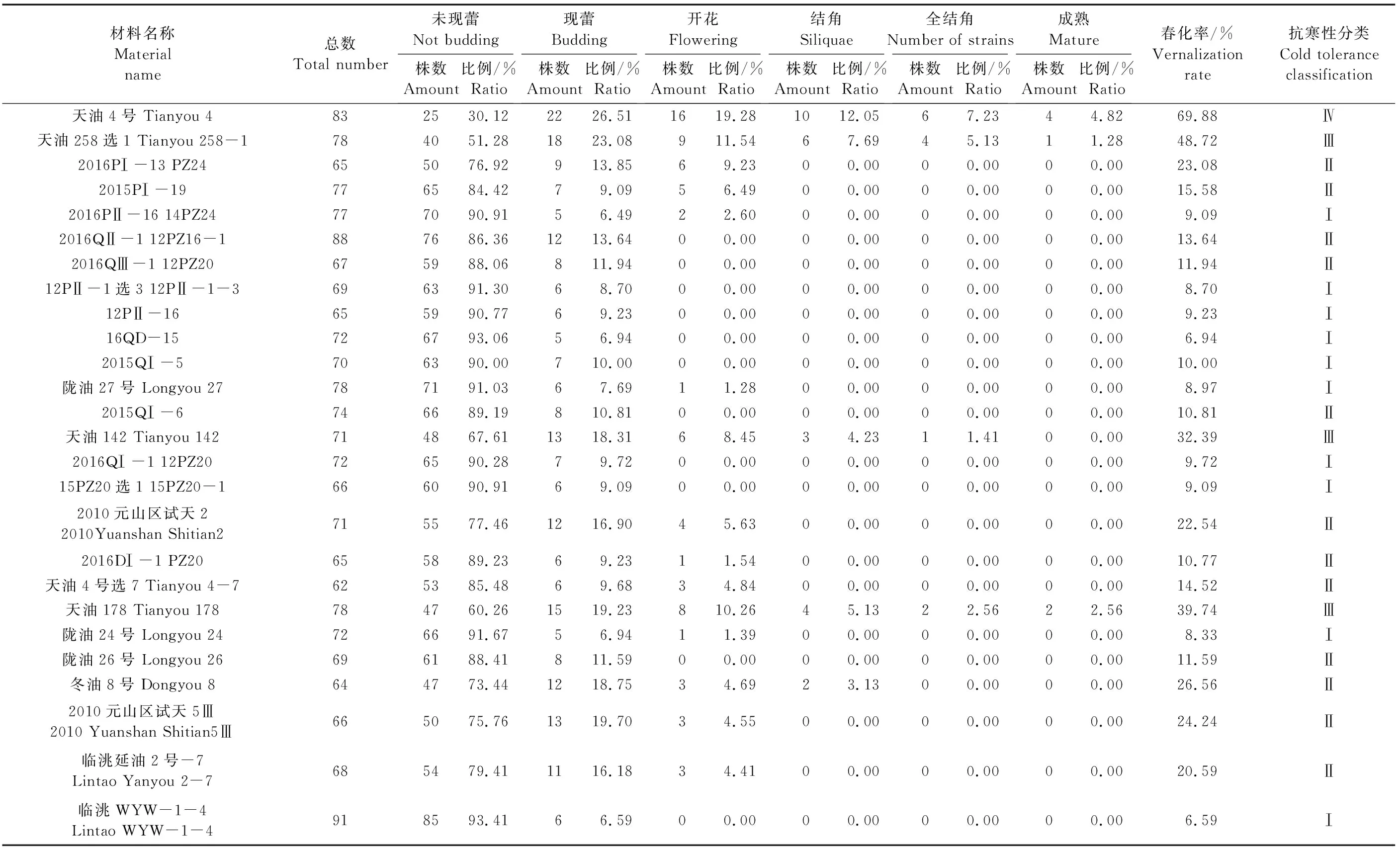
表5 供試材料春播后不同生長發育階段的植株數及所占比例Table 5 The number and proportion of plants at different growth and development stages after spring sowing
2.2 不同北方白菜型冬油菜抗寒性差異
低溫半致死溫度是評價植物抗寒性的常用指標,一般運用相對電導率結合Logistic方程計算,其高低反映抗寒性強弱。本研究根據葉片相對電導率擬合的Logistic方程的擬合度(R2)介于0.8344~0.9971,說明Logistic方程擬合度較好。26份參試材料的低溫半致死溫度變化范圍為-3.52~-9.93℃(表6),說明北方白菜型冬油菜各參試材料之間抗寒性差異較大,其中臨洮WYW-1-4的低溫半致死溫度最低,為-9.93℃;天油258選1的低溫半致死溫度最高,為-3.52℃。天水市農業科學研究所選育的材料包括天油4號、天油258、天油142等品種(系)抗寒性弱于隴油27號、臨洮WYW-1-4、隴油26號等白菜型冬油菜品種(系)。
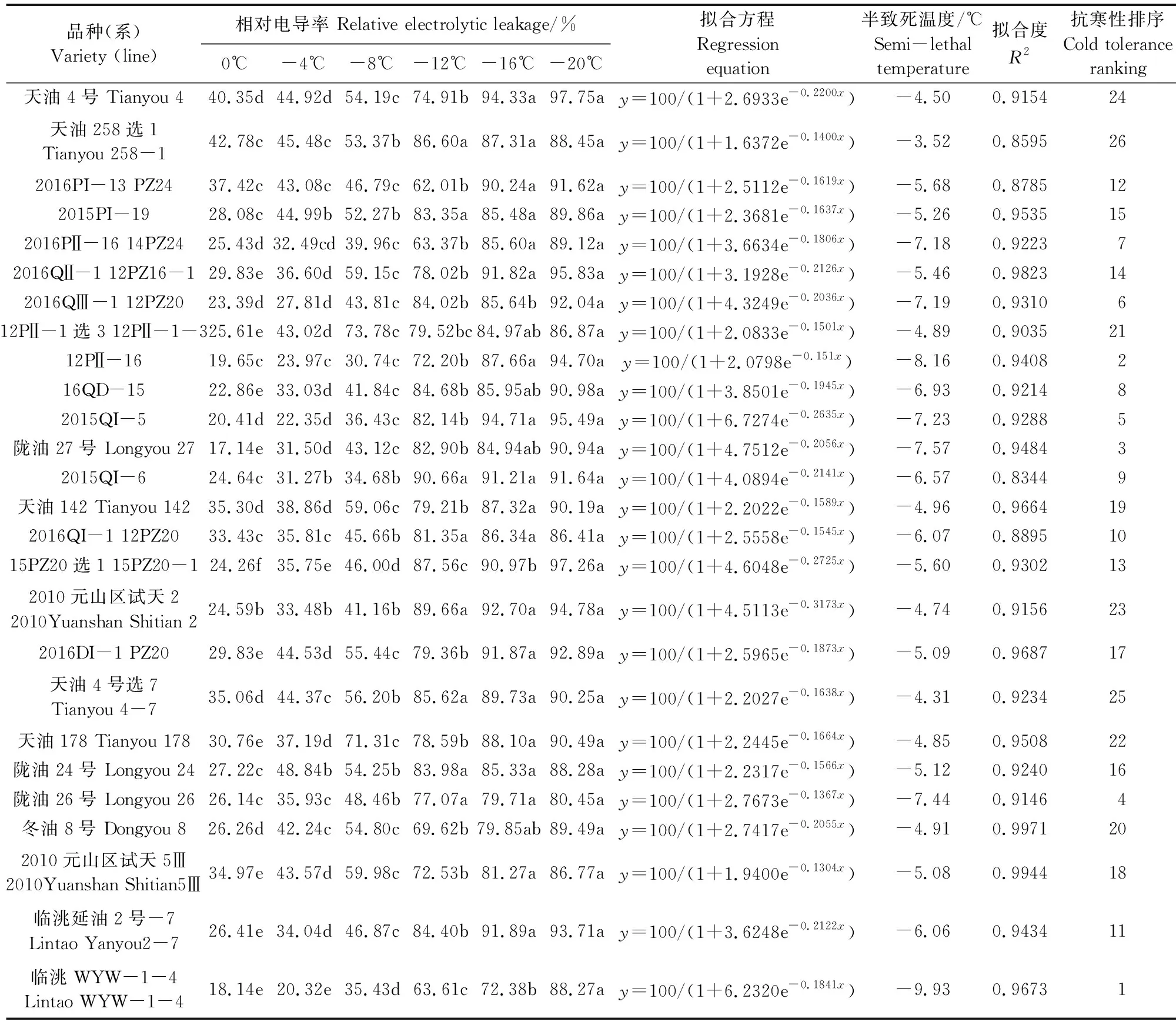
表6 26份冬油菜品種(系)的低溫半致死溫度Table 6 Semi-lethal low temperature of 26 winter rapeseed varieties (lines)
2.3 植物學性狀變化及其與抗寒性的相關性分析
通過對26份參試材料的植物學性狀觀測發現,各參試材料的生長習性有匍匐與半直立生長兩種類型,心葉色為淡綠或紫色,真葉顏色深綠居多,葉脈色多為白色,葉緣形狀多為鋸齒狀,真葉刺毛數量存在差異。對參試材料苗期4個數值型性狀(表7)及6個非數值型性狀(表8)分析結果表明,2016PⅡ-16 14PZ24葉柄長度和裂葉對數值最大,分別是12.30 cm和5.25對。