一種復合香型白酒糟醅微生物多樣性研究
2023-04-02 10:05:02薛江林楊貴張宿義范宏筠張煜亮梅海母娟楊明永杜鑫王程
釀酒科技 2023年3期
關鍵詞:優勢
薛江林,楊貴,張宿義,范宏筠,張煜亮,梅海,母娟,楊明永,杜鑫,王程
(1.瀘州國寶天釀集團股份有限公司,四川瀘州 646000;2.瀘州老窖股份有限公司,四川瀘州 646000;3.四川輕化工大學生物工程學院,四川宜賓 644000)
中國白酒由最初的濃香、醬香、清香3 種基本香型發展到如今的十三大香型[1],逐漸轉向多種香型組合發展的方向。20 世紀90 年代末,各大酒企就提倡“濃香為主、多香并舉”的生產發展戰略[2],以順應社會快節奏、多社交、多頻次等的消費需求[3]。目前針對白酒釀造工藝融合的研究,在國外研究較少,國內對于提高濃香型白酒出酒率及酒質的研究一直是行業內研究的重點,但在白酒釀造工藝融合的研究方面行業內一直在探索前進[4-6]。因此,本實驗在傳統瀘型酒釀酒工藝基礎上借鑒學習濃香型、醬香型、清香型白酒的釀造工藝,創新設計出一套特殊的白酒釀造工藝。通過分析不同工藝配料發酵過程的理化、風味、微生物以及白酒品質之間的關系不斷優化調味酒釀造工藝;由多輪次試驗得到三組較優的試驗方案,通過高通量測序技術對3 種試驗方案的堆積前、入窖、出窖的糟醅微生物多樣性、代謝途徑進行分析研究;結合糟醅理化性質,以及最終酒質的感官嘗評結果,確定出最優的工藝條件參數。本實驗為中國白酒調味酒種類的豐富提供了一定參考,也對不同香型白酒的生產技術做出了一定的歸納總結。
1 材料與方法
1.1 材料、試劑及儀器
材料:糯紅高粱,四川聯眾供應鏈服務有限公司;谷殼,瀘州市龍馬潭區永福米廠;高溫大曲,瀘州市醇香生物制造有限公司;河內白曲,山東梁山徐坊大曲有限公司;糖化曲,安琪酵母股份有限公司。試驗地點選在瀘州某酒廠生產車間。