葉片長度值最大的是2010元山區試天2(10.61 cm),最小的是臨洮延油2號-7(6.88 cm);葉片寬度值最大的是2010元山區試天5Ⅲ(9.04 cm),最小的是15PZ20選1(6.05 cm)。數值型性狀的變異系數平均值為25.39%,葉片寬度變異系數最小(21.59%),裂葉對數變異系數最大(32.00%);非數值型性狀真葉刺毛數量的變異系數最大(55.42%),生長習性的變異系數最小(7.54%),平均值為37.76%。數值型性狀的平均變異小于非數值型,說明其遺傳相對穩定。10個植物學性狀遺傳多樣性指數Simpson(D)的變化范圍為0.011~0.847,Shannon-weaver(H)的變化范圍為0.035~2.046。數值型性狀的遺傳多樣性指數D與H的平均值分別為0.826、1.917,說明參試材料的遺傳變異豐富。葉柄長度的遺傳多樣性指數最大,D、H值分別為0.847、2.046;裂葉對數的遺傳多樣性指數最小。非數值型性狀的遺傳多樣性指數D、H的平均值分別為0.395、0.627,說明數值型性狀的遺傳多樣性高于非數值型性狀,其中生長習性的遺傳多樣性指數最小。
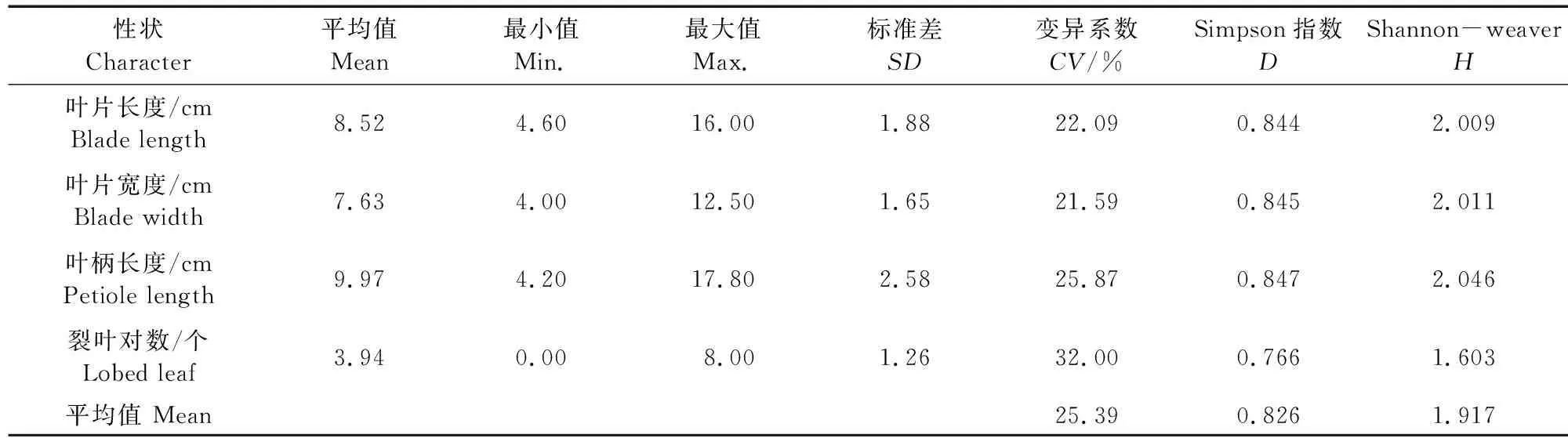
表7 26份參試材料數值型性狀的平均值、標準差、極差、變異系數及遺傳多樣性分析Table 7 Mean, standard deviation, extreme value, coefficient of variation and genetic diversity for numeric traits on 26 tested materials

表8 26份參試材料非數值型性狀的變異系數及遺傳多樣性分析Table 8 The variation parameter and genetic diversity of qualitative traits on 26 tested materials
基于植物學性狀對26份參試材料進行聚類結果表明(圖2),參試材料以歐氏距離5為界限,被分為4大類,第Ⅰ類包括2016QⅡ-1 12PZ16-1、隴油27號、2015PI-19、2016DI-1 PZ20、隴油24號、12PⅡ-16和2015QI-6共7份材料,占所有材料的26.92%;該類群的植株生長勢中等,匍匐生長,葉色為深綠,葉脈色以白色居多,真葉刺毛較多,葉柄長度、葉片長度和寬度及裂葉對數居中。第Ⅱ類包括12PⅡ-1 選3、隴油26號、2016QI-1 12PZ20、2015QI-5、16QD-15、2016PI-13 PZ24、15PZ20 選1、天油178、2016QⅢ-1 12PZ20、臨洮WYW-1-4和臨洮延油2號-7共11份材料,占所有材料的42.31%;該類群的特點是葉柄長度相對較短,心葉色以紫色居多。第Ⅲ類包括2010元山區試天5Ⅲ、2010元山區試天2、冬油8號、天油4號、天油258選1、天油142和2016PⅡ-16 14PZ24共7份材料,占所有材料的26.92%;該類群的特點是生長旺盛,葉片長度、寬度高于另外3個組群,生長勢相對較強,是甘肅天水的主要栽培品種。第Ⅳ類包括1份參試材料天油4號選7,僅占參試材料的3.85%,該組葉柄長度最長,葉片長度、寬度、裂葉對數明顯小于其他3個類群。

圖2 基于各參試材料植物學性狀的系統聚類圖Fig.2 Systematic cluster diagram of the tested materials based on botanical characters
春化率與其余的11個植物學性狀的相關性分析表明(圖3,見9頁),葉色與半致死溫度呈極顯著負相關(P≤0.01),葉柄長度與半致死溫度呈極顯著正相關(P≤0.01),說明白菜型冬油菜葉色越深、葉柄長度相對較短,其半致死溫度就越低。