試劑及耗材:氫氧化鈉、葡萄糖、鹽酸、氯化鈉、無水乙醇、五水硫酸銅、酒石酸鉀鈉、磷酸氫二鈉、磷酸二氫鈉,分析純,;2-辛醇,色譜純,Sigma-Aldrich;購自聚合化工。
儀器設備:TL2010S 中通量組織研磨儀,北京鼎昊源科技有限公司;SP-756P 紫外可見分光光度計,上海屹譜儀器有限公司;MLS-375L 全自動滅菌鍋,日本日立公司;DHG-9245A 烘箱,上海一恒科學儀器有限公司;DL-1 電爐,北京中興偉業儀器有限公司;5804R 高速冷凍離心機,德國艾本德公司。
1.2 試驗方法
1.2.1 工藝流程圖
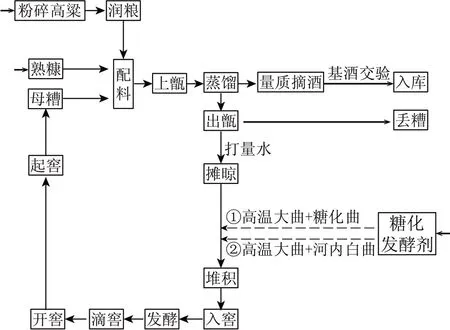
圖1 工藝流程圖
1.2.2 實驗設計
1.2.3 取樣
每個方案同時做3 口一樣大小(3.2 m×2.4 m×1.8 m)的平行窖池,取堆積前、入窖、出窖三個時間點的樣品,將統一方案的3 口窖池所采樣品置于無菌取樣袋中立即密封混合均勻,再將其分為2份,1份保存在-20 ℃用于理化特性檢測,1 份保存在-80 ℃用于基因組DNA提取。
1.2.4 理化指標檢測
理化指標檢測參照書目《白酒分析與檢測技術》和《白酒生產技術全書》中的步驟[7-8]。
1.2.5 DNA提取方法
稱取10 g 糟樣,用SDS[9]提取糟醅樣品中微生物的基因組DNA。基因組DNA 送往上海美吉生物醫藥科技有限公司進行高通量測序,測序數據在美吉生物云平臺(https:l/lcloud.majorbio.com/)進行處理。
1.2.6 基酒感官嘗評方法
基酒感官品評方法參考國標GB/T 33404—2016[10]、GB/T 33405—2016[11]。
1.2.7 數據分析利用SPSS 進行數據差異性檢驗分析,根據高通量擴增子測序數據,利用PICRUSt2(v2.2.0-b)對標記基因(16S/ITS)序列進行功能豐度預測,獲得每個樣本中對應的功能信息和豐度信息,并采用origin2018以及R語言進行繪圖。