春化率與生長習性呈顯著正相關(P≤0.05),與半致死溫度呈極顯著正相關(P≤0.01),與葉色呈極顯著負相關(P≤0.001);半直立生長,其生長勢越強,地上部分生物量越大,不利于其越冬,該品種的抗寒性就越弱,春播時,其春化率就越高。葉色與低溫半致死溫度、春化率呈極顯著負相關,即葉色越深,多為深綠,其抗寒性越強,通過春化的植株所占比例越低。
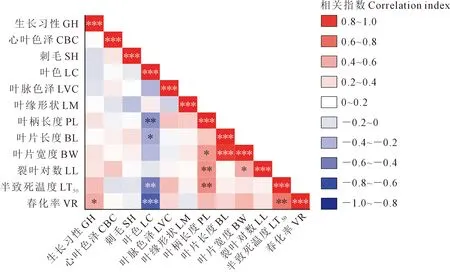
注:* P≤0.05;** P≤0.01;*** P≤0.001;Note: * P≤0.05; ** P≤0.01; *** P≤0.001; GH: Growth habit; CBC: Central blade color; SH: Spiny hair; LC: Leaf color; LVC: Leaf vein color; LM: Leaf margin; PL: Petiole length; BL: Blade length; BW: Blade width; LL: Lobed leaf; LT50: Semi-lethal temperature; VR: Vernalization rate.圖3 植物學性狀與抗寒性的相關性分析Fig.3 Correlation analysis between botanical characters and cold resistance
2.4 基于SRAP、SSR和InDel標記的遺傳多樣性及聚類分析
2.4.1 參試材料遺傳多樣性 利用27對引物對26份白菜型冬油菜種質的遺傳多樣性參數進行分析(表9),26份白菜型冬油菜的27對引物數據共檢測總等位基因數130個,其中引物基因座主要分布在A01、A02、A10這3條染色體。每條染色體的觀測等位基因數變化范圍為2~17,平均值為4.81,A01和A10染色體的觀測等位基因數最多。有效等位基因數(Ne)的變化范圍為1.26~6.81,平均值為3.14;觀測雜合度(Ho)的變化范圍為0.1238~0.7919;期望雜合度(He)的最小值是0.2081,最大值是0.8762;香農指數(I)和Nei’s基因多樣性指數(H)的平均值分別為1.2073和0.6168;多態性信息含量(PIC)的變化范圍是0.1833~0.8378,平均值為0.5681。其中SRAP標記的PIC值最高,以及PIC>0.5的位點有19個,占全部標記的70.37%,說明本研究所選的標記具有較高的多態性,可以有效地反映參試材料的遺傳多樣性信息。
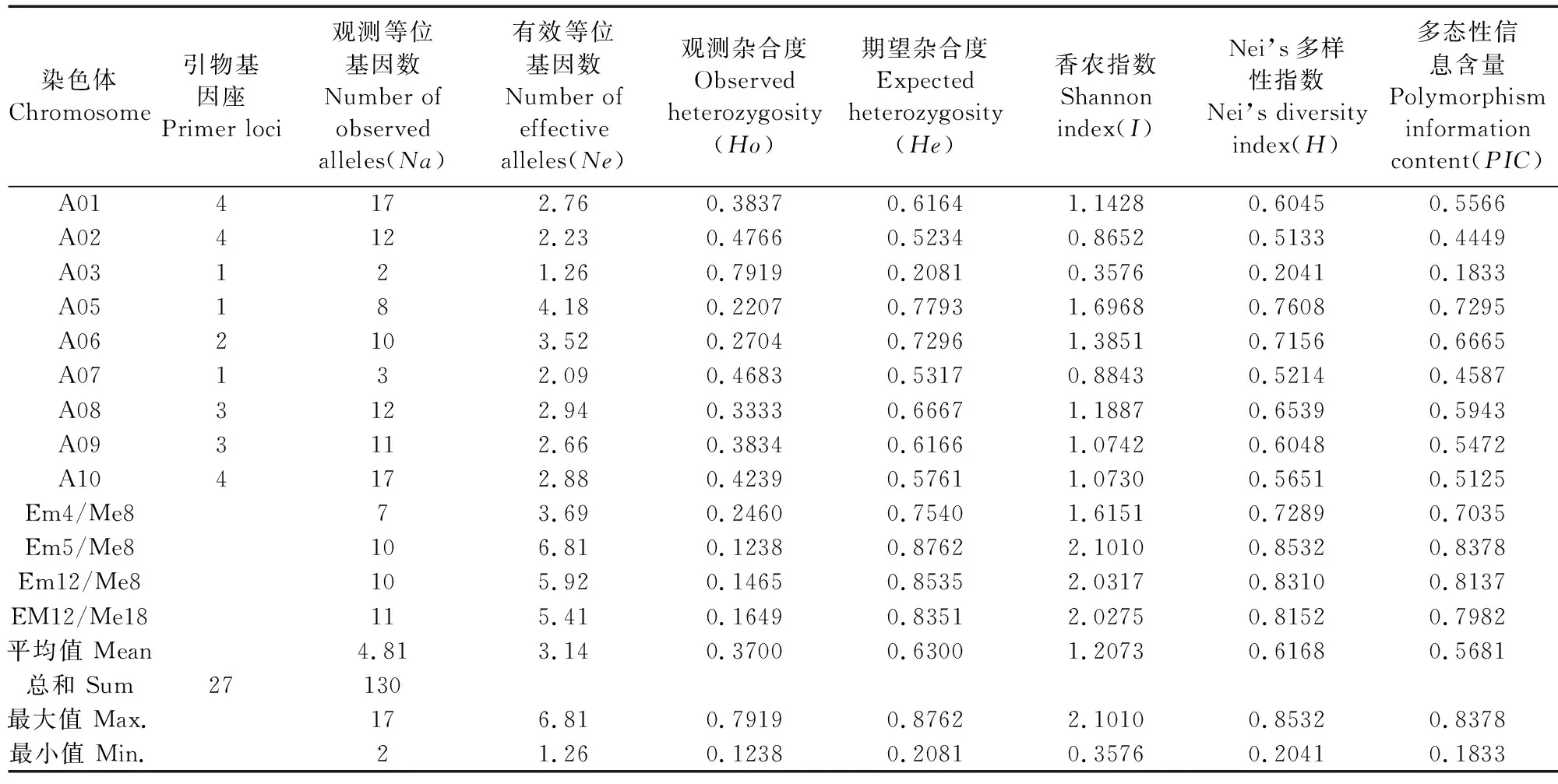
表9 26份冬油菜種質遺傳多樣性參數Table 9 Genetic diversity parameters of 26 winter rapeseed germplasms
2.4.2 基于遺傳相似系數的聚類分析 如圖4所示,26份參試材料分為6大類,第I類主要包括2010元山區試天5Ⅲ、2010元山區試天2、冬油8號、臨洮延油2號-7、2016PI-13 PZ24、2016QI-1 12PZ20和天油4號選7共7份材料;第Ⅱ類包括天油258選1、天油178、天油4號、天油142、2016DI-1 PZ20和隴油26號共6份材料;類群I和類群Ⅱ主要包括的是甘肅天水育成的材料。第Ⅲ類包括1份材料2015QI-6;第Ⅳ類包括2份材料15PZ20選1和12PⅡ-1選3;第Ⅴ類包括2015QI-5、16QD-15、隴油24號、2016QⅢ-1 12PZ20、2015PI-19和12PⅡ-16共6份材料;第Ⅵ類包括2016QⅡ-1 12PZ16-1、2016PⅡ-16 14PZ24、臨洮WYW-1-4和隴油27號共4份材料;類群Ⅲ、Ⅳ、Ⅴ和Ⅵ主要包括的是甘肅蘭州育成的材料。

圖4 基于SSR、SRAP和InDel標記聚類26份冬油菜種質Fig.4 Clustering of 26 winter rapeseed germplasm based on SSR, SRAP and InDel markers