2 結果與分析
2.1 理化指標分析
不同方案堆積、入窖、出窖糟醅水分、酸度、還原糖、淀粉含量如圖2 所示。糟醅水分含量為52.6%~59.5%,發酵過程中每個方案的水分都呈現先降低后增加的趨勢,變化差異顯著,加麩皮在堆積過程中對水分消耗相對較大。3個方案的酸度變化差異顯著,加麩皮能夠增加溶氧,在堆積過程中酸能夠快速被微生物代謝分解,在發酵過程中又能顯著增加。從圖2 可以看出,方案1 和方案3 堆積和發酵前后酸度變化差異最大,方案1 堆積前酸度最大,堆積后降為最小,發酵結束后又升至最大,方案2 在堆積和發酵前后酸度變化差異最小。3 種方案的還原糖和淀粉含量變化趨勢基本一致,差異不明顯,在堆積前后,方案2 還原糖含量增加相對大于方案1 和方案2,發酵前后,方案3 中淀粉含量消耗最大,方案1和方案2次之。

圖2 3種方案理化指標
2.2 微生物群落結構分析
2.2.1 糟醅微生物多樣性分析
對3 個不同方案糟醅樣品的ASV(Amplicon Sequence Variant)集、Chao1 指數和Shannon 指數進行比較分析(表2),可以看出,Chao1 和Shannon 指數都呈現先減少后增加的趨勢,方案1 和方案3 中細菌堆積前后豐富度變化巨大,表明加入麩皮能夠極大地降低細菌的多樣性,對真菌卻影響不顯著;方案2 整體變化相對較小;真菌堆積前后豐富度變化并不明顯,在發酵過程中增加。
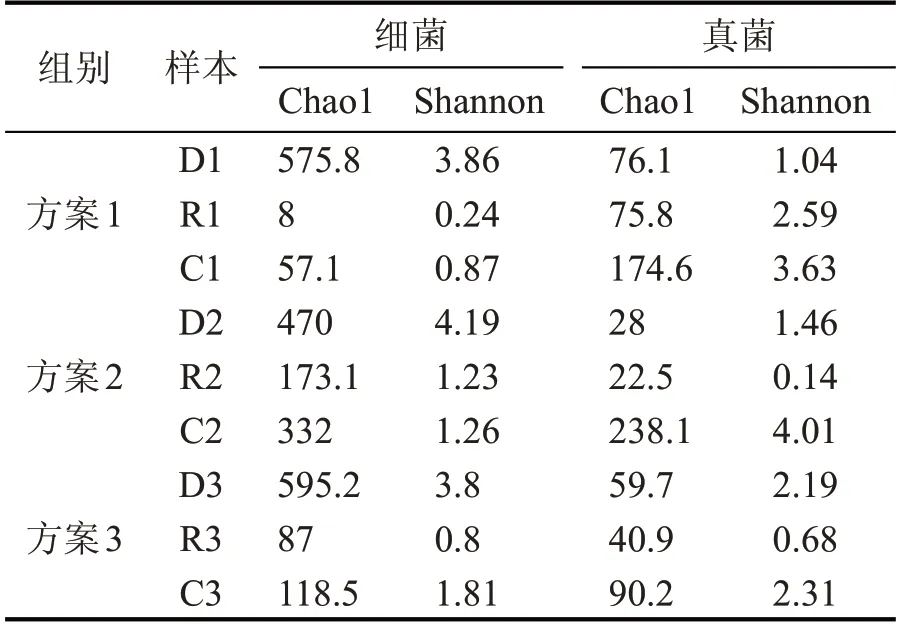
表2 不同方案的原核和真核微生物菌群多樣性指數
2.2.2 不同方案糟醅細菌群落結構對比分析
糟醅原核微生物群落組成(<1 %合并為Others)如圖3 所示,不同方案中共檢出18 種優勢細菌,細菌種類組成相似,豐度存在差異。從整體上來看,3 種方案在堆積和發酵結束時,入窖和出窖糟醅的細菌結構相似,優勢微生物分別為醋酸桿菌屬(Acetobacter)和乳酸桿菌屬(Lactobacillus),豐度超過90 %;不同點更多在于堆積過程,通過對比D1、D2 和D3,不難發現,D1 和D3 的細菌結構具有相似性,可能是由于都加有麩皮所導致,差別在于D3 中克羅彭斯特菌屬(Kroppenstedtia)豐度顯著高于D1;相較于D1 和D3,D2中Lactobacillus和芽孢桿菌屬(Bacillus)的豐度更小,糖多孢菌屬(Saccha-ropolyspora)和Solitalea的豐度更大。對比R1、R2和R3,在堆積過程中加麩皮能夠顯著增加疏松度,使得溶氧增加,好氧微生物的豐度顯著大于不加麩皮;在C3 中還存在一定量的Acetobacter,表明在方案3 的整個發酵過程中,Acetobacter因麩皮的增氧屬性,一直參與整個釀造過程。從配料差異分析,河內白曲和糖化曲在堆積前后對細菌群落結構的影響較小,麩皮影響較大。
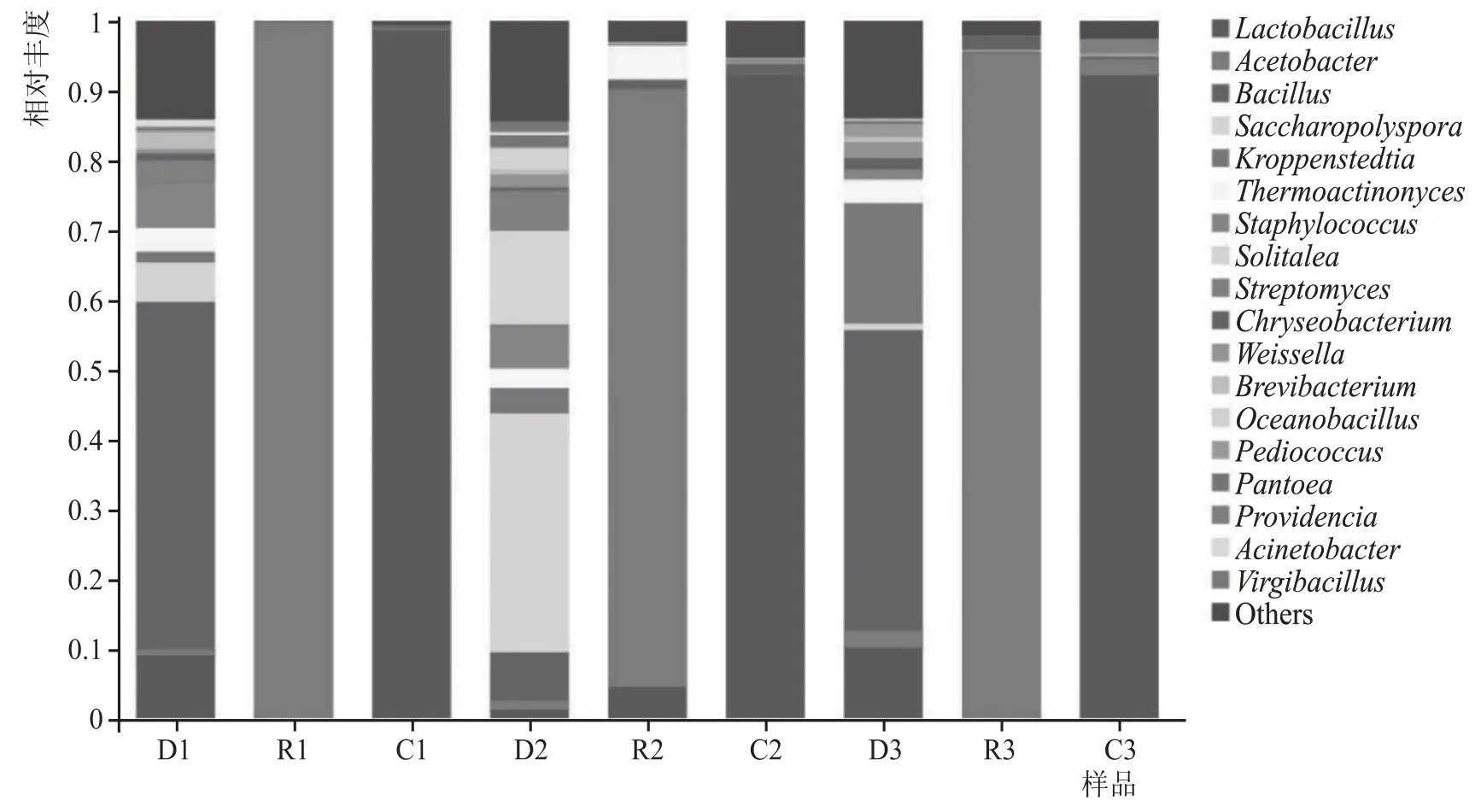
圖3 不同方案堆積、入窖和出窖糟醅細菌群落結構
2.2.3 不同方案糟醅真菌群落結構對比分析
糟醅真核微生物群落組成(<1 %合并為Others)如圖4 所示,不同方案中共檢出22 種優勢真菌,且真菌種類和豐度都存在顯著差異。對比發現,在堆積前,方案1 的真菌優勢微生物為嗜熱真菌屬(Thermomyces89 %),方案2為Thermomyces13 %和曲霉屬(Aspergillus76 %),方案3 為絲衣霉屬(Byssochlamys12 %)、Thermomyces31 %、Aspergil-lus16%、嗜熱子囊菌屬(Thermoascus30%)。