2.4.3 依據SRAP、SSR和InDel標記的26份參試材料的主坐標分析 依據遺傳相似系數,對26份白菜型冬油菜品種進行主坐標分析,前3個主坐標的方差貢獻率分別為24.4%、13.7%和9.5%。對26個參試材料分別做第1、2主坐標二維圖的排序(圖5)。第1、2主坐標總的貢獻率是38.2%,參試材料被分為6類,其中第I類包括2016PI-13 PZ24、2010元山區試天2、2010元山區試天5Ⅲ、臨洮延油2號-7和冬油8號共5份材料。第Ⅱ類包括隴油26號、天油4號、天油258選1和天油178共4份材料。第Ⅲ類包括天油142、天油4號選7和2016DI-1 PZ20共3份材料。第Ⅳ類包括2015QI-6、15PZ20選1、12PⅡ-1選3、12PⅡ-16共4份材料。第Ⅴ類包括2015PI-19、2016QⅢ-1 12PZ20、隴油27號、2016PⅡ-16 14PZ24、臨洮WYW-1-4、2016QⅡ-1 12PZ16-1、2015QI-5、隴油24號共8份材料。第Ⅵ類包括2016QI-1 12PZ20和16QD-15共2份材料。

注:1~26代表品種(系)名稱,見表1。Note: 1~26 represent the names of the varieties (lines), as shown in Table 1.圖5 依據SRAP、SSR和InDel標記進行主坐標分析的第1、2主坐標散點圖Fig.5 Scatter diagram of the first and second principal coordinates of the principal coordinates analysis based on SRAP, SSR and InDel markers
3 討論與結論