堆積后,方案1 中優勢微生物以酵母屬為主,包括伊薩酵母屬(Issatchenkia29 %)、畢赤酵母屬(Pichia29 %)以及哈薩克斯坦酵母(Kazachstania25 %);方案2 優勢微生物以Byssochlamys98%為主;方案3 優勢真菌微生物為Byssochlamys87 %和Issatchenkia10 %。出窖后,方案1 優勢微生物群落結構復雜,包括Byssochlamys8.8 %、Thermomyces2.6 %、Aspergillus7.9 %、Issatchenkia27 %、Pichia 15%、青霉菌屬(Penicillium4.2%)、節擔菌屬(Wallemia7.4 %)、Thermoascus3.1 %;方案2 真菌微生物種類呈多元化,其中優勢微生物為Byssochlamys2.7 %、Thermomyces2.4 %、Aspergillus6.9 %、Issatchenkia9 %、Penicillium9.5 %、Pichia4.7 %、Wallemia23 %;方案3 優勢微生物包括Byssochlamys14%、Aspergillus6%和Penicillium52%。
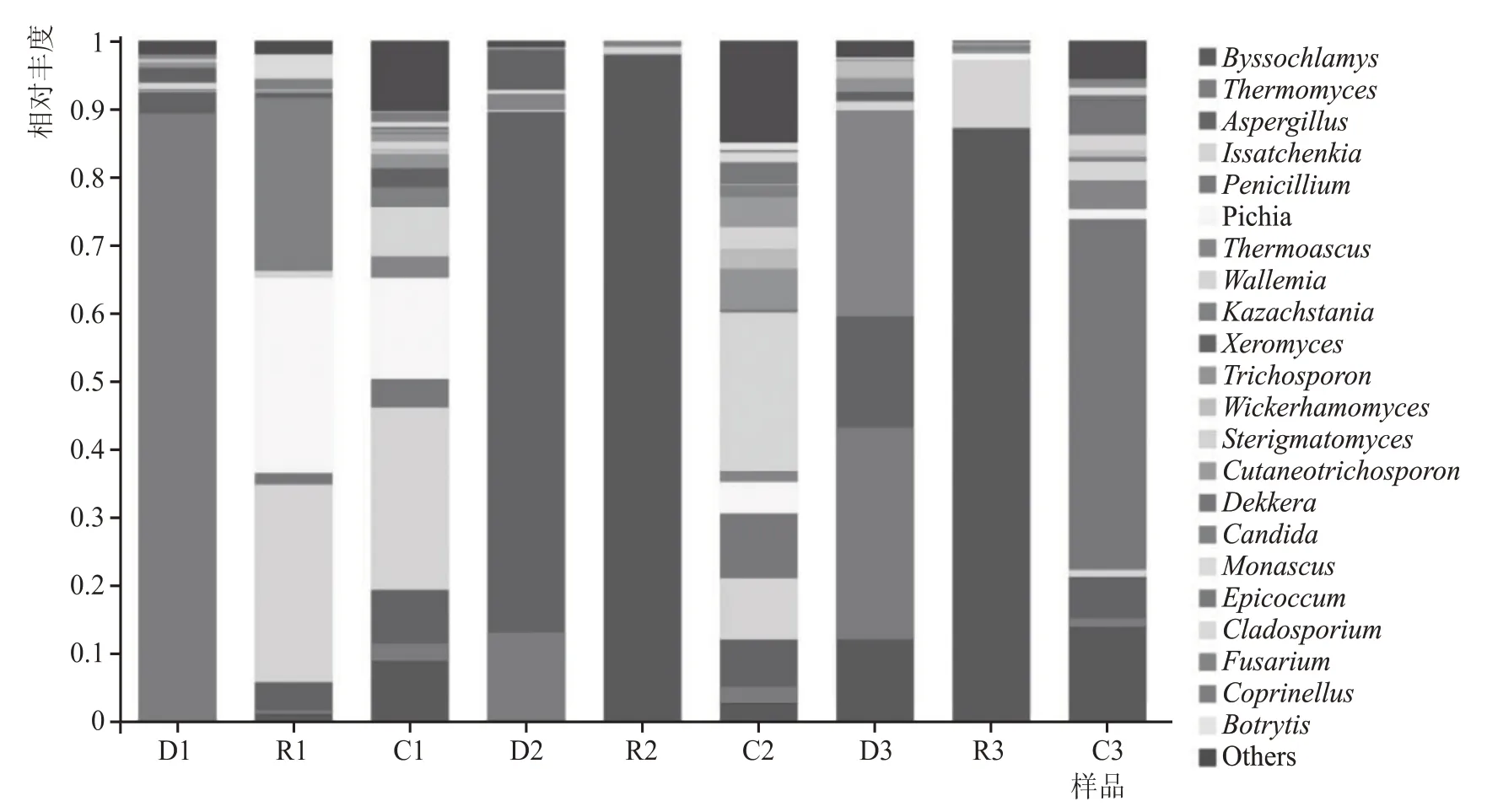
圖4 不同方案堆積、入窖和出窖糟醅真菌群落結構
對比3 種方案,從微生物的種類、多樣性和豐度進行討論,在堆積前期,不同種曲藥使優勢真菌微生物群落結構差異巨大;堆積過后,方案1 的優勢微生物主要集中在酵母屬,方案2 和方案3 的優勢微生物主要集中在霉菌屬;發酵結束后,各方案中真菌微生物種類基本一致,豐度各有不同。可見,不同種曲藥對整個發酵過程微生物群落結構的影響要遠大于輔料對白酒釀造的影響。值得注意的是,優勢微生物Thermomyces能夠產纖維素酶、淀粉酶、蛋白酶、β-葡聚糖酶和脂肪酶等酶類,為酵母的生長提供了重要動力,同時還能產生蛋白酶,為美拉德反應提供物質基礎,故該菌在白酒釀造過程中有著重要的作用[12-13];Aspergillus能夠合成纖維素酶、淀粉酶、蛋白酶、酯化酶、果膠酶,還能合成有機酸[14-15],這可能是方案2 在堆積過程中酸度降幅最小以及在堆積結束時Byssochlamys成為優勢微生物的原因之一。
2.3 微生物代謝通路及主要代謝功能分析
通過圖5 分析可見,代謝通路占比最大,3 個方案之間差異較大,方案1 和方案3 呈現先增加后減少的趨勢,方案2 呈現持續減少的趨勢。具體通過圖6 可以看出,在方案1、方案2 以及方案3 的代謝功能豐度對比中,可以明顯看出,方案2 要大于方案1 和方案3。在氨基酸、輔酶和無機離子代謝中,堆積后方案1 和方案3 豐度顯著增加,在發酵階段顯著降低,方案2 呈減小趨勢,表明加入麩皮確實能夠在堆積階段顯著增加氨基酸、輔酶以及無機離子的代謝。然而通過對比發現,方案1 和方案3 在堆積過程中氨基酸、輔酶以及無機離子的COG 代謝豐度相較于方案2 更小,表明加入麩皮,糟醅中空隙增加的同時也使得細菌微生物的平均豐度降低,從而在堆積前COG 代謝功能豐度降低,進一步表明方案2 對含氮類物質的代謝消耗更多。在碳水化合物的代謝中,方案1 和方案3 在堆積前后變化不大,在發酵過程中,逐漸增加;方案2 隨著釀造時間的增加,碳水化合物的代謝途徑呈現先減小后增加的趨勢,表明方案1 和方案3 中的細菌微生物對淀粉、糖等物質的消耗速度一直在增加,方案2在堆積過程中細菌微生物對淀粉、糖等物質的消耗速度逐漸降低,在發酵過程中逐漸增加,堆積前的代謝豐度卻大于方案1 和方案3。結合圖2 可以看出,在入窖和出窖糟醅中,3 種方案的優勢細菌微生物都是Acetobacter和Lactobacillus,且豐度相差比不顯著,說明3 種方案發酵過程中細菌微生物對淀粉、糖等的消耗速度基本一致,在堆積前后存在較大差異。
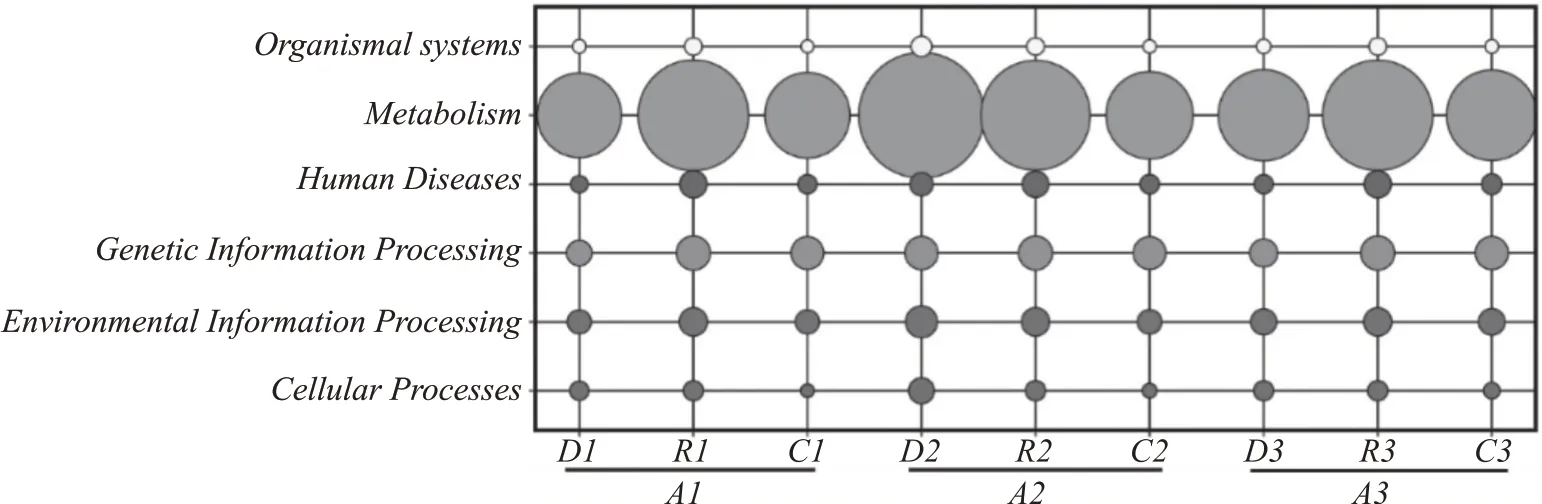
圖5 各方案堆積和發酵前后細菌微生物群落KEGG通路分布