感溫性是指作物由營養生長轉向生殖生長過程中對溫度條件的反應特性[7]。植物感溫性的田間直觀表現是春播條件下的春化率[20]。楊寧寧[7]和徐春梅等[20]研究都表明,感溫性不同的白菜型冬油菜完成春化階段所需的時間不同,隨著冬性由強到弱,春化所需時間由長到短,春化率由低到高。感溫性與抗寒性也存在內在關系。
本研究依據表型植物學性狀分析,將參試材料劃分為4個類群,類群I和類群Ⅱ主要是在蘭州育成的材料,其在生長習性、真葉刺毛、葉色等農藝性狀方面差異不大,表現出較低的春化率和半致死溫度,表明該類群可作為選育抗寒品種的優異種質資源。類群Ⅲ和類群Ⅳ表現出較好的生長勢,在葉柄長度、葉片長度、葉片寬度和裂葉對數等方面優于類群I和類群Ⅱ,表現出較高的春化率和半致死溫度,因此,該類群可用于冬季覆蓋作物材料的篩選。
通過對26份北方白菜型冬油菜品種(系)進行表型性狀多樣性分析,發現其數值型性狀平均多樣性指數高達1.917,高于非數值型性狀平均多樣性指數(0.627),說明北方白菜型冬油菜品種(系)表型性狀具有豐富變異,同李夢寒等[21]的研究結果比較相符。本研究發現,白菜型冬油菜非數值型性狀苗期生長習性的遺傳多樣性相對較低,真葉刺毛的遺傳多樣性指數相對較高,其主要原因是白菜型冬油菜苗期生長習性以匍匐生長為主,而真葉刺毛的多少受品種的影響,表現不同。
表型性狀容易受到環境因素的影響,因此,本研究在表型性狀的基礎上,采用分子標記技術從DNA水平上揭示品種之間的親緣關系。基于SRAP、SSR和InDel標記分析,將參試材料劃分為6個類群,類群I和類群Ⅱ主要是天水育成材料,其整體生長勢較強,春化率較高,主要是由于天水的地理位置、氣候等因素的影響,使該地選育成的品種(系)抗寒性弱于隴油27號、臨洮WYW-1-4、隴油26號等其他區域的白菜型冬油菜品種(系);類群Ⅲ、Ⅳ、Ⅴ和Ⅵ主要是在蘭州育成的材料,其表現出較低的春化率和較強的抗寒性;抗寒性劃分水平與植物學特征特性一致,強抗寒參試材料具有葉色深綠、葉柄相對較短的特征,而弱抗寒的類型具有葉色較淺、生長勢較強的特征。