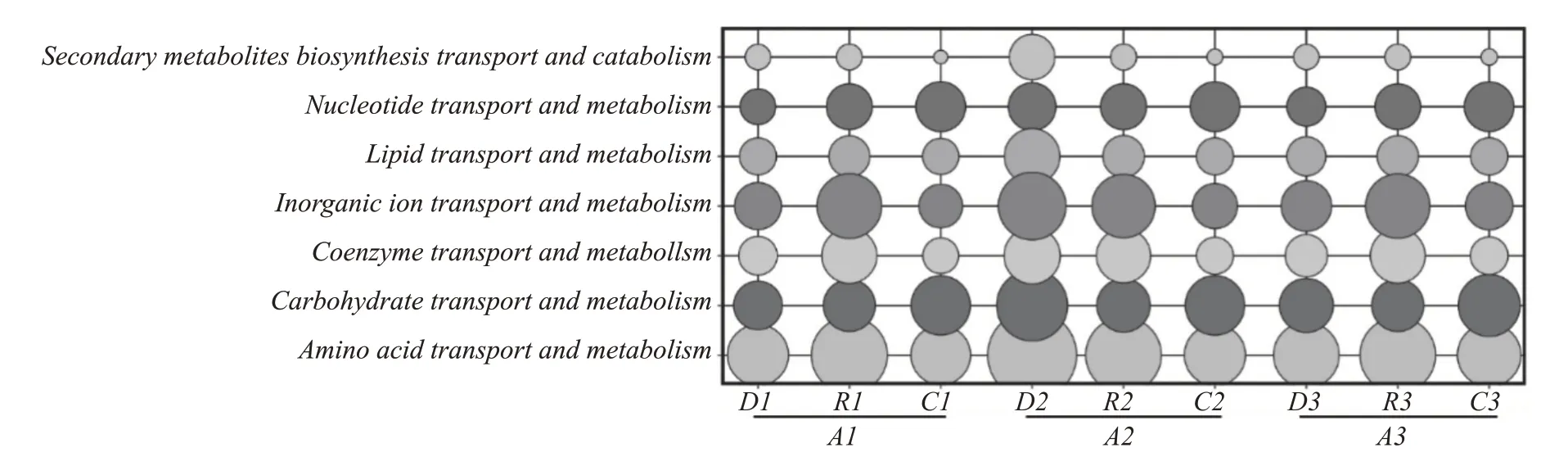
圖6 各方案堆積和發酵前后細菌微生物群落主要COG代謝功能分布
2.4 基礎酒感官嘗評結果分析
組織專家對三個試驗方案三個輪次共九個綜合酒樣進行感官品評,對每個試驗方案的三個酒樣分數進行綜合平均排序,專家組由4 名釀酒專家和6 名品酒專家組成,白酒感官品評嚴格按照國家標準GB/T 33404—2016 進行,評分細則主要包括:色澤(5分)、香氣(25分)、口感(60分)、風格(10分)四個方面。專家組給出的綜合平均結果如表3所示。

表3 基礎酒綜合樣對比品評記錄表
對評價結果繪制風味雷達圖如圖7所示。

圖7 基酒感官評價雷達圖
結合基礎酒的品評結果及圖7 可以分析總結出,經過高溫堆積、入窖發酵、量質摘酒、長期儲存等釀酒工藝后,所釀基礎酒在色澤、香氣、口感、風格等方面融合了濃香型、醬香型白酒風格之長,特點突出、復合優雅。三實驗方案三個輪次共9 個酒樣的酒體皆表現為無色或微黃透明、復合感強、風格獨特,特別是方案F1(高溫大曲+糖化曲)放置一年以后的基礎酒酒體顏色明顯微黃,醇厚綿甜、豐滿協調、具有非常舒適優雅的復合香氣。
3 結論
通過對釀造過程中微生物多樣性、代謝通路以及基酒的感官品評,發現不同方案之間差異巨大。方案2(高溫大曲+糖化曲)的評價結果優于方案1(高溫大曲+河內白曲+麩皮)和方案3(高溫大曲+糖化曲+麩皮)。通過多輪次實驗,最終確定方案2更適用于特殊香型白酒的釀造。通過研究不同香型的工藝融合,分析不同工藝配料發酵過程的理化、風味、微生物以及白酒品質之間的關系來不斷優化調味酒釀造工藝,為豐富中國白酒調味酒種類,創新傳統中國白酒的生產工藝以及滿足消費者多元化需求奠定了堅實基礎。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
